

Между «долгими криками» двух видов поморников сохраняются устойчивые различия: 1–5 — последовательные фазы долгого крика, столбики внизу — частота встречаемости у того и другого вида. (По:. Pietz P., «Condor», 1985, т. 81).
Аннотация. В социальном поведении животных важно различать, часто и противопоставлять друг другу два аспекта, которые можно условно назвать «структура» и «наполнение». Первое задаёт специфику формы данного поведения; последнее распознаётся как таковое социальными компаньонами и отреагируется «как должно» именно по специфическим характеристикам формы, а не эффекта. Второе — специфическая энергия действия, без определённого уровня которой соответствующее поведение не может ни удерживаться, ни быть действенным.
«Структура» и «наполнение» поведение как форма и энергия действия
В социальном поведении животных важно различать (а часто и противопоставлять друг другу) два аспекта, которые можно условно назвать «структура» и «наполнение». «Структура» — это форма и схема организации некоей поведенческой последовательности, скажем, связанной с патрулированием территории или привлечением самки, или ухаживанием между партнёрами в образовавшейся паре и т.п. Эти структуры поведения (именно в силу наличия специфической организации) обычно узнаваемы, типизируемы, легко дифференцируются от «фона» повседневного поведения и от других таких же единиц, то есть легко характеризуются с точки зрения «морфологии поведения». Так сказать, типичные «врождённые формы реагирования» в типических обстоятельствах взаимодействия особи со своими компаньонами (а если обстоятельства нетипичные, то поведение дестабилизируется, и никакой «аномальной» структуры мы выделить не можем, а видим отсутствие структуры).
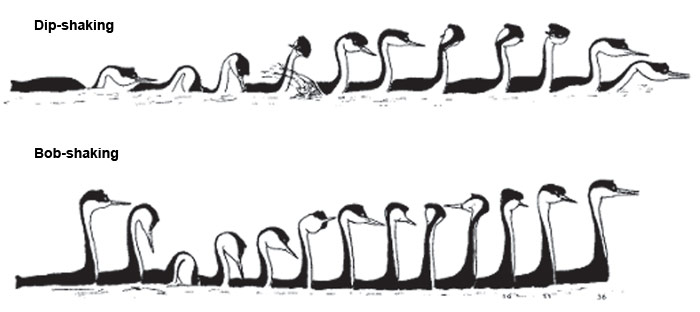
Демонстрации ухаживания «dip-shaking» и «bob-shaking» у водоплавающей птицы западной поганки очень сходны, но имеют отличия в ключевых моментах, в начале и конце. Этого достаточно, чтобы их визуальные образы четко различались. (Из: Nuechterlein С, Storer R., «Condor», 1982, т. 84. 4)
Ну, а «наполнение» — это специфика внутреннего состояния животного (эмоционального, гормонального, мотивационного или какого-то ещё), поддерживающее реализацию данной структуры, часто вопреки сопротивлению противника и прочим помехам, или служащее той специфической внутренней средой, при которой реализация данного поведения наиболее вероятна. В этом случае говорят о мотивационной подоснове поведения: скажем, реализация разных угрожающих демонстраций озёрной чайки связана с разным соотношением побуждений к нападению (тёмные столбцы) и к избеганию (светлые столбцы), т.е. с разным уровнем агрессивной мотивации, образованной конфликтом этих элементарных стремлений. Или см.структуру «долгого крика» сизой чайки здесь, стр.12
Другой пример: территориальное поведение певчих птиц обычно опосредовано повышенным содержанием тестестерона, так те территориальные самцы, которые наиболее эффективно охраняют то индивидуальное пространство, в котором они поют и куда привлекают самок, имеют повышенное содержание тестостерона, причём часто степень повышения пропорциональна превосходству в эффективности охраны.
Это и многие другие наблюдения подталкивают к выводу о достаточно точном соответствии поведения и состояния животного. Мол, всякой специфической модели поведения (специфичной благодаря форме и структуре, отличающей данную последовательность от других, запускаемых в иных случаях), соответствует «свой» специфический уровень мотивации или «своё» эмоциональное состояние или «своё» изменение концентрации некого нейромедиатора, вроде окситоцина, серотонина, кортизола и т.п. Как многие «очевидные» выводы, этот просто неверен.
Рассмотрение всей совокупности случаев толкает нас к обобщению противоположного рода – о взаимном фундаментальном несоответствии между «структурой» и «наполнением». Или, иначе, о несоответствии между моделью поведения, запускаемой в некой специфической ситуации и мотивационным состоянием (или эмоциональной окраской), поддерживающем эту модель. То есть, всякая сколько-нибудь хорошо дифференцированная структура поведения «может быть запущена» в широком диапазоне мотивационных состояний, вплоть до не отличающихся от «обычного» состояния отсутствия контактов. И наоборот, всякое специфическое внутреннее состояние (прирост агрессивности, сексуального возбуждения, страха и т.п. специфических побуждений по сравнению с «точкой 0» покоя при готовности действовать) может использоваться для «запуска» самых разных форм поведения, не только согласующихся со спецификой состояния, но и противоречащих ей.
И это наглядно видно при сравнении однородных структур поведения у разных видов или в разных обстоятельствах контекста общения. Скажем, у краснокрылых трупиалов Agelaius phoeniceus эффективность охраны территории резидентами пропорциональна уровню тестостерона у них в плазме крови или даже превышению индивидуальной концентрации над средним уровнем. Однако, в том же поселении или около всегда присутствуют нетерриториальные особи, взрослые и молодые, которые при наличие резидентов пытаются, но не могут занять территории на стыке участков (хотя вполне могут «поделить» пустующие территории, если мы резидентов отловим и выдержим в неволе день или более).
Так вот, при имплантации тестостерон-пропионата нетерриториальным особям их агрессивность резко повышалась, они чаще завязывали драки и чаще выигрывали силовое противоборство с резидентами. Однако, несмотря на рост агрессивности, занять территорию они были по-прежнему неспособны, так как выигрыш конфликта определяется не столько уровнем агрессивности, сколько точностью её «укладки» в специфические формы территориальных демонстраций и структуры обмена территориальными демонстрациями во взаимодействиях (чтобы соответствовали видоспецифическим «правилам игры»).

Краснокрылый трупиал
Здесь же инъекции тестостерона нетерриториальным особям а) больше увеличивают неритуализованное выражение агрессии чем ритуализованное и б) даже уменьшают точность соответствия индивидуального выражения территориальной агрессии тем «формам» видовых демонстраций, обмен которыми в определённым образом организованных последовательностях только и ведёт к выигрышу (см. пример про «дуэли кивания» тетеревов).
А вот инъекции тестостерон-пропионата резидентным особям увеличивают их способность выигрывать конфликты, поскольку 1) усиливают ритуализованную агрессию больше ритуализованной, и 2) в целом увеличивают точность соответствия индивидуальной агрессии специфическим формам видовых демонстраций и специфике их организации в последовательности, ведущие к выигрышу одной и проигрышу другой птицы, без чего результат территориальных конфликтов не будет ни устойчивым, ни «общезначимым» (также как в помянутых «дуэлях» тетеревов).
Аналогичное несоответствие между эффективностью агрессивного поведения и параметрами гормонального состояния, вроде бы влияющими на агрессивность, было также обнаружено в стаях буроголовых гаичек, где внутривидовая агрессия реализуется посредством внутригрупповой иерархии (отношений доминирования-подчинения в группе), а не территориальности. См. аналогичные данные по песчанкам, как введение антидепрессанта парната, снижающего уровень агрессивности, и в первую очередь у доминантов, не привело к изменения социальной структуре группировки, которая держится, вроде бы, на агонистическом доминировании одних особей над другими.

Буроголовая гаичка
Другой пример несоответствия «структуры» и «наполнения» состоит в том, что одна и та же форма социального поведения у вида реализуется в разные сезоны, причём в одни сезоны её реализация связана с внутренним переносчиком, определяющим состояние, а в другие – нет. Скажем, многоголосый пересмешник (Mimus polyglottоs) наиболее жёстко охраняет территории осенью, когда новые особи пытаются внедриться в существующие группировки. Агрессивная мотивация неполовозрелых самцов существенно выше, они больше поют и чаще инициируют территориальные конфликты, чем половозрелые самцы. Тот же уровень различий зафиксирован между новыми вселенцами и местными птицами – первые больше поют и больше заняты пресечением нарушений только что установившихся границ. В обоих случаях более агрессивные и территориально активные птицы по содержанию тестостерона, дигидротестостерона и лютеиннизирующего гормона в плазме крове не отличались от птиц противоположной категории.
Осенний всплеск территориальной агрессии M.polyglottos происходит на фоне неуклонного снижения концентрации половых стероидов в крови, в соответствии с обычным для птиц годовым циклом изменений гормонального статуса (управляемым фотопериодически). Однотипный подъём территориальной агрессии, тот же тип территориальных отношений между резидентами и новыми вселенцами происходит у M.polyglottus весной, но на фоне подъёма концентрации половых стероидов (Logan, Wingfield, 1990). См. аналогичное явление у певчего воробья Melospiza melodia.

Таким образом, есть основания утверждать диалектическую противоположность «структуры» и «наполнения» в социальном поведении животных и особенно в его самой специфической части — последовательностях ритуализированных демонстраций (вместо «согласия» того и другого, предполагаемого той частью этологов 50-х-70-х годов, которые следовали за Н.Тинбергеном — но не К.Лоренцом).
Аналогичным образом, касаясь генетики поведения, можно сказать, что наследуются признаки, относящиеся к «наполнению», но не к «структуре» соответствующего поведения. «Структуры» обычно возникают путём самосборки из субъединиц, которые есть в наличии. Поэтому «поведение управляет генами», а вот гены поведением – нет. Они лишь участвуют в создании «расходных материалов» для сборки нужных структур в нужном месте. Но это отдельная тема.
Гормоны и поведение
Хороший пример актуальности противопоставления «структура vs наполнение» — споры о функциях крайне популярного сейчас гормона окситоцина. Сперва их интерпретировали исключительно как поддерживающие взаимную любовь, доверие и (и вместе с дофамином) создающие социальную привязанность, но затем появились данные о том, что в ряде ситуаций он, наоборот, усиливает агрессию и чувство враждебности. Или у имеющих некоторое психическое отклонение он увеличивал недоверие и подозрительность. У тех, кто связывал действие окситоцина со спецификой внутреннего состояния деятелей (любовь vs агрессия) это вызвало понятное замешательство, но усилило позиции тех, кто связывает эффект окситоцина с «социальным творчеством» — возможностью генерировать новые модели поведения в изменившейся ситуации, не нарушая устойчивости социальной связи, существовавшей между данными индивидами доселе.
«…если вернуться к теме стирания навыков и знаний, то в этом случае окситоцин действует особым образом. У овец выработка окситоцина происходит в обонятельной луковице — той части мозга, которая участвует в восприятии запахов – при появлении каждого нового помета ягнят. Овцы и многие другие животные устанавливают связь со своим потомством, или «запечатлевают» его в памяти, с помощью запаха. Они проявляют материнскую заботу в отношении своих собственных ягнят и отвергают чужих. Но если ввести овце окситоцин, одновременно показывая ей незнакомого ягненка, она будет заботится о нем как о своем собственном.
Причем выработка окситоцина не происходит в момент появления первого помета, а только при рождении следующих пометов. Это позволяет предположить, что окситоцин выполняет функцию стирания связей матери с ее первым пометом, создавая условия для возникновения связи со вторым пометом. (Фриман допускает, что связь матери с первым пометом возникает под действием других нейрохимических веществ.). Способность окситоцина уничтожать предыдущую модель поведения натолкнула ученых на мысль назвать его «гормоном забывания». Фриман предполагает, что окситоцин уничтожает существующие нейронные связи, чтобы создать условия для формирования новых привязанностей. Согласно этой теории окситоцин не учит родителей заботиться о своем потомстве. Он также не делает влюбленных более сотрудничающими и добрыми; он просто способствует освоению новых моделей поведения.
…С помощью таких нейромодуляторов, как окситоцин, природа позволяет двум влюбленным пережить период повышенной пластичности, приспособиться друг к другу и сформировать новые намерения и ощущения. По мнению Фримана, мозг представляет собой, главный образом, орган социализации, поэтому в нем должен существовать механизм, который время от времени ослабляет присущую нам склонность быть излишне индивидуализированными, слишком эгоцентричными и занятыми самими собой.
Фриман пишет: «Важнейшее значение сексуального опыта заключается не в получении удовольствия или даже не в размножении, а в возможности преодолеть пропасть индивидуализма… В процессе формирования доверия важнее то, что следует за любовной игрой, а не предшествует ей» / Норманн Дойдж. Пластичность мозга
Тем более, что окситоцин усиливает «социальное притяжение» и «закрепляет» результаты социальных контактов в ситуации, когда социальные связи и модели поведения были прежними. Когда он делает то же самое в ситуации изменённых моделей поведения, внесения социальными компаньонами новшеств, он так же «работает» на устойчивость социальных связей, только динамическую в дополнение к статической. Поэтому он чаще всего усиливает ту тенденцию во взаимодействии с другими людьми, которая свойственна индивиду- доверчивых делает ещё доверчивее,подозрительных и враждебных – ещё враждебнее, но при этом компенсирует недостаток социабельности у тех людей, у которых она низка в силу разных причин.
Кроме окситоцина в роли «генератора» новых моделей поведения выступает кортизол, опосредующий реакции стресса. С точки зрения этолога его эффект оказывается «в том же роде», что у окситоцина (поддержание новых моделей поведения без ущерба для устойчивости прежних связей), но противоположным по знаку.
Окситоцин поддерживает новые модели поведения «положительного» характера, связанные с новыми факторами «социального притяжения», обычно с позитивной эмоциональной окраской. Кортизол же поддерживает новые модели поведения, вызванные новыми факторами тревоги и стресса, т.е. окрашенные отрицательно. Так сказать, изменения концентрации окситоцина опосредуют модели поведения, вызванные желанием «потянуться к новой морковке» (где морковка – перспективный социальный партнёр и взаимодействия с ним), изменения концентрации кортизола – побуждение «отшатнуться от нового кнута», и при этом с минимальной угрозой устойчивости прежних связей. Например, «для красной полёвки Gletrionomys rutilus показано, что «в годы высокой численности (пик и спад) усиливается нейросекреция вазопрессина в портальный кровоток гипофиза, что может приводить к активации гипоталамо-гипофизарно-надпочечниковой системы, а в годы низкой численности возрастает синтез окситоцина, играющего ключевую роль в стимуляции генеративной функции (Владимирова и др., 2006)» (op.cit.).

Красная полёвка
Связь стрессированности с социальным статусом
Говоря о влиянии стресса и посредников в передаче стресс-воздействия, кортиколиберина и кортизола, на поведение, нужно сказать два слова о контринтуитивной связи степени стрессированности животного с социальным статусом. Априори нам кажется, что в сообществах, особенно построенных по принципу иерархии, наиболее стрессированными должны быть низкоранговые особи, а доминанты – практически нет (они же свободны проявлять своё поведение в присутствии других, не считаясь с их реакциями!).
Реальность куда сложнее и интереснее. Я уже писал, что близкие виды делятся на «социальные» и «несоциальные», причём те и другие могут формировать поселения с высокой плотностью, правда, по разным причинам. [Первые — под действием первичных сигналов, связанных с концентрацией корма, убежищ и т.п. ресурсов; вторые — под действием вторичных сигналов, связанных с социальными взаимодействиями конспецифических особей соответственно]. Так вот, когда высокая плотность поселений наблюдается у несоциальных видов, то там – да, более стрессированы подчинённые особи, и стресс сильно влияет на их поведение, особенно на репродуктивную функцию. А вот у социальных видов, скорей, наоборот – при столь же высокой плотности поселений (которую они, в отличие от близкородственных несоциальных видов, стараются поддерживать хотя бы на части пространства даже в условиях резкого падения численности) наиболее стрессированы доминанты, а не подчинённые.
Реакции стресса здесь определяют здесь дифференциацию поведенческих ролей (т.е. создают новые модели поведения для исходно «равноправных» особей), задают соответствие поведение роли вместо «общего подавления» у несоциальных видов.
«…интересны результаты выполненного Крилом (Creel, 2001) сравнения опубликованных данных, характеризующих базальные уровни глюкокортикоидов у доминирующих и субдоминантных особей среди кооперативно размножающихся и некооперативных видов пресмыкающихся, птиц и млекопитающих, исследованных в природе или в условиях, приближенных к естественным. Проведённый им сравнительный анализ 25 публикаций, включая собственные, показал, что у «некооперативно размножающихся» видов – сборной группы, включающей широкий спектр социальных систем, более высокая стрессированность чаще встречается у собординантных особей (Creel, 2001). Важно отметить, что случаи подавления репродуктивной функции стрессом описаны в основном у тех млекопитающих, для которых репродуктивная кооперация не характерна.
Напротив, у кооперативно размножающихся видов с наиболее устойчивыми системами доминирования-подчинения повышенный уровень глюкокортикоидов в большей степени свойствен доминантам, а не подчинённым, как это принято было считать, исходя из результатов в основном лабораторных исследований. В экспериментах по объединению животных в клетках и вольерах, исключавших возможность бегства, само ограниченное пространство выступало в качестве фактора стресса у подвергшихся агрессии индивидов. В естественной обстановке субординантные особи у кооперативно размножающихся видов способны предвидеть развитие социальных конфликтов и уходить от стрессовых ситуаций. Повышенный уровень стрессированности у доминантов в природе отмечен у самцов альпийского сурка (Marmota marmota; из 4 типов субординантов у одного всё же уровень глюкокортикоидов был выше; Arnold, Dittami, 1997), у самок лемура катта (Lemur catta, Cavigelli, 1999), у самок флоридской кустарниковой сойки (Aphelocoma coerulescens, Schoech et al, 1991), у самок карликовых мангустов (Helogale parvula; Creel et al, 1992), самцов и самок гиеновых собак (Lycaon pictus; Creel et al, 1997). К этому нужно добавить недавнее исследование дикой популяции волка (Canis lupus; Sands, Creel, 2004), в котором повышенным уровнем глюкокортикостероидов обладали как высокоранговые самцы, так и высокоранговые самки. При этом у волков высокий уровень глюкокортикоидов у доминировавших особей, в отличие от карликовых мангустов и гипновых собак не был связан с повышенным уровнем агрессивности. Иерархия отношений у волков поддерживалась исключительно на основе ритуализованных форм поведения.
Повышенная стрессированность доминантов среди кооперативно размножающихся особей интерпретируется Крилом в терминах «цены доминирования» [единственным исключением из этого правила является знаменитый голый землекоп, где альфа-самка, где фиксируется отрицательная корреляция концентрации глюкортикоидов с социальным статусом особи. Однако здесь взаимодействия с доминанткой приводят к развитию у подчинённых самок стресс-синдрома, полностью и необратимо выключающей их из размножения, в отличие от всех вышеназванных видов. Так что исключение подтверждает правило.]. Поддержание высокого социального ранга энергетически дорого, однако повышенная стрессированность доминантов не отражается на их репродуктивной функции. Исключение же из размножения субординантных особей осуществляется благодаря поддерживаемой доминантом иерархии социальных рангов, то есть в основном за счёт поведенческих ограничений контактов с самками»/ источник.
Стресс как модификатор поведенческих программ
Теперь, собственно, про роль кортизола как создателя новых моделей поведения, индуцированных стрессом. В обзоре Е.А.Новикова и М.П.Мошкина «Роль стресса в модификации поведенческих программ» (2009) приведены данные о том, что физиологические последствия длительной активации адренокортикальной функции в ответ на разного рода набор средовых и социальных стрессоров оказывают модифицирующее воздействие на поведение, в диапазоне от новых ситуативных моделей последнего до новых вариантов организации жизненного цикла в зависимости от силы и долгосрочности эффекта. Главнейшие изменения состоят в устойчивой модификации эндокринной системы, задержке полового созревания, усиления миграционной активности и ослаблении социальной конкурентоспособности, в т.ч. по причине направленного отказа животного конкурировать.
Эти эффекты, традиционно рассматриваемые как негативные, в целом ряде ситуаций могут быть полезными и спасительными в силу сдвига трейд-оффа между размножением и самосохранением в пользу второго в случае «упадка сил», или экстремальности внешней среды (подробнее здесь, п.6). Например, «… стресс, тормозящий развитие и репродуктивную функцию в неблагоприятных условиях, может предотвращать и иммуносупрессию, которую в размножающихся животных вызывают половые гормоны – тестостерон у самцов (Grosmann, 1985, Folstag, Karter. 1992) и прогестерон у беременных самок (Szekeres-Bartho, 1992).Сохранение высокой иммунореактивности у неполовозрелых сеголеток уменьшает вероятность их гибели в зимние месяцы, несмотря на высокую адренокортикальную активность (Bujalska et al, 1994; Moshkin at all, 1998)».
Стресс и опосредующие его гормональные изменения формируют новые модели поведения несколькими путями. Во-первых, это модифицирующее влияние на поведение пренатального стресса. «…Стрессирующие воздействия, вызванные переуплотнением, недостатком корма, хищниками и т.д., которые испытывают беременные самки, сказываются затем на темпе роста, созревания и на поведение детёнышей, оказывают влияние на способности к расселению и репродукцию. Известно, что действие стрессоров в пренатальном и раннем постнатальном периоде развития вызывает снижение стресс-реактивности потомков во взрослом возрасте. В опытах на крысах Н.Н.Дыгайло и Е.В.Науменко (1983) показали, что адренокортикальная реакция потомков на эмоциональный стресс находится в обратной зависимости от уровня глюкокортикоидов в материнском организме в последнем триместре беременности.
Эти нарушения сходны с наблюдаемыми у людей, подверженных эндогенным депрессиям, и связываются с дефицитом рецепторов глюкокортикоидов в гипоталамусе и других отделах мозга…». Кроме того, наблюдается демаскулинизация самцов и снижение фертильности самок. У рыжей полёвки пренатальные стрессы во второй половине беременности приводят к замедлению полового созревания, ослабления социальной конкурентоспосбности самок и повышению агрессивности самцов. Кроме того, запах пренатально стрессированных особей менее привлекателен для их половых партнёров, чем запах интактных животных.
Кроме того, во всех случаях стресс-индуцированное перераспределение ресурсов материнского организма включает ухудшение кровоснабжения брюшной полости, что ведёт, в т.ч. к гипоксии плодов с ухудшением их трофического обеспечения.
…Даже у яйцекладущих животных гормональный статус родителей может модифицировать ход онтогенеза потомков за счёт глюкокортикоидов, попадающих в яйца из материнского организма. Так, искусственное увеличение содержания кортикостерона в птичьих яйцах приводит к снижению в скорости роста птенцов, изменениям в их поведении, подавлению клеточного иммунитета и модификации адренокортикальной реактивности на острые стрессирующие воздействуия, которая увеличивается у птенцов и снижается у взрослых птиц […]. Степень проявления и направленность этих эффектов могут зависеть от пола особи. Инъекция кортикостерона в яйца японского перепела у самцов вызывала снижение скорости роста, у самок – снижение стресс-реактивности во взрослом состоянии […]. Введение кортикостерона «беременным» живородящим ящерицам вызывало уменьшение линейных размеров, массы тела и упитанности у ювенильныхъ самок (что отрицательно влияло на их жизнеспособность), но повышало выживаемость самцов […]». См.также про «сигнальную наследственность» у птиц.
Во-вторых, это модифицирующее влияние эмоционального стресса — если он не выходит за некоторые пределы, то мобилизует силы индивида и трансформирует поведение так, что может реализовываться и в стрессовой ситуации или позволяет «переждать» её, как бы пропустив ход (особенно это касается «пропуска» размножения, исключительно рискованного для особи в т.ч. вследствие ослабления иммунной защиты пропорционально инвестициям в токование).
«Исходно предполагалось, что имеет особо негативные последствия для организма имеет т.н. эмоциональный стресс, в основе которого лежит фрустрация какой-либо мотивации – невозможность спастись бегством при опасности, получить доступ к репродуктивному ресурсу, не рискуя быть выдранным доминирующими особями и т.п. Причём надо сказать, что противопоставление «физических» стрессов «эмоциональным» возникло в исследованиях патофизиологов, подвергавших подобпытных животных жёстким и фиксированным экспериментальным воздействиям. В природе же дела обстоят совершенно иначе – все воздействия и даже внешние (пища, вода, хищники, укрытия и т.п.) воспринимаются особью неспецифически, как некое нарушение, проблема, затрудняющая устойчивое взаимодействие с социальными компаньонами в типичном для вида режиме. Поэтому в природе «все стрессы эмоциональны»…
…при таких традиционно считавшихся «физическими» стрессах, как острое охлаждение и голодание, животное не может реализовать мотивацию к достижению температурного комфорта и к удовлетворению голода. У лабораторных и домашних животных предъявление хищника или его запаха вызывает лишь краткосрочные изменения гипоталамо-гипофизарно-надпочечниковой системы […], однако в природе угроза нападения хищника может приводить к развитию эффектов, проявляющихся при продолжительных стрессах […]. Концентрация кортикостерона в крови у молодых самцов больших песчанок положительно зависит от частоты посещения их колонии серым вараном […]. У самок каменок при обитании на их гнездовых участках сорокопутов повышается уровень кортикостерона в крови, снижается вероятность повторной кладки и задерживается её начало […].
Исследования, проведённые на американских зайцах-беляках и певчем воробье Melospiza melodia показали, что совместное влияние хищничества и дефицита пищи на демографические параметры популяций оказывается более значимым, чем влияние каждого фактора по отдельности […]. Гипотеза, сформулированная на основе этих данных Бунстрой и Кребсом […] предполагает, что перманентным источником стресса в данном случае является эмоциональное напряжение, связанное с необходимостью поиска пищи в условиях риска хищничества» (Новиков & Мошкин, op.cit.).
Всё это в конечном счёте включает онтогенетическую программу отсроченного размножения (сейчас пока непонятно, в какой степени она обратима, и у каких видов), что ведёт к поливариантности онтогенеза разных особей внутри популяции. Скажем, «у сеголеток красной полёвки, изъятых из популяции на пике численности последней и одиночно содержащихся в лаборатории, масса семенников была выше, а содержание глюкокортикоидов в крови – ниже, чем у зверьков, содержащихся в группах по 4, или чем у продолжающих онтогенетическое развитие в природе…». Это позволяет особям делать выбор между немедленным и отсроченным размножением сообразно экологической и социальной ситуации, соответственно манипулируя реализуемой моделью поведения и/или жизненного цикла особи.
«У бореальных мышевидных грызунов сеголетки первых генераций успевают созреть и принять участие в размножении уже на 1-м году жизни в возрасте 1,5-2 мес., тогда как особи поздних генераций остаются неполовозрелыми до следующей весны и приступают к размножению в возрасте 8-10 мес. […]. У белок репродуктивные показатели в текущий сезон размножения коррелируют не столько с предшествующим, сколько с будущим урожаем орехов […]…У большой песчанки весной концентрация глюкокортикоидов в крови не коррелирует с локальной плотностью, но находится в обратной зависимости от будущего обилия кормовых растений во время летней бескормицы […]» (Новиков & Мошкин, op.cit.) и т.д.
Соответственно, на смену представлениям о «правильном» ходе онтогенеза и разных отклонениях от него, приходит представление о поливариантности онтогенетических программ (или жизненных циклов), гибко адаптирующих особей разного «качества» (различающихся, скажем, объёмом ресурсов и/или конкурентной мощностью) к среде большей или меньшей степени экстремальности, так что в устойчивой ситуации все варианты в среднем имеют близкую приспособленность. И это поведенческое многообразие воспроизводится в популяции, работая на её устойчивость.
Надо сказать, что слишком сильное стресс-воздействие (или нетипичное для данного вида) явно нарушает поведение, вместо того, чтобы приспособительно модифицировать его. «Экспериментальные данные, полученные на грызунах и обезьянах [т.е. видах с хорошо защищёнными гнёздами и развитой заботой о потомстве.] свидетельствует о том, что испытавшие пренатальный стресс особи во взрослом состоянии оказываются неспособными адекватно реагировать в ситуациях опасности, более пугливы в случаях новизны. У них наблюдается нарушение функционирования гипоталамо-гипофизарно-надпочечниковой системы, выражающейся в повышенной концентрации циркулирующего АКТГ, что отчасти объясняется снижением чувствительности механизма отрицательной обратной связи по причине того, что у пренатально стрессированных животных уменьшается плотность глюкокортикоидных рецепторов в гиппокампе, а также подавляется активность стресс-ингибирующих механизмов, в которых принимают участие эндогенные опиоиды и ГАМК-эргические или бензодиазепиновые рецепторы (Weinstock, 1997). Эти нарушения сходны с наблюдаемыми у людей, подверженных эндогенным депрессиям, и связываются с дефицитом рецепторов глюкокортикоидов в гипоталамусе и других отделах мозга…» (Мошкин & Роговин, op.cit.).
То есть, стресс-воздействие в избыточной дозе означает разрушение адекватной модели поведения, а вот в физиологической дозе – её изменение сообразно обстоятельствам, чтобы, изменяя структуру жизненного цикла в пользу одного из возможных вариантов, достичь максимума репродуктивного успеха на «финише». Так у птиц и ряда млекопитающих стрессы, испытанные матерями в период беременности или формирования яиц, подготовки к яйцекладке и пр. рассматриваются как адаптирующие потомство к тем условиям среды, в которых они сами существовали во время беременности, т.е. в отличие от приведённых данных по грызунам и приматам как не вполне патологические.
Поэтому так важно, может ли эмоциональное напряжение в каждой из специфических проблемных ситуаций быть «оформлено», то есть реализоваться в виде специфических структур поведения, запуск которых разрешает соответствующую «проблему». Если да, то мобилизация ресурсов, связанная со стрессом, идёт организму только на пользу – скажем, в ответ на появление захватчика животное не просто атакует его, но вступает в организованный обмен демонстрациями, и выбирает из видового репертуара именно те, которые будут наиболее успешны применительно к данному противнику. То есть модифицирует модель поведения сообразно конкретному раздражителю и общей напряжённости социальных контактов в сообществе. Если нет, и эмоциональное напряжение остаётся лишь «наполнением», не преобразуясь в «структуру», стресс разрушителен.









{kind=link}
{kind=link}