
Д.А.Кишкинев
Миллионы птиц каждый год совершают миграции, порой пересекая тысячи километров над сушей и морем. Более того, представители многих видов мигрируют в ночное время, часто в одиночку или рыхлыми непостоянными группами. Известно, что птицы могут возвращаться в район зимовок или к гнезду даже после смещения в очень удаленные районы, которые они ранее никогда не посещали (Perdeck 1958; Mewaldt 1964). Другими словами, птицы способны к настоящей навигации.
Каким образом птицы навигируют? Какие сенсорные механизмы для этого используются? Эти вопросы ученые задают уже не первое столетие. Исследования последних лет еще не принесли окончательных ответов на данные вопросы, но существенно продвинули наше понимание данной проблемы.
Ниже я кратко рассмотрю наиболее значимые результаты, достигнутые за последние годы.
Навигация молодых птиц во время первой миграции
Каким образом молодые птицы находят дорогу к местам зимовок, которые до этого никогда не посещали? Проведенные до недавнего времени исследования показывают, что ответ на этот вопрос зависит от конкретного вида птиц.
Молодые птицы тех видов, которые мигрируют стаями или образуют скопления (например, паря в термальных потоках воздуха), могут следовать за взрослыми, параллельно запоминая путь. Такую стратегию используют «социальные» мигранты, например, аисты, гуси, кулики, некоторые хищные птицы. Вероятнее всего, молодые птицы запоминают ландшафтные ориентиры и их последовательность уже во время первой миграции. Запомнив маршрут, повзрослевшие птицы способны очень точно следовать одному и тому же коридору миграции (Berthold et al 2004). Что произойдет, если молодые птицы не могут следовать за взрослыми во время первой миграции, например, если их искусственно задержали на месте рождения и все взрослые птицы покинули территорию или же если молодых птиц переместили в район, находящийся вне ареала данного вида? Данные, полученные в эксперименте, показывают, что молодые белые аисты во время осенней миграции, перемещенные в удаленный район, где они не могли встретить особей своего вида, всё же могут следовать в примерном миграционном направлении (Chernetsov et al 2004).
При этом разброс направлений осенней миграции у молодых аистов, летящих без взрослых, намного выше по сравнению с ситуацией, когда они летят со старшими (Berthold et al 2004; Chernetsov et al 2004). По-видимому, молодые белые аисты обладают врожденной программой «лететь примерно в южном направлении» (Chernetsov et al 2004).
Как добираются до зимовок молодые воробьиные мигранты, летящие без родителей? Известно, что многие молодые певчие птицы часто летят на зимовки позднее взрослых, при этом многие виды (например, представители семейств Sylviidae, Muscicapidae, Turdidae, Parulidae) мигрируют ночью и в одиночку или образуя рыхлые нестабильные скопления. Исследования 1930–1980-х гг. легли в основу так называемой гипотезы «часов и компаса», или векторной навигации (Drost 1938; Perdeck 1958, 1967; Mewaldt 1964; Gwinner 1968; Berthold and Querner 1981, 1982; Berthold 1988; Berthold et al. 1990).
Согласно этой гипотезе, молодая птица имеет врожденную генетическую программу, которую можно описать следующим образом: «Следуй в данном генетически заданном компасном направлении α в течение X дней, затем смени направление на β и продолжай путь в течение еще Y дней, после чего остановись, так как, скорее всего, это район зимовки». Подтверждением данной гипотезы является то, что молодые птицы в отличие от взрослых не способны компенсировать географические смещения, то есть после смещения они продолжают следовать в миграционном направлении, которое уже не ведет к зимовкам (Perdeck 1958, 1967; Thorup et al. 2007). Это значит, что молодые птицы не имеют представления о своем географическом положении и следуют некому заданному курсу. Взрослые же птицы способны компенсировать смещение, проявляя так называемую истинную навигацию (Perdeck 1958, 1967; Thorup et al. 2007). Подтверждением того, что направление осенней миграции у молодых птиц задано генетически, являются эксперименты со скрещиванием птиц из разных популяций с разными врожденными миграционными направлениями (Helbig 1991), хотя гены, определяющие направление миграции, еще не найдены и являются предметом современных исследований (Liedvogel et al. 2011). Интересно, что эволюция наследственной ориентационной программы у того или иного вида может происходить относительно быстро, т.е. в течение нескольких десятилетий. К примеру, с 1970-х гг. доля славок-черноголовок, летающих из континентальной Европы на новые места зимовок в Великобританию, а не в традиционные зимовочные места на Иберийском полуострове, резко возросла (Fiedler et al. 2003), что, по-видимому, связано с преимуществом генов северо-западной осенней миграции над генами юго-западной миграции.
Каким образом молодые птицы понимают, когда нужно начинать и когда заканчивать миграцию? Вероятнее всего, базовые временные рамки миграции задаются эндогенными генетическими механизмами на основе циркануальных часов (от лат. circa – «около», annual – «годовой»).
Биологические часы могут многие годы идти независимо от внешних стимулов, однако довольно быстро рассинхронизируются с годовыми событиями. Для синхронизации биологических часов птицы используют различные внешние стимулы, главным из которых является продолжительность светового дня (Gwinner 1986, 1996 для обзора). Генетическая основа времени миграции была показана в ряде экспериментов. В одном из них методом искусственной селекции из группы славок-черноголовок из Южной Германии, имеющих недолгий период так называемого миграционного беспокойства – поведенческого синдрома у мигрирующих птиц, содержащихся в помещении, и заменяющего им миграционный полет – вывели линию «дальних мигрантов» (длинный период миграционного беспокойства) и «резидентов» (короткий период миграционного беспокойства) (Berthold et al. 1990). Кроме того, при скрещивании славок-черноголовок из популяций, удаленных на разное расстояние от африканских зимовок, потомки демонстрируют промежуточный уровень миграционного беспокойства по сравнению с родителями (Berthold and Querner 1981). Всё это указывает на то, что существуют гены, определяющие длительность миграции. Недавние генетические исследования указывают на ряд конкретных генов, которые скоррелированы с продолжительностью некоторых стадий годового цикла у птиц (Liedvogel et al. 2012).
Однако гены определяют лишь грубые временные рамки, в которые происходит миграция. Точная дата отлета и прилета часто выбирается на основе местных погодных и кормовых условий (Ramenofsky and Wingfield 2007; Mitchell et al. 2012). При этом дальние мигранты больше полагаются на эндогенные механизмы и синхроннее улетают из мест рождения и размножения и прилетают в места зимовок, нежели ближние мигранты, время миграции которых в большей степени модулируется местными погодными и кормовыми условиями (см. Newton 2008 для обзора). Механизм терминации миграции еще довольно плохо изучен. Вероятнее всего, при этом также общие рамки задаются динамикой физиологических процессов, находящихся под контролем генов. Другими словами, птица перестает мигрировать, когда ее миграционная программа остановится. Однако точная дата завершения миграции определяется внешними условиями (погода, корм, социальные контакты).
Здесь следует упомянуть, что довольно часто птицы среди зимы вновь переходят в миграционное состояние и совершают зимнюю миграцию, порой в удаленный район (тысячи км), после чего завершают зимовку уже в новом районе (Newton 2008 для обзора).
Достаточно ли точно описывает миграцию молодых птиц гипотеза «часов и компаса»? В частности, имеют ли молодые птицы во время миграции хотя бы приблизительное представление, в каком районе они находятся? За последние 10–0 лет накопились данные, указывающие на то, что молодые мигранты в определенных условиях могут определять свое положение на трассе миграции или используют какие-то дополнительные механизмы, повышающие точность первой миграции к местам зимовок (Thorup and Rabø 2001; Gschwenk et al. 2009).
В частности, если предположить, что молодые птицы «знают» только генетически заданное направление, то по мере удаления от мест рождения разброс индивидуальных направлений должен постоянно возрастать, причем разброс у молодых птиц должен быть больше, чем у взрослых, уже знакомых с маршрутом. Такое предположение подтверждается некоторыми исследованиями (Mouritsen and Mouritsen 2000). Однако районы зимовок у некоторых видов настолько малы, что при постоянно возрастающем разбросе направлений до зимовок добиралась бы очень небольшая доля вылетевших с мест рождения молодых птиц, чего на самом деле не происходит (Thorup and Rabø 2001). Недавние данные спутниковой телеметрии напрямую показывают, что кроме участков миграции, где разброс индивидуальных направлений возрастает, есть промежуточные участки миграции, где разброс направлений уменьшается (Gschweng et al. 2009).
Каким образом молодые птицы концентрируются в некоторых районах, лежащих далеко от мест зимовок, еще предстоит выяснить. Возможно, что молодые птицы используют некоторые врожденные «правила», чтобы уменьшить разброс миграционных направлений. Например, существуют данные о том, что птицы предпочитают следовать вдоль определенных географических направляющих, если они не сильно отклоняют птицу с маршрута. Такими направляющими могут быть береговые линии (т.н. береговой эффект; Meyer et al. 2003) или горные хребты.
Кроме того, последние годы всё больше данных указывают на то, что молодые мигранты не совсем лишены навигационных способностей и способны грубо определять свое местоположение на трассе миграции, используя те или иные природные координаты (астроориентиры, параметры геомагнитного поля, ольфакторные ориентиры: Fransson et al. 2001; Kullberg et al. 2003; Thorup et al. 2011). Возможно, что для этого птицам не нужно иметь «врожденный глобальный атлас», а достаточно реагировать на некоторые стимулы или комбинацию таковых, как только они встретятся на трассе миграции (т.н. «дорожные знаки» –signposts). К примеру, данные, полученные на шведских обыкновенных соловьях, указывают на то, что магнитные параметры, соответствующие Северному Египту, вызывают резкий рост жиронакопления, в то время как соловьи, помещенные в магнитное поле Швеции, такого резкого роста жиронакопления не демонстрируют (Fransson et al. 2001). Возможно, что магнитное поле Египта выступает своего рода триггером жиронакопления, указывающим молодым птицам на то, что впереди лежит широкий экологический барьер –пустыня Сахара, который лучше всего пересечь быстро, для чего нужно запастись «топливом».
Сенсорные и когнитивные механизмы навигации
Как уже было сказано выше, исследования показали, что взрослые мигранты, начиная с первой весенней миграции, способны компенсировать смещение, т.е. определять свое положение относительно цели и следовать в направлении, ведущем к данной цели. Другими словами, взрослые мигранты способны навигировать. Какими способами можно осуществлять навигацию? Рассмотрим некоторые из них.
Если вы бродите по лесу в поисках грибов вокруг поляны, где вас ждет оставленная машина, то скорее всего вы используете векторную интеграцию, т.е. рассчитываете направление и расстояние до «цели» на основе уже пройденного расстояния и совершенных поворотов. Такой способ позволяет в любой момент времени определять, куда и как долго нужно идти, чтобы примерно выйти на цель. Однако по мере удаления от цели и совершения всё большего количества поворотов, ошибка векторной интеграции возрастает.
Исследования показывают, что данный метод навигации широко используют насекомые (например, муравьи рода Cataglyphis: Wittlinger et al. 2006; и медоносные пчелы; Cruse and Wehner 2011), а из позвоночных – грызуны (Maaswinkel and Whishaw 1999). Птицы, по-видимому, также способны к векторной интеграции, но только при небольших локальных кормовых перемещениях. Почтовые голуби, завезенные далеко от голубятни на постоянно вращающихся платформах, способны возвращаться домой, т.е. совершать т.н. хоминг (от англ. homing) с такой же точностью и скоростью, как те особи, которых завозили без вращения (Wallraff 1980; Wallraff et al. 1980). Таким образом, векторная интеграция скорее всего не используется птицами при навигации на большие расстояния.
Следование по цепочке знакомых ориентиров называется пилотированием. Данный способ нахождения цели был показан в ряде экспериментов по хомингу почтовых голубей (Biro et al. 2002, 2004). Вполне возможно, что именно пилотированием объясняются очень узкие коридоры миграции у белых аистов (Chernetsov et al. 2004). Однако многочисленные эксперименты с завозами почтовых голубей и диких мигрантов показывают, что птицы способны находить дом, будучи завезенными далеко вглубь незнакомой территории (Perdeck 1958; Thorup et al. 2007; см. Wallraff 2006 для обзора данных по хомингу голубей), что исключает использование пилотирования, по крайней мере, на большей части пути. В этих случаях предполагают, что птицы используют истинную навигацию.
Термин истинная навигация используется для случаев нахождения направления, ведущего к цели, даже из незнакомых районов, где невозможно прямое восприятие идущей от цели информации. Другими словами, если птица сильно удалилась от знакомого маршрута миграции или от места гнездования, или же ее завезли в рамках эксперимента, так что из данного района она не ощущает знакомых ориентиров (зрительных, запаховых, акустических и прочих), и при этом птица способна определить свое местоположение и курс, ведущий к цели, то мы имеем дело с истинной навигацией. Говоря об истинной навигации, исследователи часто подразумевают, что птицы, подобно людям, находят свои координаты, одна из которой примерно соответствует широте (т.е. положению вдоль оси север – юг), а вторая –долготе (положению вдоль оси запад –восток). Понимая антропоморфизм данных размышлений, попытаемся предположить, какие внешние стимулы могли бы использоваться птицами для определения данных координат, и обязательно ли птицы используют две координаты для навигации.
Известно, что техника определения широты намного проще, нежели определение долготы, т.к. для первого есть большее количество внешних стимулов. К примеру, для определения широты можно использовать высоту Полярной звезды (в северном полушарии) или, в общем, любого центра вращения звездного неба. Кроме того, можно использовать высоту Солнца в местный полдень. Напряженность и инклинация геомагнитного поля также позволяет грубо определить широтное положение. Возможности же определения долготы сильно ограничены. Если не учитывать искусственных сигналов, как-то сигналы наземных и спутниковых навигационных систем, единственным глобальным способом определения своего долготного положения является определение местного времени и сравнение его с неким референтным временем. Зная разницу между ними, можно определить удаленность местного меридиана от референтного. Таким образом, нужно уметь точно измерять местное время и всегда иметь с собой источник референтного времени (например, носить с собой точные часы, установленные на Гринвичское время). Именно из-за этой сложности люди решили «проблему долготы» довольно поздно по историческим меркам –лишь в XVIII в. английский инженер Гаррисон изобрел хронометр, достаточно точно идущий на протяжении месяцев, несмотря на перепады в давлении, температуре и влажности. Такой хронометр можно было использовать в качестве хранителя референтного времени.
Если долготу так сложно измерить, используют ли ее птицы? Датский исследователь Хенрик Моуритсен предположил, что навигация птиц монокоординатная, так как птицы измеряют только широту (Mouritsen 2003). Более того, данная гипотеза предполагала, что большую часть времени мигрирующие птицы используют лишь компасные направления. Птице достаточно держать примерный курс из района гнездования к промежуточным миграционным остановкам и далее зимовкам для того, чтобы выйти на знакомые зрительные ориентиры вокруг этих целей. Весной птица летит в район гнездования, зная только широту данного района. После достижения данной широты птица останавливается и начинает поисковые движения вдоль широты, стараясь выйти на район визуально знакомых ориентиров (Mouritsen 2003). Данная гипотеза была недавно проверена в эксперименте двумя независимыми группами исследователей (Thorup et al. 2007, Chernetsov et al. 2008). В обоих экспериментах птицы были перемещены с трасс миграции вдоль оси запад–восток на значительные расстояния (более 1000 км), после чего их ориентация была проверена на новом месте. Данные, полученные в ходе проведения этих экспериментов, четко показывают, что мигрирующие птицы способны определять свое долготное положение. Каким же образом птицы способны определять долготу? Используют ли для этого птицы какие-либо внутренние часы?
Использование птицами внутренних биологических ритмов было давно показано в многочисленных экспериментах (см. Gwinner 1986, 1996 для обзора). Однако ни у птиц, ни у иных животных не было показано существование часов, не синхронизируемых внешними стимулами, которые могли бы играть роль референтных хронометров. Самым известным синхронизатором является смена света и темноты (фотопериод). Возможно ли использование биологических часов для определения долготы, если несинхронизируемый хронометр в организме птиц до сих пор не был обнаружен? Недавно была выдвинута гипотеза т.н. «двойных часов», которая пыталась объяснить навигацию птиц по долготе без предположения о существовании несинхронизируемого хронометра, а на основе уже найденных биологических часов (Kishkinev et al. 2010).
Исследования в области изучения биологических ритмов показывают, что при резком смещении фотопериода наблюдается временная десинхронизация внутренних осцилляторов. Именно этим объясняется явление джетлэга (jetlag) при быстром пересечении нескольких временных поясов Земли (например, трансатлантический перелет). На клеточном уровне происходит следующее: в первый день резкого изменения фотопериода пик электрической активности в нейронах супрахиазматического ядра мозга («дирижер» биологических ритмов у млекопитающих и важный осциллятор у птиц) резко десинхронизируется с ритмом света и темноты. В следующие дни разные популяции «часовых» нейронов ведут себя по-разному. Вентральная популяция нейронов (нейроны «быстрых часов») быстро подстраивается к новому времени, в то время как дорзальной популяции (нейроны «медленных часов») на это требуется несколько дней (de la Iglesia et al. 2004; Piggins and Loudon 2005). Чем больше первоначальная разница между «старым» и «новым» фотопериодами, тем дольше занимает синхронизация между «быстрыми» и «медленными» популяциями. В гипотезе двойных часов было предположено, что время синхронизации «медленных» нейронов с «быстрыми» может быть основой долготной навигации. Однако экспериментальная проверка данной гипотезы показала, что скорее всего мигрирующие птицы не используют механизм двойных часов для определения своей долготы (Kishkinev et al. 2010).
Таким образом, механизм определения птицами своего географического положения еще предстоит выяснить. Исследования последних десятилетий указывают на то, что на роль координат могут претендовать, по крайней мере, два естественных стимула: запахи и магнитное поле Земли. Эксперименты с почтовыми голубями, а также трубконосыми, достаточно убедительно показывают, что эти птицы используют запахи, а не геомагнитное поле (Benhamou et al. 2003; Bonadonna et al 2003; Mouritsen et al. 2003; Bonadonna et al. 2005). В то же время тщательной проверки гипотезы ольфакторной навигации у других групп птиц, в частности, у воробьиных мигрантов, еще не проводилось, а существующие немногочисленные экспериментальные данные пока неубедительны (Holland et al. 2009). Более того, имеется ряд работ, показывающих важность магнитного поля для определения положения у воробьиных (Fransson et al. 2001; Fischer et al. 2003; Kullberg et al. 2003, 2007; Deutschlander et al. 2012).
Поиск органа магнитной навигации еще не завершился.
В поисках органа магнитной навигации
В начале 2000-х годов группа профессора Фляйснера сообщила о находке магниторецепторов, содержащих оксиды железа (минералы магнетит и магемит), в надклювье у почтовых голубей (Fleissner et al. 2003, 2007). Лаборатория Фляйснера описал железосодержащие дедритные окончания, ассоциированные со зрительной ветвью тройничного нерва (Fleissner et al. 2003, 2007). Авторы находки назвали совокупность железодержащих нервных окончаний «клювным органом» и предположили, что данный орган предназначен для магнитной навигации (Fleissner et al. 2003, 2007). Впоследствии та же исследовательская группа сообщила о находке клювного органа у других групп птиц (курообразные, воробьинообразные: Falkenberg et al. 2010).
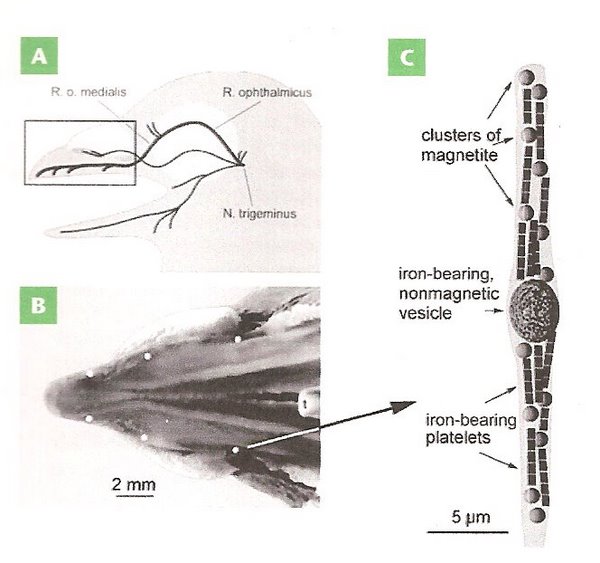
Для чего на самом деле нужен предполагаемый клювный орган? Работа, выполненная в лаборатории Хенрика Моуритсена, показала, что информация, передаваемая по зрительной ветви тройничного нерва, не нужна для функционирования магнитного компаса у обыкновенной зарянки (Zapka et al. 2009). Таким образом, если найденные Фляйснером клетки воспринимают магнитную информацию и передают ее в мозг, используется она не для определения направления. Передается ли вообще магнитная информация по данному нерву? Имеются несколько независимых исследований, подтверждающих, что какая-то магнитная информация передается по зрительной ветви тройничного нерва (Beason et al. 1995; Mora et al. 2004; Heyers et al. 2010). Так как данная ветвь иннервирует переднюю часть головы и надклювье, магниторецепторы нужно искать в этой части головы. Но действительно ли группа Фляйснера нашла магниторецепторы? Недавняя попытка повторить исследования Фляйснера показала, что в надклювье голубей действительно много железосодержащих клеток, но подавляющее большинство данных клеток –макрофаги (Treiber et al. 2012). Однако утверждать, что магниторецепторов в надклювье у птиц нет, пока еще рано, так как имеются факты, подтверждающие, что зрительная ветвь тройничного нерва передает магнитную информацию в мозг (Beason et al. 1995; Mora et al. 2004; Heyers et al. 2010). Поиски магниторецепторов в надклювье, а также установление функции предполагаемых магниторецепторов продолжаются.
Выводы и перспективы
Птицы, имеющие опыт миграции, способны к истинной навигации. Птицы способны определять свое местоположение как по широте, так и по долготе. Механизм такой бикоординатной навигации еще не ясен. Определение долготы скорее всего не основано на биологических часах. Последние данные указывают на то, что разные группы птиц могут использовать разные внешние стимулы для навигации. В частности, почтовые голуби и трубконосые, вероятнее всего, используют запахи, а воробьиные мигранты –магнитное поле Земли.
Последние работы указывают на то, что в клюве птиц имеются магниточувствительные клетки, иннервированные тройничным нервом, хотя сами клетки пока являются предметом активного поиска.
Литература
1. Beason RC, Dussourd N, Deutschlander ME (1995) Behavioural evidence for the use of magnetic material inmagnetoreception by a migratory bird. J Exp Biol 198:141–46.
2. Benhamou S, Bonadonna F, Jouventin P (2003) Successful homing of magnet-carrying white-chinned petrels released in the open sea. Anim Behav 65:729–34.
3. Berthold P (1988) Evolutionary aspects of migratory behavior in European warblers. J Evol Biol 1:195–09.
4. Berthold P, Kaatz M, Querner U (2004) Long-term satellite tracking of white stork (Ciconia ciconia) migration: constancy versus variability. J Ornith 145: 356–59.
5. Berthold P, Wiltschko W, Miltenberger H, Querner U (1990) Genetic transmission of migratory behavior into anon-migratory bird population. Experientia 46:107–08.
6. Berthold P, Querner U (1981) Genetic basis of migratory behavior in European warblers. Science 212:77–9.
7. Berthold P, Querner U (1982) Genetic basis of moult, wing length, and body weight in a migratory bird species, Sylvia atricapilla. Experientia 38:801–02.
8. Biro D, Guilford T, Dell’Omo G, Lipp H-P (2002) How the viewing of familiar landscapes prior to release allows pigeons to home faster: evidence from GPS tracking. J Exp Biol 205: 3833–844.
9. Biro D, Meade J, Guilford T (2004) Familiar route loyalty implies visual pilotage in the homing pigeon. Proc Natl Acad Sci USA 101:17440–7443.
10. Bonadonna F, Bajzak C, Benhamou S, Igloi K, Jouventin P, Lipp HP, Dell’Omo G.140 (2005) Orientation in the wandering albatross: interfering with magnetic perception does not affect orientation performance. Proc Biol Sci. 272:489–95.
11. Bonadonna F, Chamaille-Jammes S, Pinaud D, Weimerskirch H (2003) Magnetic cues: are they importantin black-browed albatross Diomedea melanophris orientation? Ibis 145:152–55.
12. Chernetsov N, Berthold P, Querner U (2004) Migratory orientation of first-year white storks (Ciconia ciconia): inherited information and social interactions. J Exp Biol 207:937–43.
13. Chernetsov N, Kishkinev D, Mouritsen H (2008) A long-distance avian migrant compensates for longitudinal displacement during spring migration. Curr Biol 18:188–90.
14. Cruse H, Wehner R (2011) No need for a cognitive map: decentralized memory for insect navigation. PLoS Comput Biol 7:e1002009.
15. de la Iglesia HO, Cambras T, Schwartz WJ, Díz-Noguera A (2004) Forced desynchronization of dual circadian oscillators within the rat suprachiasmatic nucleus. Curr Biol 14:796–00.
16. Deutschlander ME, Phillips JB, Munro U (2012) Age-dependent orientation to magnetically-simulated geographical displacements in migratory Australian Silvereyes (Zosterops l. lateralis). Wils J Ornith 124:467–77.
17. Drost R (1938) Üer den EinflussvonVerfrachtungen zur Herbstzugzeiten auf den Sperben, Accipiter nisus(L.). Zugleichein Beitrag zur Frage nach der Orientierung der Vöel auf dem Zuge insWinterquartier. In: ProcIntOrnitholCongr(Rouen 1938). 9:502–21.
18. Falkenberg G, Fleissner G, Schuchardt K, Kuehbacher M, Thalau P, Mouritsen H, Heyers D, Wellenreuther G, Fleissner G (2010) Avian magnetoreception: elaborate iron mineral containing dendrites in the upper beak seem to be a common feature of birds. PLoS One 5:e9231.
19. Fiedler W (2003) Recent changes in migratory behaviour of birds: a compilation of field observation and ringing data. In: Avian Migration (Berthold P, Gwinner E, Sonnenschein E. eds.). Berlin, Heidelberg, Springer Verlag, 21–8.
20. Fischer JH, Munro U, Phillips JB (2003) Magnetic navigation by an avianmigrant? In: Avian Migration. (Berthold P, Gwinner E, Sonnenschein E. eds.). Berlin, Heidelberg, Springer Verlag, 423–32.
21. Fleissner G, Holtkamp-Rözler E, Hanzlik M, Fleissner G, Petersen N, Wiltschko W (2003) Ultrastructural analysis of a putative magnetoreceptor in the beak of homing pigeons. J Comp Neurol 458:350–60
22. Fleissner G, Stahl B, Thalau P, Falkenberg G, Fleissner G (2007) A novel concept of Fe-mineral-based magnetoreception: histological and physicochemical data from the upper beak of homing pigeons. Naturwiss 94:631–42.
23. Fransson T, Jakobsson S, Johansson P, Kullberg C, Lind J, Vallin A (2001) Magnetic cues trigger extensive refuelling. Nature 414: 35–6.
24. Gschweng M, Kalko EKV, Querner U, Fiedler W, Berthold P (2008) All across Africa: highly individual migration routes of Eleonora’s falcon. Proc Biol Sci 275:2887–896.141
25. Gwinner E (1968) Circannuale Periodikals Grundlage des jahreszeitlichen Funktionswandels bei Zugvöeln. Untersuchungen am Fitis (Phylloscopus trochilus) und am Waldlaubsäger (P. sibilatrix). J Ornithol 109:70–5.
26. Gwinner E (1986) Circannual rhythms: endogenous annual clocks in the organization of seasonal processes. Berlin, Springer Verlag.
27. Gwinner E (1996) Circadian and circannual programmes in avian migration. J Exp Biol 199:39–8.
28. Hake M, Kjelle N, Alerstam T (2001) Satellite tracking of Swedish Ospreys Pandion haliaetus autumn migration routes and orientation. J Avian Biol 32:47–6.
29. Helbig AJ (1991) Inheritance of migratory direction in a bird species: across-breeding experiment with SE- and SW-migrating blackcaps (Sylvia atricapilla). Behav Ecol Sociobiol 28:9–2.
30. Heyers D, Zapka M, Hoffmeister M, Wild JM, Mouritsen H (2010) Magnetic field changes activate the trigeminal brainstem complex in a migratory bird. Proc Natl Acad Sci USA 107:9394–399.
31. Holland RA, Thorup K, Gagliardo A, Bisson IA, Knecht E, Mizrahi D, Wikelski M. (2009) Testing the role of sensory systems in the migratory heading of a songbird. J Exp Biol. 212:4065–071.
32. Kishkinev D, Chernetsov N, Mouritsen H (2010) A double clock or jetlag mechanism is unlikely to be involved in detection of east-west displacement in a long-distance avian migrant. Auk 127:773–80.
33. Kullberg C, Henshaw I, Jakobsson S, Johansson P, Fransson T (2007) Fuelling decisions in migratory birds: geomagnetic cues override the seasonal effect. Proc R Soc B 274:2145–151.
34. Kullberg C, Lind J, Fransson T, Jakobsson S, Vallin A (2003) Magnetic cues and time of season affect fuel deposition in migratory thrush nightingales (Luscinia luscinia). Proc R Soc Lond B 270: 373–78.
35. Liedvogel M, Åesson S, Bensch S (2011) The genetics of migration on the move. TREE 26:561–69.
36. Liedvogel M, Cornwallis CK, Sheldon BC (2012) Integrating candidate gene and quantitative genetic approaches to understand variation in timing of breeding in wild tit populations. J Evol Biol 25:813–23.
37. Maaswinkel H, Whishaw IQ (1999) Homing with locale, taxon, and deadreckoning strategies by foraging rats:sensory hierarchy in spatial navigation. Behav Brain Res 99:143–52.
38. Mewaldt R (1964) California sparrows return from displacement to Maryland. Science146:941–42.
39. Meyer SK, Spaar R, Bruderer B (2003) Sea crossing behaviour of falcons and harriers at the southern Mediterranean coast of Spain. Avian Science 3:153–62.
40. Mitchell G, Newman AEM, Wikelski M, Norris R (2012) Timing of breeding carries over to influence migratorydeparture in a songbird: an automated radiotracking study. J Anim Ecoldoi: 10.1111/j.1365–656.2012.01978.x.142
41. Mora CV, Davison M, Wild JM, Walker MM (2004) Magnetoreception and its trigeminal mediation in the homing pigeon. Nature 432: 508–11.
42. Mouritsen H, Huyvaert K P, Frost BJ, Anderson DJ (2003) Waved albatrosses can navigate with strong magnets attached to their head. J Exp Biol 206:4155–166.
43. Mouritsen H, Mouritsen O (2000) A mathematical expectation model for bird navigation based on the clock-and-compass strategy. J Theor Bio l207:283–91.
44. Mouritsen H. (2003) Spatiotemporal orientation strategies of long-distance migrants. In: Avian Migration. (Berthold P, Gwinner E, Sonnenschein E, eds.). Berlin, Heidelberg, Springer Verlag, 493–13.
45. Newton I (2008) The migration biology of birds. London, Academic Press, 479– 522.
46. Perdeck AC (1958) Two types of orientation in migrating Sturnus vulgaris and Fringilla coelebs as revealed by displacement experiments. Ardea 46:1–7.
47. Perdeck AC (1967) Orientation of starlings after displacement to Spain. Ardea55:194–02.
48. Piggins HD, Loudon A (2005) Circadian biology: clocks within clocks. Curr Biol 5:R455–457.
49. Ramenofsky M, Wingfield JC (2007) Regulation of migration. Bioscience 57:135– 143.
50. Thorup K, Bisson IA, Bowlin MS, Holland RA, Wingfield JC, Ramenofsky M, Wikelski M (2007) Evidence for a navigational map stretching across the continental U.S. in a migratory songbird. Proc Natl Acad Sci USA 104:18115–8119.
51. Thorup K, Ortvad TE, Rabø J, Holland RA, Tøtrup AP, Wikelski M (2011) Juvenile songbirds compensate for displacement to oceanic islands during autumn migration. PLoS One 25:e17903.
52. Thorup K, Rabø J (2001) The orientation system and migration pattern of long distance migrants: conflict between model predictions and observed patterns. J Avian Biol 32:111–19.
53. Treiber CD, Salzer MC, Riegler J, Edelman N, Sugar C, et al. (2012) Clusters of iron-rich cells in the upper beak of pigeons are macrophages not magnetosensitive neurons. Nature 484:367-370.
54. Wallraff H (1980) Does pigeon homing depend on stimuli perceived during displacement? I. Experiments in Germany. J Comp Physiol 139:193–01.
55. Wallraff H (2006) Avian Navigation: Pigeon Homing as a Paradigm. Berlin, Springer, 242 p.
56. Wallraff H, Foà A, Ioa1éP (1980) Does pigeon homing depend on stimuli perceived during displacement? II. Experiments in Italy. J Comp Physiol139: 203–08.
57. Wittlinger M, Wehner R, Wolf H (2006) The ant odometer: stepping on stilts and stumps. Science 312:1965–967.
58. Zapka M, Heyers D, Hein CM, Engels S, Schneider N-L, Hans J, Weiler S, Dreyer D, Kishkinev D, Wild M, Mouritsen H (2009) Visual, but not trigeminal, mediation of magnetic compass information in a migratory bird. Nature 461:1274–277.143
«Бутурлинский сборник». Матералы 4-х Международных Бутурлинских чтений. Ульяновск, 2012. С.131-137.
Рекомендуем прочесть
 «Лето Свободы» отмечает 50-летие
«Лето Свободы» отмечает 50-летие Малоизвестная победа короля Фейсала над коммунизмом и СССР
Малоизвестная победа короля Фейсала над коммунизмом и СССР Развенчивание мифов. Подробности убийства Китти Дженовезе сильно преувеличены
Развенчивание мифов. Подробности убийства Китти Дженовезе сильно преувеличены «Левиафан»
«Левиафан» Высокие цены на продовольствие не зависят от урожая
Высокие цены на продовольствие не зависят от урожая Жизнь гастарбайтеров в ОАЭ
Жизнь гастарбайтеров в ОАЭ РПЦ «укрепит единство нации» за бюджетные деньги
РПЦ «укрепит единство нации» за бюджетные деньги  Школьная травля: анализ социальных факторов
Школьная травля: анализ социальных факторов
