
Чем глубже изучаем мы социальность у животных, тем яснее необходимость провести грань между «социальным» и «несоциальным» в их поведении и действиях. Как отличить взаимодействие независимых индивидов (которые в рамках видовых форм социальности дифференцировали свои поведенческие роли, взаимодействуя в определённом сообществе, и дифференцировали настолько, что сообщество кажется «сверхорганизмом») от настоящих колоний с дифференцированными зооидами. Скажем тех, что встречаются у сифонофор?
Или поставим вопрос шире – как отличить социальное взаимодействие от «механического» управления в «настоящем» организме? Первое предполагает независимых индивидов, каждый из которых преследует свои «интересы» или действует на основе собственных побуждений, а если управляется социальной связью извне, так это управление устанавливается после взаимодействия и держится настолько, насколько устойчивы его результаты.
Во втором случае часть (орган, ткань, аппарат и т.п. единицы морфологической иерархии) не просто «подчиняется целому», реагируя нужным образом на нужный гормональный или нервный сигнал, её биологическая форма образуется целым и поддерживается отличной от форм других морфоструктур ровно настолько, насколько необходимо для функционирования организма.
Иными словами, в случае «несоциальных» (механических) взаимодействий речь не идёт ни о независимости единиц, образующих соответствующее системное целое и объединённых системными связями, ни о «конфликте интересов» между ними, ни о конкуренции за долговременный выигрыш, тогда как в социальных взаимодействиях всё перечисленное как раз и составляет суть дела [1].
Сообразно этим различиям Букварёва с Алещенко выделяют два типа противоположно организованных систем, иерархические и статистические. Идеальный пример первых — организм с внутриорганизменной регуляцией через «команды», распространяющиеся системами нервной и гуморальной регуляции. Примеры второй – общество человека, сообщества животных, связанные с их видоспецифической социальной организацией, и популяционные системы, от локальных популяций до биологического вида включительно.
Во втором случае регуляторный сигнал распространяется специализированными знакомыми системами, передающими «позиционную информацию» о существенных событиях и взаимодействиях за пределами непосредственного окружения независимых индивидов, за счёт чего происходит своего рода дальнодействие на таких масштабах времени и пространства, которые недоступным индивидуальным усилиям, с жёсткой кооперацией последних результатами коммуникации. Всё это предполагает свободу выбора, активное уклонение особей от ретрансляции системных сигналов или, наоборот, активное следование коммуникативным «правилам игры», с последующей сортировкой особей первой и второй категории между подразделениями системы по результатам выбора каждой из них (п.3).
Противоположные полюса в противопоставлении иерархических систем статистическим – организмы и популяции – прекрасно отличаются друг от друга, проблемы спутать их просто не возникает. А вот с социумом / колонией ситуация хуже. Социум с дифференцированными ролями индивидов кажется трудноотличимым от колонии с дифференцированными зооидами, особенно учитывая распространённость метафоры «сверхорганизма» в исследовании эусоциальных видов животных, в первую очередь беспозвоночных. И не просто «распространение», но успешное использование этой метафоры для понимания принципов социальной зависимости и социальной регуляции, её способность «наводить» на продуктивные обобщения и т.п.
Опять же специфическая морфология зооидов у колониальных беспозвоночных формируется той же децентрализованеной регуляцией, что и дифференциация поведенческих ролей особей разного статуса в сообществах животных.
И по мере того, как эусоциальность описывается у всё новых групп животных, в том числе у таких низкоорганизованных форм, как сосальщики Himasthla, и одновременно столь высокоорганизованных, как голые и дамарские землекопы, вопрос о том, как провести грань между «социальным» и «морфологическим» формообразованием, социумом и организмом (колонией) из философского делается естественнонаучным.
Применительно к социальности у животных он имеет две стороны.
- Действительно ли муравейник это сверхорганизм, или отдельные особи в муравейнике не только сохраняют собственную индивидуальность, но и коллективные взаимодействия вместе с внутрисистемными формами регуляции не только не нивелируют индивидуальность особей, но и формируют её?
- Действительно ли случаи, вроде описанного у сосальщиков, относятся к социальности, а не к колонии с дифференцированными зооидами, пусть даже непосредственно не соединёнными друг с другом, а взаимодействующими «через пустоту»?
Что касается муравейника, тут вроде бы ситуация проясняется. Это не «сверхорганизм», социальная среда муравейника не только не нивелирует индивидуальности рабочих особей, но, напротив, формирует её. Также как у людей свою неповторимую личность можно развить лишь в коллективе, выделившись в рамках общей работы на фоне остальных, развитие индивидуальности рабочих особей происходит через их совместную деятельность в группах фуражиров, охранников и т.п.
В работах Ивана Яковлева по рыжим лесным муравьям показано [2], что исходно рабочие муравьи различаются лишь уровнем агрессивности, а вот индивидуальные различия куда как более сложны и многосторонни. Они включают в себя поведенческую, социальную и когнитивную специализацию, которая развивается в групповом взаимодействии муравьёв за пределами муравейника с их пищевыми объектами, естественными врагами тлей – личинками сирфид, врагами типа жужелиц и хищниками вроде певчих птиц (три последних связаны с существенным риском, хотя и разной природы). И, соответственно, один и тот же опыт (скажем, отрицательный опыт взаимодействия с личинкой сирфиды, рис.2), у одной группы муравьёв увеличивает готовность идти на риск, у другой – стремление избегать опасности, так что по мере накопления опыта и совместной деятельности достигается всё более дробная дифференциация, «доходящая» до индивида (рис.4).

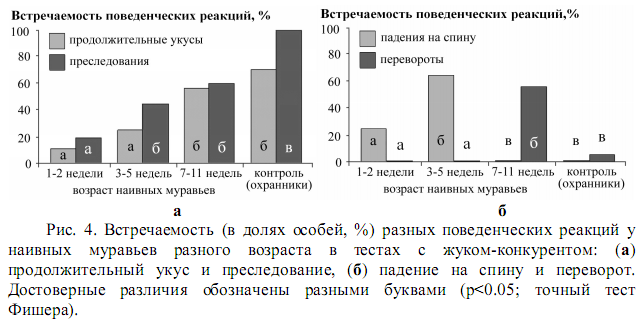
«На выходе» рабочие особи различаются ещё и по таким признакам, как уровень исследовательской активности, способность избегать опасности и накапливать опыт, причём межиндивидуально, а не только между функциональными классами охотников, охранников и сборщиков пади. Это хорошо видно при сравнении с искусственно сформированными семьями «наивных» муравьёв с точно датированным возрастом: индивиды «настоящего» муравейника того же возраста показывают много большую дифференциацию и более богатый синдром индивидуальных особенностей. Соответственно, по мере накопления опыта у всех индивидов по сравнению с «наивными муравьями» увеличивается адекватность реакций на «врагов».
Скажем, у наивных охранников она «на градус ниже» требуемого по агрессивности и готовности пойти на самопожертвование, вцепившись в жужелицу мёртвой хваткой, у охотников – «выше требуемого». По мере накопления опыта вторые учатся короткими ударами прогонять жука вместо реакций, к которым их подталкивает исходно повышенная агрессивность, охранники – наоборот, а соотношение тех и других действий (вкупе с исследовательской активностью) составляет индивидуальные различия. И, конечно, серьёзно уточняется «образ врага» у тех особей, кто по долгу службы должен взаимодействовать с ним, опять же, у разных в разной степени (рис.5). Наивным муравьям для адекватной реакции нужен целостный образ жужелицы, опытные могут по отдельным ключевым признакам «достроить» его и правильно среагировать.

Опять же у муравьёв вполне развито социальное обучение, и большинство видоспецифических стереотипов «спящие», т.е. нужен соответствующий индивидуальный опыт, чтобы их «разбудить» и «достроить» до эффективного и адекватного действия, а это предполагает развитую индивидуальность. По крайней мере, у позвоночных предполагает, начиная аж с рыб, и муравьи тут никак не должны отличаться.
Всё перечисленное позволяет использовать понятие «индивидуальность», ранее относимое лишь к «социальным» видам позвоночных [3](существенно, что к «несоциальным» видам оно неприменимо!), к отдельным особям в муравьином сообществе. Аналогичные проявления индивидуальности были зафиксированы у ос Polistes и, видимо, присущи всем перепончатокрылым. (А вот как с термитами, не знаю).
Но, так или иначе, грань между «социальным» и «несоциальным» взаимодействием, сообществом и колонией интересно определить в общем виде, теоретически, чем я и попробую заняться. Грань эта тонка, но отчётлива, как все социальные грани, никакого «континуума плавных переходов» между социумом и колонией (с дифференцированными зооидами), социальным взаимодействием и «механическим» управлением внутри организма нет и в помине. Все перечисленное – диалектические противоположности, которые, как им и положено, находятся в единстве и в состоянии борьбы, чем и осуществляется прогрессивное развитие системы.
Как эту грань провести? Лучший критерий я нашёл у американского философа и педагога Джона Дьюи:
«Даже работая вместе для достижения общей цели, люди вовсе не обязательно составляют социальную группу. Детали машины тоже работают согласованно, ради общего результата, они ведь не образуют общества. Вот если бы они осознавали общую цель и были настолько заинтересованы в ней, что согласовывали бы с этой целью свои конкретные действия, тогда – да, они образовали бы общество. Но для этого надо общаться [4]. Каждый должен знать, чем занимаются другие, и иметь возможность постоянно держать их в курсе своих намерений и достижений. Консенсус [5] невозможен без общения.
Таким образом, мы вынуждены признать, что даже внутри самых социализированных групп существует множество отношений, которые на деле не являются социальными. Большое число человеческих взаимоотношений в любой социальной группе всё ещё находится на машиноподобном уровне. Нередко одни люди используют других для достижения своих целей, не обращая никакого внимания на их эмоциональные или интеллектуальные установки и не спрашивая их согласия, а интересуясь только результатом. Так ведут себя люди, обладающие превосходством в физической силе, социальном положении, ловкости, технических или финансовых возможностях. И пока отношения между учителем и учеником, нанимателем и работником, начальником и подчинённым остаются на этом уровне, все они не образуют подлинно социальную группу, даже если работают вместе. Приказы и их исполнение влияют, конечно, на то, как протекает деятельность, и каковы её результаты, но сами по себе не обеспечивают совместности целей, не создают общности интересов»
Задача натуралистов – «перевести» эту мысль философского характера на корректный естественнонаучный язык. Если, конечно, факты естественной истории «социальных» и «несоциальных» видов животных подтверждают эту идею «в обе стороны»: социальное взаимодействие в противоположность «органическому» предполагает индивидуальность и связано с коммуникацией, несоциальное исключает индивидуальность и свободу выбора, предполагая «механическое» (или «автоматическое») подчинение стимулу.
Так вот оказывается, что определение Дьюи в полной мере применимо к социальности животных – даже больше чем к человеческому обществу, так как здесь много легче достичь полного «машиноподобия» взаимодействий, превращающего сообщество в колонию, поскольку коммуникация у животных в отличие от нас своего рода «предмет роскоши».
Действительно, наблюдая взаимодействия одного класса, например, агонистические или брачные, мы видим чёткое противопоставление «социальных контактов» «несоциальным», как только начинаем анализировать происходящее со структуралистской стороны (вместо функциональной).
Структура поведения в «социальных» и «механических» взаимодействиях оказывается прямо противоположной, без каких-либо переходов между этими классами, притом, что в функциональном отношении это одно и то же.
Скажем, с функциональной точки зрения ухаживания у моногамных видов птиц и млекопитающих вроде ничем не отличается от такового у промискуитетных и полигамных, собирающихся на токах. В обоих случаях самцы агонистически взаимодействуют друг с другом, распределяя территории по результатам побед и поражений в конфликтах. Затем на «охраняемое» и монополизируемое пространство они привлекают самок и дальше сближаются с привлечёнными самками, что завершается образованием пары (постоянной или временной, всё равно) и копуляцией – либо отвержением партнёра и привлечением новых, если «тестирование» оказывается неудачным.
Но стоит взглянуть на структуру поведения ухаживания самца за самкой, особенно на наличие-отсутствие специфических демонстраций-посредников, мотивированных именно сексуальными побуждениями, то противоположность организации процесса в первом и во втором случае становится очевидной (и эта противоположность именно того рода, что очерчена выше).
Дело в том, что у полигамных/промискуитетных видов и особенно у собирающихся на токах самцы взаимодействуют реально только друг с другом и лишь в контексте распределения территории, соответственно, мотивированном агонистически. Им определяется анизотропность пространства тока, с образованием градиента «центр-периферия» и с соответствующими различиями в поведении «центральных», «срединных» и периферийных самцов.
Самки же на ток привлекаются «автоматически», выбирая территории «лучших» (центральных самцов), тех, что демонстрируют наиболее интенсивно, развёрнуто и устойчиво. Сами самцы для достижения этого результата должны взаимодействовать лишь с другими самцами, а не с самками (и только в агрессивном, а не в сексуальном контексте). И в любом случае «привлекающие» демонстрации самцов мотивированы агрессивно; самки не отвечают на них своим демонстрированием, партнёры не вступают в сигнальный обмен, самка просто «подчиняется» релизерному эффекту, принимает позу подставления и с момента начала сближения не может «отказаться», отвергнуть самца, сменить его на соседа и т.п.
То есть собственно взаимодействие самца и самки завершающееся копуляцией, носит сугубо односторонний характер. В отличие от территориальных взаимодействий между самцами, где есть и коммуникация и опосредующие специфические демонстрации.
Напротив, у моногамных видов взаимодействие самца с привлечённой самкой – отдельный специфический контекст общения, предполагающий свои собственные брачные сигналы, и в любой момент сближения и «тестирования» потенциального партнёра самка может прервать процесс и отправиться на поиски другого. Также как и самец. Иными словами, коммуникация с наличием специализированных сигналов-посредников придаёт взаимодействию симметричность, в отличие от сугубой асимметрии на токах.
Так, у самца вОрона при образовании пары чётко виден ключевой момент «перелома взаимодействия», когда он переключается от демонстраций угрозы, адресованных самцам-соперникам [6], к демонстрациям ухаживания за самкой [7]. Первые демонстрации характеризуются выпрямленным положением корпуса, увеличенной, как бы «капюшонной» головой и шеей. Самец словно пытается оказаться выше потенциального адресата демонстраций. Во втором случае, напротив, он нагибается и кланяется, так что линия корпуса и движущегося клюва оказывается ниже корпуса самки (Lorenz, 1940; Gwinner, 1964).
Сказанное хорошо иллюстрируется данными по онтогенезу демонстраций у птенцов тетерева. Развивающееся брачное поведение молодых самцов тетерева, как и их территориальная агрессия, обычно направлено на других самцов, а не на самок. Птенцы тетерева, даже при сексуальных реакциях, гораздо чаще выбирали в партнёры самцов, нежели самок, особенно меньшинство самых агрессивных птенцов (Gwinner-Hanke, 1991). Это соответствует специфике взаимоотношений полов в токовой системе тетерева, где самка (если она уже привлечена на ток) – пассивный объект удовлетворения сексуальных побуждений самца, а не равноправный участник коммуникации, направленной на оптимизацию выбора партнёра у каждого пола. Самец просто «кроет» самку, привлечённую его территорией (качество которой символически выражено через «продукцию» демонстраций, песен и пр.), но отнюдь не вступает с ней в коммуникацию, сколько-нибудь аналогичную отношениям данного самца с соседними владельцами территорий на току.
Иными словами, «несвобода» взаимодействий выбора партнёра у полигамных и проимискуитетных видов (в особенности спаривающихся на токах) состоит в предопределённости этого выбора отношениями и взаимодействиями совсем иного контекста. Обычно это отношения территориальности или агрессивного агрессивного доминирования в группах. Собственно, именно поэтому их нельзя отнести к социальным взаимодействиям, они «машиноподобные» по Дьюи.
«Машиноподобие» достигает наибольшего выражения в явлении, которое у англоязычных этологов называется mate choice copying, что на русский осмысленно перевести как «копирование (или тиражирование) чужого выбора». Впервые оно было обнаружено у тетерева.
Обычно предполагается, что в силу конкуренции самцов на току и активного выбора их самками выбор и спаривание разных самок должны быть взаимно-независимыми событиями. Оказывается, это совершенно не так. Самки посещают ток и перемещаются между территориями самцов группами, так что каждая видит сближение с самцами и спаривание всех остальных. Как только первая самка спарится, прочие члены группы с очень высокой вероятностью слепо повторяют её выбор. Верно и обратное – как только самец спаривается хотя бы раз «на глазах» других членов группы самок-посетительниц, его привлекательность для остальных резко подскакивает – и тем сильней, чем крупнее ток, выше интенсивность демонстрирования собравшихся на нём самцов и т.п.
Иными словами, привлекательность самца для самок есть система с положительной обратной связью. Это что подтверждается, во-первых, прямыми наблюдениями на выборке в 19 токов. Анализируя временные ряды изменений в брачной активности самцов резидентов, мы видим, что как только какой-то один привлёк самку и спарился, это немедленно повышает вероятность его выбора другими самками из числа находящихся на току, обычно ближайшими к предыдущей, хотя бывают случаи и подлёта. Во-вторых, это подтверждается прямым опытом с подстановкой чучел самок на территории разных самцов. На пике токования возбуждённые самцы пытаются спариваться даже с чучелами и это делает их куда более привлекательными для живых самок (Höglund et al., 1990, 1995).
Это показано для самок и других видов, спаривающихся на току; у не-токовых видов отмечено тиражирование выбора партнёра и для самцов, не только для самок.
Иными словами, у самых разных видов животных выбор партнёра как самцами так и самками оказался не независимым, а подверженным социальным влиянием, обусловленным «копированием выбора» — в той мере, в какой соответствующие взаимодействия являются «механическими», не связаны с опосредованием сигналами и коммуникацией.
То есть во всех этих случаях субъектом выбора оказываются не отдельные индивиды, а их объединения, связанные специфическими социальными связями (по которым предсказывается образование именно таких объединений, а не иных), причём не имеющими отношения к собственно размножению. Проблема эта интенсивно обсуждается в современной этологии, ибо плохо укладывается в социобиологические максимы.
Аналогичный эффект «тиражирования выбора партнёра» найден у людей. Мужчины и женщины ранжировали по привлекательности фото представителей противоположного пола (из своей среды; о кросс-культурных исследованиях я не знаю). А затем показывали тех же самых лиц уже не одиночку, а в паре с более или менее привлекательным представителем пола собственного. Оказывается, это сильно изменяет оценку, соответственно увеличивая или уменьшая её, и «масштаб сдвига» можно прогнозировать по интенсивности перевода взглядов с одного члена парноё фотографии на другого. Понятно, что у людей данный эффект существует в рамках «социального бессознательного»; как только люди осознают, что действовали не самостоятельно, а уступили социальному влиянию, они могут освободиться от него – в отличие от тетеревов, перепелов, пецилий и гуппи [8].
Взаимодействие выбранного самца и самки после того, как она пришла на его территорию, тоже обходится без коммуникации. Он её просто «кроет» без каких-либо опосредующих демонстраций. Лишь единственный раз «честность коммуникации» самца и самки подвергается «испытанию» — но тоже со стороны соседних самцов, при попытке сближения образующих неспецифические «помехи» самцу, привлёкшему самку и также в в агрессивном контексте. Выбранный самкой самец должен мочь сохранить устойчивость демонстрирования несмотря на помехи, созданные вторжением соседей, тогда он сможет подойти и покрыть самку, а она не проявит агрессию, не убежит и т.п., так что коммуникация будет успешной. Но эти демонстрации тоже мотивированы территориальной агрессией и адресованы не самке, а конкурентам за территорию или за социальный статус – что у тетеревов/манакинов и т.п. на току, что у павлина с его гаремной полигинией в гареме.
Вот как это выглядит, скажем, у каменных петушков Rupicola rupicola — примитивных воробьиных птиц-котинг, самцы которых собираются на токах, как и родственные котингам манакины. Исследования тока данного вида в Суринаме показали важную роль социально обусловленных помех брачному поведению самцов как фактора отбора «лучших демонстраторов», которые в основном и спариваются с самками. «Помехи» состоят во вторжениях соседних взрослых самцов (30%), реже – годовалых самцов, которые регулярно посещают ток и демонстрируют в присутствии самок, хотя приобретают территории только к 3-4 годам (11%).
Ещё реже помехи создают сами самки, когда реагируют агрессией на попытки сближения самцов. Посторонние особи вторгаются обычно в момент сближения владельца территории с привлечённой им самкой. Что важно подчеркнуть, они начинают немедленно преследовать самца, совершенно не интересуясь самкой. Следовательно, «помехи» — просто «сбой» в системе территориальной коммуникации, определяющей распределение участков разного «качества» на току между самцами в зависимости от успешности участия последних в территориальных конфликтах друг с другом, а совсем не проявление побуждений спариваться с «чужой» самкой.
У центральных самцов, осуществляющих 30-40% копуляций, успех спариваний не коррелирует значимо с частотой внешних вторжений, у самцов со средним и низким успехом спаривания эта корреляция значима. Частота вторжений на участок самца прямо пропорциональна посещаемости территории самками и успеху спаривания хозяина: максимальное «давление» помех приходится на успешных «центральных» самцов, а их брачное поведение более устойчиво к помехам этого рода.
После прерванной (вторжением) копуляции самки чаще прерывают брачное поведение и контактируют с большим количеством самцов, чем после успешного спаривания. Каждая самка контактирует с небольшим кругом самцов на току, а спаривается только с 2-4 за несколько последовательных посещений. Самцы, регулярно вторгающиеся на соседние участки и успешно прерывающие ухаживание соседей, сами спариваются в среднем чаще (Trail, 1985; Trail, Koutnick, 1986) [9].
Следовательно, у видов, самцы которых собираются на токах, собственно взаимодействия брачных партнёров – несоциальные, «механические» или «автоматические». У моногамных видов птиц, в первую очередь певчих воробьиных функционально такие же взаимодействия «сближения и тестирования» потенциальных партнёров в процессе ухаживания истинно социальные, поскольку предполагают коммуникацию, а отклик самки на демонстрации самца оказывает важное формирующее влияние на структуру его брачной активности в последующем – так же, как и наоборот.
Так, у краснокрылого трупиала Agelaius phoeniceus самки по сравнению с самцами имеют большие коммуникативные возможности в опознании видовой песни. Самки по сравнению с самцами лучше отличали конспецифическую песню конкретного самца от песни с изменёнными начальными строфами или песни чужого самца. Распознание определяли по числу демонстраций готовности к спариванию после введения эстрадиола и предъявления песни. Опережая самцов в распознавании «типа сигнала», самки краснокрылых трупиалов сопоставимы с ними по способности узнать одну и ту же песню после разделения на три части, проигрываемые в различной последовательности (Searcy, 1990). Очевидна значимость такой способности при коммуникации в густых зарослях, где гнездятся трупиалы: и между самцами, конкурирующими за территорию, и между самцом и привлечённой самкой.
У восточной воловьей птицы Molothrus ater ater демонстрации самки способствуют формированию репертуара самца, включению в него вариантов песни, привлекательных для данной конкретной самки. В опытах использовали молодых самцов, отловленных в возрасте »50 дней, и взрослых самок (тестировали 8 пар). Песенное поведение самцов стимулируется почти любым сильным звуком или объектом (скажем, самкой канарейки). 94% песен не вызвали ответной реакции самок, но иногда те начинали демонстрационные взмахи крыльев. В ответ самцы часто прерывали пение и наблюдали за действиями самки, но достаточно часто не реагировали вовсе. Через год от 5 из 8 пар записали пять серий песен, каждая состояла из 8 песен перед взмахами крыльев, одной – одновременно со взмахами и 7 – после. При проигрывании песен самкам, готовым к откладке яиц, обнаружено, что максимальную реакцию подставления вызывают «одновременные» песни, минимальную – «последующие». До демонстраций самки самцы обычно не повторяли песню несколько раз подряд, после – повторяли от 2 до 6 раз (West, King, 1988).
Изучение локальных диалектов песни буроголовой воловьей птицы показало, что способность самца издавать сигналы в точном соответствии с диалектом прямо пропорциональна привлекательности для самки и успешности спаривания. На востоке Сьерра-Невады выделено 3 диалекта свистов, издаваемых в полёте: в центре каждой диалектной зоны частота самцов со смешанной песней 0-6%, в зонах контактов двух диалектов – 38% и 54%. Диалекты отличаются по структурным, а не фонетическим характеристикам. Не обнаружено межгодовой изменчивости диалектов, диалектный состав популяций M.a.ater стабилен на обширных территориях (Rotstein, Fleischer, 1987).
Прямой эксперимент показывает, что песенные диалекты восточной воловьей птицы формируются под управлением самки. Всего зафиксировано 12 типов песен, из которых 6 с высокой вероятностью вызывают готовность спаривания у самки (типичный ключевой стимул). Восемь молодых самцов в возрасте 30-50 дней помещали в паре с взрослой самкой той же популяции, и затем проигрывали им смесь 12 типов песен. Затем в возрасте 150 и 300 дней фиксировали их собственный песенный репертуар. В возрасте 150 дней вокализация самцов не была ещё дифференцированной на типы песен. В 300 дней 6 «привлекательных» диалектов составляли только 20% вокализации, 6 «непривлекательных» — 26%, 54% было представлено оригинальными типами песни (уже не стимул, а сигнал для межсамцовой коммуникации).
Иные результаты дал аналогичный опыт с пятью самцами воловьей птицы, которым проигрывали песни в присутствии самки канарейки. Здесь вокализация самцов дифференцировалась уже в возрасте 150 дней при явном доминировании «привлекательных» типов песни (49%). «Непривлекательные» составили 22%, неклассифицируемые вокализации – 26%. В возрасте 300 дней «привлекательные» типы песни составили 81%, «непривлекательные» — 16%. Оригинальные типы песни отсутствуют вовсе.
Следовательно, без коммуникативного взаимодействия с самкой вокализация самца насыщается стимулами, «понуждающими» к спариванию любых конспецифических самок. Однако в ней так и не формируются сигналы, демонстрация которых позволяет самке той же популяции произвести «оптимальный выбор» определённого самца по критериям, существенным для всех самок – таким, как принадлежность к местному диалекту, поведенческая стратегия, качество территории и пр. (King, West, 1989).
Именно в силу того, что коммуникативный ответ самки в процессе ухаживания имеет сильно формирующее влияние на последующие изменения песни самца (её структуры, разнообразия, темпа переключения между типами песни и пр.), в одной из работ она названа «молчаливым учителем пения». Это верно для воловьих птиц также, как для других певчих воробьиных. У тетеревов и других видов птиц, спаривающихся на току (вроде многажды упоминавшихся мной манакинов) взаимодействия самца с привлечённой самкой происходят без коммуникации, «чисто механически». В отсутствие «диалога» реакции последней никак не значимы для формирования поведения самца.
Одни и те же демонстрации (в данном случае свистовые вокализации) при разных условиях формирования и разном режиме использования в коммуникативном процессе функционируют как сигналы или ключевые стимулы, и в зависимости от этого ведут к разному результату в однотипных взаимодействиях между компаньонами. В общем, если в соответствующем процессе взаимодействия существует свобода выбора действий в проблемной ситуации и функционирует информационный обмен, способный оптимизировать этот выбор, то специфические для этого процесса структуры действий – демонстрации ли, ритуалы, функционируют как знак и специализированный посредник в устанавливающемся «диалоге» между индивидами.
Если соответствующее взаимодействие носит «механический» характер, по выражению Дьюи, те же самые специфические демонстрации оказываются просто ключевыми раздражителями или даже неспецифическими стимулами: какими именно они окажутся, зависит от обстоятельств контекста, но именно специализированным воздействием, а не сигналом, передающим некую информацию, но не «обязывающим» реагировать на неё.
Другое отличие социальных взаимодействий от несоциальных – первые оказывают формообразующее влияние на последующее поведение и внутреннее состояние животного, изменяя то и другое через последействие результатов успешной коммуникации. Вторые могут происходить с той же самой или большей частотой, напряжением и интенсивностью, но не имеют таких долговременных последствий.
Подобней всего это различие исследовано на примере агрессивных взаимодействий у «социальных» и «несоциальных» видов песчанок, близких таксономически, и обладающих очень сходной экологией. В условиях переуплотнения, обусловленного обилием предпочитаемого корма на ограниченном пространстве, «несоциальные» виды песчанок образуют столь же плотные группировки, что и социальные. И поскольку в такой ситуации контакты зверьков вынужденно часты, интенсивность агрессивных взаимодействий несоциальных видов будет не меньшей, а то и большей, чем у «социальных».
Но вот долговременные последствия агрессии у тех и других совершенно иные:
«…«социальные» песчанки формируют парные агонистические связи дольше, но их связи более асимметричны и дольше сохраняются.
Сравнивая процессы формирования агонистической парной связи у «несоциальной» светлой песчанки (Gerbillus perpallidus) и «социальной» песчанки Виноградова Meriones vinogradovi, мы обнаружили следующее. При первой встрече самцы обоих полов были очень агрессивны, причём у светлых песчанок агрессия носила «взрывной» характер и, достигнув максимума, уже на 3-ей минуте быстро спадала. Агрессия в паре социальных песчанок Виноградова медленнее развивалась и медленнее спадала. После первого выяснения отношений тех же зверьков снова ссаживали через 2-4-6-8 дней. Как и ожидалось, уровень агрессии в последующих ссаживаниях падал, но в этом случае быстрее у «социального» вида. Это свидетельствует о том, что сходные воздействия оказали на представителей «социального» вида более глубокий и стойкий эффект. (См. рисунок. В.К.)». (Попов, Чабовский,1995).
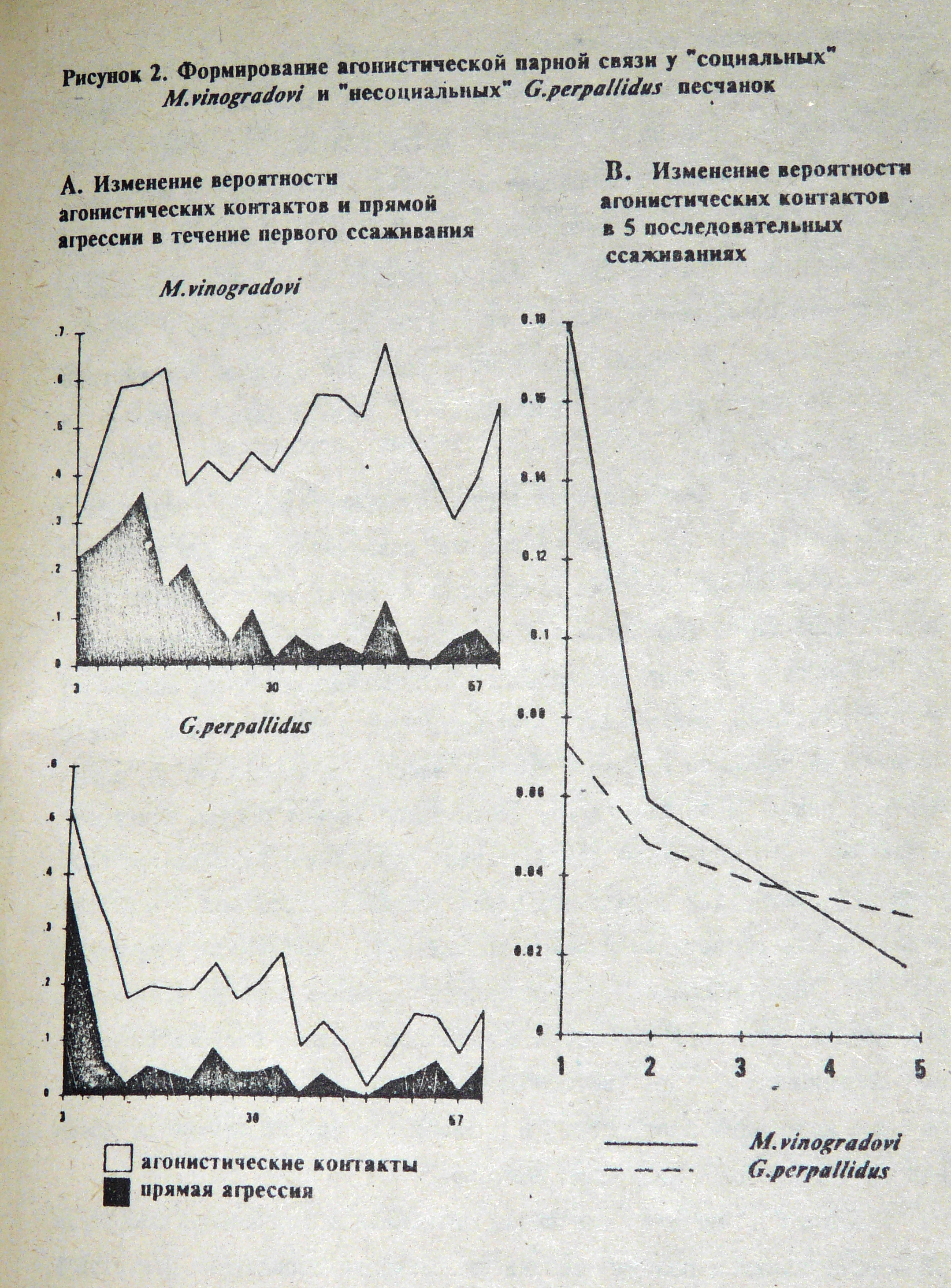
Поэтому у социальных видов агрессивные взаимодействия при меньшей интенсивности позволяют создать и поддерживать устойчивую систему иерархии в сообществе, делающую роли зверьков дифференцированными, а поведение членов сообщества – предсказуемым и друг для друга, и для внешних вселенцев.
У несоциальных видов агрессивное поведение неэффективно, также как бессмысленны и победы, достигнутые с его помощью, ибо даже существенное учащение агрессивных контактов с возрастанием напряжённости взаимодействий, большим риском ранения и т.п. отнюдь не ведёт ни к становлению устойчивой системы отношений, ни к комплементарной дифференциации ролей.
Зато они куда менее чувствительны к ранам, возникающим вследствие агрессии в условиях переуплотнения, даже к очень сильным, самки размножались иногда даже при оскальпировании. И совсем они нечувствительны к психологическому истощению и стрессу, который возникает при длительной безуспешности социальных контактов, когда частота и/или напряжение взаимодействий вышли за оптимальный уровень. А вот социальные виды тут исключительно уязвимы, то есть внутренний мир первых «пластичный», а вторых – «хрупкий».
Примечания
[1] Плюсы социобиологии и Эгоистические животные
[2] Здесь я вторгаюсь в область поведения беспозвоночных, в котором не вполне копенгаген, и если где совру, поправьте меня. В.К.
[3] См. Оптимальные сочетания у синиц, Про альтернативные стратегии разрешения проблемной ситуации, Д.А.Жуков. Нужно ли контролировать ситуацию?
[4] курсив мой – В.К.
[5] относительно общей цели – В.К.
[6] «имея в виду» их отвлечение от избранного партнёра
[7] привлечённой успешностью демонстраций первой группы и держащейся рядом с самцом
[8] Понятно, данный феномен касается выбора не только брачного партнёра, но и всех остальных отношений, существенных в социальной жизни. Главное, в нём несвобода, зависимость твоего собственного поведения от взаимодействий, в которых ты сам не участвуешь, и не можешь участвовать, ибо система сконструирована так, что не рассматривает тебя как полноценного участника. Тут сразу же вспоминается положение женщины в патриархальной семье и специфика вытекающего отсюда «счастья». В.К.
[9] Отсюда устойчивость собственных брачных демонстраций к помехам и способность создавать помехи собственной активностью – две равноправные стороны конкуренции за партнёра между самцами на току, как сеть и трезубец гладиатора на арене, щит и меч воина в сражении и т.д. Этот дуализм эффективности направленных действий самого животного с эффективным созданием помех всякому развёртыванию однотипных действий конкурента зафиксирован во всех формах социальной активности, в первую очередь – при любом обмене демонстрациями в коммуникативном процессе, как агонистическими, так предостерегающими об опасности, брачными, умиротворяющими и пр.
Тогда «помехи» в виде активных вторжений соседей именно в момент сближений территориального самца с привлечённой самкой становятся фактором отбора особей, наиболее способных к устойчивой демонстрации сигнальных инвариантов. За счёт этой устойчивости сигнализации подобные самцы наиболее привлекательны для партнёрш и одновременно наиболее эффективны в охране собственной территории, поскольку предшествующими успехами территориальной коммуникации способны минимизировать риск вторжений соседей. В.К.
Список литературы
- Попов С.В., Чабовский А.В., 1995. Плотность популяции, социальная среда и поведение: возможные взаимосвязи.// Экология популяций: структура и динамика, ч. 1, М., с. 183-202.
- Gwinner E., 1964. Untersuchungen über das Ausdruck- und Socialverhalten des Kolkraben// Z. fűr Tierpsychologie. Bd.21. S.657-748.
- Gwinner-Hanke H., 1991. Ontogenese des Socialverhaltens von Birkhühnen (Tetrao tetrix)// J. für Ornithol. Bd.132. H.2. S.121-143.
- King A., West M., 1989. Presence of female cowbirds (Molothrus ater ater) affects vocal
imitation and improvisation in males// J. Compr. Psychol. Vol.103. №1. Р.39-44. - Lorenz K., 1940. Die Paarbildung beim Kolkraben // Zeitschrift für Tierpsychologie. Bd. 3. S.278—292.
- Searcy W.A., 1990. Species recognition of song by female red-winged blackbirds// Anim. Behav. Vol.40. №6. Р.1119-1127.
- Trail P., 1985. Courtship disruption modifies mate choice in a lek-breeding birds// Science. Vol.227. №4688. Р.778-780.
- Trail P., Koutnik D., 1986. Courtship disruption at the lek in the Guianan cock-of-the-rock// Ethology. Vol.73. №3. Р.197-218.
- West M., King A., 1988. Female visual displays affect the development of male song in the cowbirds// Nature. Vol.334. №6179. Р.244-246.
Рекомендуем прочесть
 Разбор популярных мифов про охрану природы
Разбор популярных мифов про охрану природы Узники совести в Южной Корее: ликбез
Узники совести в Южной Корее: ликбез Преступники страдают от криминализации социальной среды
Преступники страдают от криминализации социальной среды Про установление классового барьера в образовании
Про установление классового барьера в образовании![Усекновение [большей] части тела у тихоходок](http://www.socialcompas.com/wp-content/uploads/2016/02/waterbear-150x150.jpg) Усекновение [большей] части тела у тихоходок
Усекновение [большей] части тела у тихоходок Урбанизация — не доместикация
Урбанизация — не доместикация Реформа образования в Мексике для «чайников»
Реформа образования в Мексике для «чайников» Восприятие расы в США
Восприятие расы в США
