

Большие полынные тетерева на току
В продолжение темы индивидуальности у животных
Я много писал про альтернативные стратегии поведения (или «жизни в целом»), когда одну и ту же проблему разные особи в популяции решают противоположными способами. Например, самцы снежного барана Ovis canadensis сanadensis на с.-з. Монтаны демонстрируют 3 стратегии спаривания[1] – основную («ухаживание», 1) и 2 альтернативные («курсирование», 2 и «блокирование», 3). В случае 1 самец просто следует за самкой в эструсе, не пробуя направлять её движений, а лишь охраняя от прочих самцов. Коитус частый и короткий (0.90±0.11 и 2.30±0.12), во время которого самка стоит. Эта стратегия свойственна самцам из верхней трети иерархии доминирования, и демонстрируется в пределах ограниченного участка территории стада, одного и того же в разные годы, чем аналогична таковой гаремных (территориальных) самцов благородного оленя.

Снежный баран
В случае 2 самец завязывает поединок с «ухаживающим» самцом и при успехе спаривается с охранявшейся им самкой (она при этом двигается). Коитус редкий (0.39±0.06 раз/час), и самки здесь с почти равной частотой спариваются с «охраняющими» и «курсирующими» самцами. Эта стратегия напоминает рейдеров благородных оленей, но осуществляется не «потихоньку», а «на глазах у всех». В случае 3 самец заранее, ещё до наступления эструса, отгоняет самку или группу самок к себе на участок, и там блокирует, а дальше действует по стратегии 1. Ряд самцов может менять стратегии в зависимости от социального контекста (и, таким образом, иметь несколько стратегий спаривания в репертуаре), но чёткость их выделения остаётся неизменной.
Альтернативные стратегии как источник социального полиморфизма
Альтернативы вроде вышеописанных лежат в основе внутривидового социального полиморфизма – когда наблюдается несколько форм отношения к территории, или взаимоотношения полов, или систем доминирования, переходящих друг в друга при изменении обстоятельств контекста, таких как плотность и интенсивность взаимодействий, корм, хищники и другие условия существования[2].
Красивый пример альтернативных стратегий, баланс которых сопрягается с динамикой популяций и изменяется в зависимости от стадий последней – поведение особей красно-серой полёвки, относящихся к цветовой морфе «чёрный огузок» (см.М.А.Потапов и др., 2011). С этим же признаком сопряжена более интенсивная окраска чепрака, см.рис.1-2. В период депрессии численности вида под Новосибирском такие зверьки встречались единично, но в период подъёма их доля быстро росла, достигнув 10-15% на пике. Аналогичные цветовые формы со сходной динамикой встречаемости отмечены и в других популяциях вида.


Дальше этологические эксперименты О.Ф.Потаповой с соавт. (2014) показали, что «чёрный огузок» имеет ряд преимуществ над типично окрашенными зверьками в условиях высокой плотности. Прежде всего, как положено меланистам, он «разумней дозирует» агрессию, более миролюбив в «непонятных» и новых ситуациях, и лучше отличает их от требующих агрессии, проявляет там больше исследовательской, чем тревожно-оборонительной реакции. Поэтому именно при высокой плотности такие самцы более привлекательны для самок, а также набирают большую массу и занимают больший ранг в социальной иерархии.
Вообще, у этого вида и у серой полёвки самыми привлекательными для самок оказываются субдоминанты 1-го порядка, с максимальным миролюбием при достаточной агрессивности. Частота же миролюбивых контактов партнёров при парном ссаживании – важный предиктор успешного размножения пары. Именно они преобладают среди «чёрных огузков», среди «типичных» — менее привлекательные субдоминанты 2-го порядка (доминанты же, с наибольшей агрессивностью и минимальным миролюбием, наименее привлекательны). См.рис.3-4.
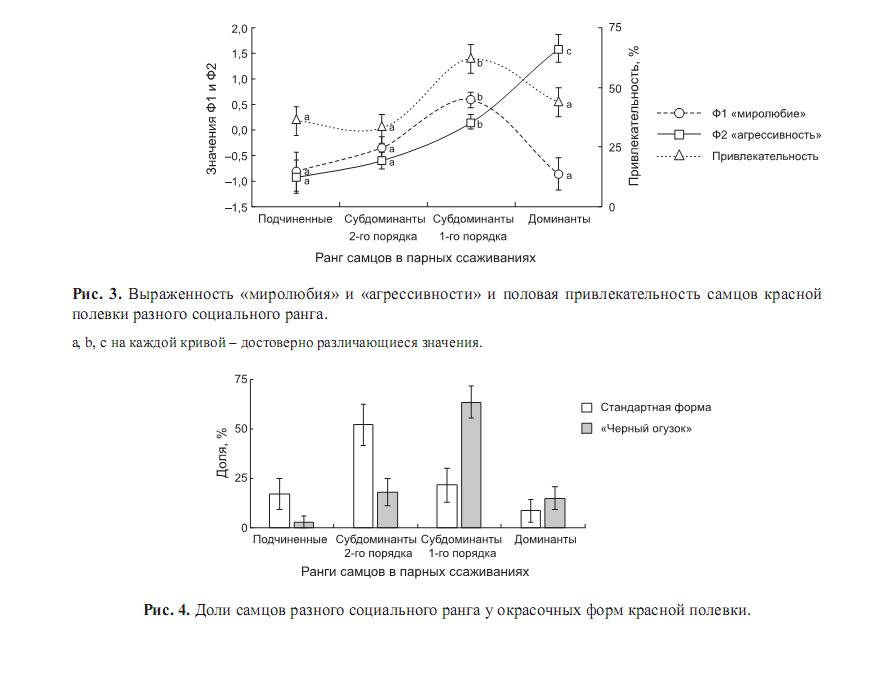
Соответственно весной они оставляют больше потомков (относительно «типичных»), тем более что и начинают размножаться раньше. А к осени их доля падает, и из-за большей элиминации, и потому что приплод «типичных» поступает в популяцию позже.
Также показано дифференцированное участие разных морф в циркуляции очага клещевого энцефалита, существующего под Новосибирском. Иммунореактивность «миролюбивых» самцов существенно выше, а их, при любом ранге, больше среди «чёрных огузков». В разреженной популяции все соответствующие преимущества теряют значение, и доля «чёрных огузков» падает.
Другой пример: самцы тетерева для привлечения самок и спаривания могут собираться на токах (и очень привязаны – много лет – к выбранному току), а могут и токовать одиночно. Или самцы дрофы Otis tarda из 9 популяций, изученных в венгерской степи, в 2-х придерживаются строгой моногамии, в 7 других в разных сочетаниях наблюдаются моногамия, полигиния и промискуитет.
В конце зимы стаи дроф распадаются и самцы занимают токовые площадки. У моногамных и полигинных самцов они охраняются, развито специфическое территориальное поведение, у промискуитетных же – нет, они токуют группами рядом. Моногамные самцы спариваются лишь с одной самкой из привлечённых к месту тока, а потом постоянно держатся в первых десятках метров от гнезда. Участки полигинных самцов крупнее, охраняются лучше, и привлекают группы самок – до 4-х, между которыми устанавливается иерархия. После спаривания поведение полигинных самцов неустойчиво – он может держаться у гнезда одной из самок, как моногамный, или плюнуть на родительские обязанности и примкнуть к группам неразмножающихся самцов, как делают промискуитетные после всех спариваний. При групповом токе контакт полов ограничивается лишь краткой копуляцией. См. статьи I.Sterbetz (1980: c.32-141) и Carranza J. et al. (1989).

а — токующий дрофич, б — спокойный

а

а

б
Третий пример – западноафриканские бюльбюли Andropadus latirostris. Половой диморфизм у них отсутствует, даже около экватора сохраняется сезонность размножения: яйца и гнёзда с птенцами находят только в период с 15.12 по 10.4. Вид крайне подвижен, нетеррриториален; ведя номадный образ жизни, формирует популяции с очень высокой плотностью населения.
Самцы не принимают участия в заботе о потомстве и (в месте исследований A.Brosset) формируют тока, местоположение которых было постоянным все 10 лет наблюдений (1970-1980 гг.). Самцы на токах нетерриториальны: дистанция между ними варьирует от 10 до 20 м, сам ток занимает ≤0/5 га. Поющие самцы на току могут размещаться линейно (наибольшая плотность здесь 20 одновременно поющих / 40 м), или «по вертикали», занимая всю толщу леса от невысокого кустарника до вершин крон на высоте ≈50 м. Среднее расстояние между токами 600-1200 м.; число самцов на них постоянно меняется, и может достигать нескольких десятков одновременно.

западноафриканские бюльбюли Andropadus latirostris
Вместе с тем постоянно встречаются одиночные самцы, поющие вне токов. Они не приурочены к постоянным территориям и остаются номадными даже в период ухаживания (тем более что самцы A.latirostris никак не участвуют в заботе о потомстве). Они так же спариваются, как поющие на току, и не «хуже» их в этом плане. В других популяциях вида, живущих с существенно меньшей плотностью, ни токов, ни номадных особей нет, моногамия, территориальность и дисперсное гнездование (Brosset, 1982).
Происхождение альтернативных стратегий: переходный полиморфизм или устойчивый баланс?
Каждый из противоположных способов решения проблемы не остаётся частностью, имеющей отношение к ней, и только. Он так или иначе влияет на побочные и/или смежные аспекты биологии вида, почему каждая из альтернатив может рассматриваться широко, как модель или стратегия поведения, реализуемая разными особями в популяции, на разной основе (в смысле различий в «генетике»-«биологии» индивидов), с разной точностью и успехом. Каждая особь, принявшая одну из альтернатив и реализующая её более-менее успешно и точно – статистическое испытание модели (или стратегии), общую сумму которых оценивает отбор, который одновременно и совершенствует обе модели, и увеличивает точность подбора» им реализаторов.
Скажем, альтернативные стратегии брачного поведения тетеревов (собираться на ток vs токовать одиночно) объяснимы двумя разными способами. Первый — токующие одиночно это «худшие» особи, неудачники, не умеющие удержать территорию на току, и воспользоваться выгодами постепенного «продвижения» самца вместе с участком от периферии к центру по мере его старения и выбытия центральных птиц. Второй вариант — это альтернативная стратегия, равнозначная токованию на току, в том смысле, что даёт своим приверженцам равный уровень приспособленности, как минимум пока пространственно-этологическая структура группировки устойчива.
Как проверить, какая из версий лучше соответствует фактам? Проанализировать многолетние данные о соотношении тех и других в устойчиво растущей популяции. Если первая т.з. верна, увеличение среднего размера токов, ожесточение конкуренции за территории больший % особей делают «худшими», исключёнными, заставляя токовать одиночно. Соответственно, при верности первой т.з. в растущей популяции % токующих одиночно самцов будет тоже расти, при верности второй — не изменится, будет сохраняться определённое равновесие между стратегиями. Что было сделано в работе D. E. Chamberlain et al. (2012).
Если первая точка зрения верна, будут выполняться ещё 3 предсказания: 1) при росте численности популяции существует некий порог, после которого число особей, собирающихся на токах, больше не возрастает, растёт только число одиночек; 2) местообитания, к которым приурочены тока — лучшие, и поэтому занимаются тетеревами раньше, чем районы одиночного токования; 3) одиночно токующие тетерева в целом приурочены к худшим местообитаниям, и их численность там будет возрастать при росте численности всей популяции.
Однако анализируемые авторами данные по 25-летней динамике растущей популяции тетерева на западе итальянских Альп опровергают все эти предсказания. С ростом численности популяции пропорции спаривающихся на току и токующих одиночно остаются без изменений; число токов и средняя численность птиц на току плавно растут вслед за ростом численности популяции, порога не выявляется; не никаких данных о лимитированности биотопов, в которых располагаются тока. Однако подтверждается предположение, что сперва в каждом районе сперва растёт численность той части популяции, что собирается на токах, а затем — токующей одиночно (хотя и не в худших биотопах). Иными словами, одиночное токование для тетерева — это не «подчинённая» или «худшая», а просто альтернативная стратегия.
Исследование социального поведения в сообществах позвоночных, особенно птиц и млекопитающих (и построенных на системе агрессивного доминирования, и организованных по системе территориальности, когда ранг связан со способностью перераспределять общее пространство группировки в свою пользу) потребовало скорректировать эту точку зрения. Если отслеживать все сближения с самками и копуляции, не только «официальные» (выполненные в рамках типичной схемы социальных отношений группировки, но и «теневые») подчинённые особи оказываются не менее успешны, чем доминанты.
По крайней мере, пока социальная организация устойчива, есть избыток ресурсов для реализации соответствующего паттерна отношений и т.д. На самом деле поведение подчинённых особей образует специфическую стратегию, альтернативную по отношении к стратегии доминантов, и не менее успешную чем она. Вот кто всегда проигрывает после выделения в сообществе специфических стратегий — это особи среднего статуса, реализующие смешанную стратегию и запаздывающие с дифференциацией/специализацией социального поведения как доминантного/подчинённого. Они если и выигрывают, то только в периоды неустойчивости социальной организации или на периферии ареала, где оная устойчивость ещё не создана.
Итак, стратегии доминантов и подчинённых в норме сопоставимы по «гарантируемой» величине итоговой приспособленности, но прямо противоположны по образу реализуемого поведения. Доминант выигрывает за счёт прямого и демонстративного перераспределения социальных ресурсов (лучших территорий, «верхних» социальных статусов и т.п.) в свою пользу и таким образом монополизирует репродуктивные ресурсы — самок, которые таким же образом оценивают «качество» территории, социального статуса самца и т.п. Однако большую часть времени доминантные самцы тратят во взаимодействия друг с другом, «монополизированные» ими самки испытывают сексуальную депривацию… И тут возникает возможность для проникновения подчиненных самцов — рейдеров и успешного спаривания с «обделёнными» самками (тем более что в ответ на агрессию доминанта они не отвечают агрессией, а просто терпят, не отказываясь, однако от своих планов).
Обратите внимание, что в такой системе доминантные самцы несут существенную «альтруистическую нагрузку» по формированию той пространственно-этологической структуры группировок, «дырами» в которой пользуются подчинённые особи. И если обстоятельства вдруг складываются так, что доминанты избирательно попадают под выстрел, как самцы-рогачи благородного оленя, или элиминируются по другой какой-то причине, одни только рейдеры не могут восстановить деградирующую социальную структуру группировки — они должны «выдвинуть» из своей среды особей, реализующих стратегию доминанта, но нечувствительных к фактору элиминации[3].
Другой важный вопрос, к которому мы вернёмся потом – почему соответствующие стратегии всегда выступают именно как альтернативы друг другу (особенно с т.з. особей – реализаторов, как бы выбирающих, в какую «вложиться» своей активностью, жизнью, демографическим потенциалом и пр.)? Почему они «не умеют» решать эту проблему «просто по-разному»?
Классификация типов альтернативности: основные трейд-оффы в популяционной системе
Как известно, сложную систему нельзя оптимизировать одновременно более чем по одному параметру. Поэтому «альтернативность» стратегий (моделей поведения), «имеющих хождение» в данной популяции, основана на трейд-оффах, отражающих невозможность для организма направить ресурсы сразу в несколько конкурирующих активностей (форм поведения). Поэтому приходится выбирать, всякий сделанный выбор канализирует возможности изменения ещё больше и пр.
Поэтому интересно описать более-менее полный набор основных видов трейд-оффов у позвоночных, производных от них способов решения проблемы, «оформляющихся» в биологии вида как альтернативные стратегии.
Всего их (разных) удалось насчитать 5:
А. Между конкурентной и патиентной стратегиями поведения относительно стрессоров из «двух сред» — социальной и экологической.
Они различаются по распределению усилий и ресурсов животного между приспособлением к социальной и экологической среде. Первая отличается большей приспособленностью к социальному стрессу, большей конкурентоспособностью в социальной жизни, вторая — большей приспособленностью к средовому стрессу. Первое характеризуется максимумом устойчивости реализации форм социального поведения, стереотипностью и «чёткостью» исполнения, за счёт больших энергозатрат, большего BMR, как у «чёрно-белых самцов» мухоловки пеструшки. Второе, наоборот — максимумом изменчивости и пластичности поведения при минимальных энергозатратах и сниженной «мощности деятельности.
Пример. Чёрно-белые самцы vs куропёрые у мухоловок-пеструшек; «Летуны» vs «собиратели» vs «помоечники» у сизарей, что коррелирует с разной долей черночеканных особей и окрашенных по «дикому типу»; девятииглые колюшки из прудовой (низкое хищничество и высокая конкуренция) vs морской прибрежной популяций (высокое хищничество и низкая конкуренция) и т.д.
Повышенная плотность нитчатых водорослей, обусловленная эвтрофикацией, реверсирует опосредованный паразитизмом отбор во время размножения у трехиглой колюшки (Gasterosteus aculeatus). В Балтийском море большинство самцов колюшки делает гнезда в плотных зарослях растительности, однако такие самцы испытывали высокий пресс паразитизма. Высокая вероятность размножения таких самцов в плотных зарослях водорослей может быть обусловлена ухудшением видимости, ослабляющей конкуренцию между самцами или снижением разборчивости самок. Наоборот, в условиях редкой растительности отбор, обусловленный хищничеством, действует против самцов с высоким прессом паразитизма.
Б. Между коммуникативным и некоммуникативным способами конкуренции за социальный ресурс. В первом случае действия особей друг в отношении друга опосредованы обменом демонстрациями, по результатам которого определяются победитель и побеждённый (лучше и хуже использующий специфическую сигнальную информацию для закрепления и улучшения собственного социального статуса). После чего они «делят» между собой совокупные репродуктивные ресурсы популяции, распределяя доступ к самкам для спаривания либо во времени, либо между альтернативными социальными ситуациями, либо как-то ещё, так что на круг выходит примерное равенство.
Второй способ – некоммуникативный – предполагает прямое воздействие животных друг на друга в конкурентной борьбе, не опосредованное видовыми демонстрациями, обмен которыми делит особей на выигравших и проигравших автоматически. В лучшем случае оно аранжировано индивидуальными экспрессиями животного, отражающими индивидуальную же динамику состояния по ходу конкурентной борьбы (чем, естественно, могут пользоваться его партнёры и противники, если достаточно интеллектуально развиты, как сигналами ad hoc).
Такое бывает у немногих видов животных, где видоспецифический набор демонстраций ещё не развит, а взаимодействие без коммуникации – просто смертоубийство, и у тех высокоорганизованных позвоночных (обезьян Старого Света), где система видовых демонстраций уже подвергается постепенной редукции. Здесь индивиды уже «достаточно умны», чтобы понимать содержание поведения друг друга и прогнозировать последствия чужой активности в своих интересах и без поддержки видовыми сигналами инстинктивной природы, которая к тому же сковывает возможность передачи в сообщениях новой информации, накопленного опыта животного и прочего «индивидуального знания».
И вот при некоммуникативном способе социальной конкуренции приёмы борьбы оказываются прямо противоположными у доминантов и подчинённых (среднеранговые особи используют то ту, то другую тактику в зависимости от ситуации). Доминант преимущественно кроет самок «силой», пользуясь преимуществом своего статуса и используя систему быстрого спаривания. Среднеранговые особи (отчасти и доминанты) создают долговременные взаимодействия с самками через систему «дружбы» и «полового супружества», что препятствует посягательствам доминантов. А вот подчинённые особи выигрывают за счёт конкуренции спермы (sperm competition).
На уровне поведения это отражается в разном распределении усилий животного между репродукцией и удовлетворением витальных потребностей, включая самосохранение. Первое означает большие вложения в социальные взаимодействия, на выходе которых оказывается репродукция, что связано с готовностью идти на риск, второе — ограничение репродукции или отказ от неё, «экономия» на социальном взаимодействии, коммуникации, с минимизацией риска. На уровне поведения первые максимально «инвестируют» в коммуникативную сеть группировки, стереотипное предъявление должных демонстраций в должные моменты взаимодействий, поддерживающие информационный обмен в поселении в целом, что выгодно всем, но наиболее рискованно и затратно именно для активных демонстраторов. Что развивает у «лучших» смелость и связанные с ней качества.
Вторые реализуют те же формы активности наиболее «экономным способом», с максимумом использования «общей» коммуникативной сети через «подглядывание» или «обман» прочих участников коммуникации, при минимуме вложений в неё. Что даёт кратковременный выигрыш, но при ухудшении ситуации эти особи первыми исключаются из сообщества, делаются нерезидентными и утрачивают шансы на размножение в данное время и в данном месте.
Второе становится долговременно выгодным лишь, когда риск гибели от хищников и паразитов, ориентирующихся именно на сигналы, вырастает до неприемлемых величин, как у сверчков Teleogryllus oceanicus. Не стоит забывать, что у животных продуцирование сигналов, в отличие от человеческой речи — вещь весьма рискованная. Соответствующий риск сравним с риском радиста разведгруппы, передающего в зоне контроля функабвера, а его величина пропорциональна действенности сигналов[4]. Ровно то же случается при таком ухудшении погоды, снижении доступности корма и пр., из-за чего затраты на социальное поведение делаются неприемлемо высоки (а коммуникация и в норме — «предмет роскоши»).
Пример. Гуппи из разных популяций в речках Тринидада и др. местностей, различающиеся риском нападения хищника; самцы певчего воробья (п.13); райской птицы Paradisaea decora. копуляции в паре и внебрачные у певчих воробьиных и др. видов птиц; активация иммунитета от инфекции vs развитие брачных «украшений» и социальной активности у разных позвоночных; дихотомия «социальных» и «несоциальных» видов млекопитающих; дихотомия поведения жертв в «психологической дуэли» с хищником – окрикивать, совершенствуя соответствующие средства коммуникации, или тихо спасаться?

Райская птица Paradisaea decora
В. Между А- и Б-стратегией социального взаимодействия, дихотомия которых впервые описана В.В.Иваницким (1981) на близких видах сорокопутов, воробьёв и др. певчих воробьиных. В первом случае успех прошлых взаимодействий повышает возбуждённость участников, их взаимодействия ещё больше учащаются (и концентрируются в пространстве). Во втором случае, наоборот, прошлый успех ведёт к урежению воздействий, рассредоточению участников в пространстве и пр.
Соответственно А-стратегия снижает пороги всех форм демонстративного поведения («высокая эмоциональность» в социальной сфере). При Б-стратегии больший успех социальных взаимодействий снижает уровень общего возбуждения и поднимает пороги всех форм демонстративного поведения («низкая эмоциональность»). Поэтому первым присущи контагиозные группировки и интенсивные взаимодействия, вторым, напротив, разреженные с равномерным распределением участков, и «вялые» низкоинтенсивные взаимодействия, хотя и со столь же высокой стереотипностью демонстраций.
Пример. См. альтернативность социальных организаций белоспинного и большого пёстрого дятла; эффект воспитания у трёхиглой колюшки. Здесь потомство выращивается самцами, и у вида 2 формы: бентическая и лимнетическая. Второй стайность полезна, помогая избегать хищных рыб и пернатых хищников, первой безразлична. Опыты Kozak G.B. & Boughman J.W. (2013) по предъявлению макетов родителей или хищника, показывают, что лимнетическая молодь повышает стайность в ответ на те и другие, а также реальные атаки хищника, а бентическая ничего не меняет. И, самое главное: забота о молоди лимнетических отцов ведёт к повышению стайности потомства обеих форм. Козак и Боуман показывает, что разница в степени стайности и её модифицируемости в ответ на опасность или сигналы о ней – результат сочетания родительских эффектов, вызванной хищником пластичности и генетических различий между формами.
Г. Между попытками контролировать ситуацию в ответ на неблагоприятные события в месте пребывания животного или в ближайшем его окружении, резко перемещаясь в ответ в поисках нового и благоприятного места, или оставаться на прежнем, «приспосабливаться и терпеть».
На уровне поведения первое проявляется в том, что в ответ на проблемную ситуацию максимально интенсифицируется деятельность по активному изменению обстоятельств животного, чтобы уйти от проблемы, сменить среду и пр. Второе, наоборот — в максимуме пассивности, чтобы неблагоприятную ситуацию «вытерпеть» и перенести. («Подождать, пока бог переменит Орду»). Немного утрируя, первые больше и резче двигаются, вообще действуют, чтобы уйти от проблемы, вторые «на месте» меняют своё поведение (или меняются внутренне, физиологически), чтобы «приспособиться, не уходя».
Пример. Гаремные самцы vs рейдеры у благородных оленей; «Центральные» vs периферийные самцы у тетеревов и других видов, спаривающихся на току; доминантные гаремные самцы vs подчинённые у домовых мышей, пенсильванских полёвок (п.3) и ряда других видов грызунов; детёныши газели Томсона, замирающие и бросающиеся бежать (п.8); «Любовники» и «бойцы» у колюшек и пр.; «быстрые» или «медленные» особи у синиц, их гомологи – у грызунов, благородные олени в минуту опасности; особи, участвующие и нет в покатной миграции у рыб (см.ниже).
Д. Между прохождением особью разных стадий годового цикла (размножение, занятие территорий, зимовка и пр.) в одиночку или в составе плотных поселений, стай и других группировок. Скажем, самцы практически всех «токовых» видов могут занимать территорию на току или токовать одиночно.
Первые больше инвестируют в «брачные украшения» и другие эксцессивные (морфо)структуры, демонстрируют наибольшее внимание к брачным (и, шире) социальным сигналам при минимуме осторожности и большей подверженности гибели от хищника. В силу большей социабельности (социальной связанности и социальной зависимости, как это определяется у М.Е.Гольцмана) они наиболее подвержены групповому эффекту, эффекту массы и пр., но меньше стрессируются сигналами присутствия хищника и иными опасными ситуациями. Вторые, наоборот, наиболее «асоциальны»: у них максимум внимания к внешним опасностям, «указывающим» на них сигналам и пр., больше вложений в предостерегающую окраску. У них наиболее развиты нервные структуры, связанные с бегством от хищника при минимуме чувствительности к социальным влияниям или внимания к связанным с ними сигналам.
Важно подчеркнуть, что трейд-оффы А-Д а) существуют не для индивида, а для вида или популяции, б) однако они «задают индивидуальность особей», структурируя «пространство возможностей» для селективно-значимого разнообразия их в популяции. Индивидуальность здесь — точка в гиперпространстве осей координат, описывающих разнообразие индивидуальных стратегий внутри «типа». Сейчас факторы, определяющие индивидуальность особей, интенсивно изучаются, и с приращением материала становится ясно, что среди них практически нет собственно индивидуальных, не связанных со стратегиями, существующими в популяции. Соответствующая изменчивость если и есть, она не определяет биографию особей, их перспективы развития жизненного цикла и пр., тогда как «координаты» в параметрах А-Д детерминируют эти вещи почти полностью, см.примеры выше.
Как задано разнообразие стратегий: набором «ролей» в социальной системе или действительным изменением индивидуальности особей?
Последний важный вопрос – альтернативность стратегий в популяции отражает разнообразие индивидуальностей особей или наборы «ролей» в популяционной системе, в которые индивиды втискиваются, как в ложе Прокруста. Скорее второе, чем первое, и вот почему.
Изменчивость чисто индивидуальная часто встречается в популяциях позвоночных. Это и цветовые вариации у хищных птиц, скажем у канюка Свенсона[5] и других видов канюков (хотя не у всех, п.45), и разнообразие способов обучения аверсивной реакции у разных видов рыб, и т.д. Однако эта изменчивость всегда «расплывается» вокруг некого типа, как масляное пятно на бумаге, или вокруг нескольких типов, независимых друг от друга и не упорядоченных как противоположности.
Устойчивость индивидуальных различий в разные моменты времени основана на оценках повторяемости (repeatability) поведения[6]. В простейшем случае её оценивают корреляцией двух последовательных измерений одной и той же поведенческой характеристики в одном и том же тесте, разделённых большим или меньшим временем. В общем случае это внутриклассовый коэффициент корреляции (intra-class correlation coefficient), оцениваемый методом корреляционного или дисперсионного анализа, или на основе моделей смешанных эффектов. Так или иначе, поведенческие характеристики и «качества» особей тем больше характеризуют их индивидуальность как таковую (атомарные характеристики), чем выше повторяемость. И наоборот: чем повторяемость ниже, тем больше соответствующее поведение описывает роль особей в социуме и популяции (реляционные характеристики)
Мета-анализ исследований повторяемости поведенческих характеристик у рыб показывает, что она достаточно низка, в среднем 0.37 и обычно тем выше, чем короче временной интервал между измерениями. Так, у цихлиды Neolamprologos pulcher повторяемость смелости 0.83, если измерения делаются в течение одного дня, и падает до 0.19 для измерений, разделённых интервалом в три года. В целом при малых значениях повторяемости индивидуальные различия, однако, оказываются устойчивыми, но их величина сильно варьирует между популяциями одного и того же вида (и экологическими обстоятельствами).

Neolamprologos pulcher из оз.Танганьика.
Так, на заре современного интереса к индивидуальности животных (animal personality) в 1990-е годы в качестве основного критерия индивидуальных различий у рыб, D.S.Wilson c соавторами предложили континуум «пугливость — смелость» (shyness — boldness). Преобладание первого в проблемной ситуации толкает рыб к избеганию, второго — к пренебрежению риском и подходу к новым и потенциально опасным объектам.
Так Wilson et al. (1993) исследовали эту дихотомию у солнечных окуней Lepomus gibbosus; новыми объектами были а) длинная линейка, б) большой ставной невод, в) приближавшийся к рыбам пловец-наблюдатель. Более смелые особи меньше избегали а-в, быстрей акклиматизировались в лаборатории, но хуже объединялись в группы. Положение индивидов на градиенте пугливости-смелости оказывается предиктором особенностей их питания, предпочтительного использования определённых биотопов и (с ним связанных) особенностей паразитофауны.

Солнечный окунь Lepomus gibbosus
Сходные данные получены С.В.Будаевым на рыбах-зеленушках Symphodus ocellatus и ряде других видов. Более смелые особи активней исследовали, большей перемещались в новой незнакомой среде, предпочитали открытые биотопы и хуже объединялись в группы. Пугливые особи вместо исследований старались затаиваться, больше держались в зарослях и жили группами. Аналогичные межындивидуальные различия выявлены у птиц, вместе с социальными факторами, определяющими дифференциацию исходно однородных особей на склонных к риску как при исследовании, так и при демонстрировании, и пугливых, бегущих риска, с преобладанием тревожно-оборонительной реакции на новизну.
Смелость — пугливость индивидов у рыб оценивают или в тесте открытого поля (новая незнакомая среда, лишённая фиксированных ориентиров), или в предъявлении 1) незнакомых объектов разного размера и формы; 2) моделей хищника; 3) живого хищника за прозрачной перегородкой. При этом измеряется время до начала питания в присутствии хищника, или же до самостоятельного выхода рыбы из «домашнего» отсека аквариума в незнакомый отсек через малое отверстие в перегородке, время, проводимое в потенциально опасном открытом биотопе и пр.

Глазчатая зеленушка Symphodus ocellatus
Оценки смелости индивидов разными тестами довольно устойчивы, т.е. получившая высокую оценку в одном из них скорей всего будет высоко оцениваться и в других. Однако достаточно много исключений, когда особи, смелые в одном тесте, вели себя трусливо в другом (Coleman, Wilson, 1998; Wilson, Stevens. 2005; Dingemanse et al., 2007).
У рыб чётко выделяются альтернативные стратегии реагирования на стресс, т.н. копинг в ситуациях стресса[7] (coping with stress) — проактивную и реактивную, каждая из которых включает в себя когнитивные, эмоциональные и поведенческие компоненты, связанные между собой. Обе они используются особями, чтобы «взять себя в руки», справиться с ситуацией стресса. Рыб с проактивным реагированием на стресс отличает большая смелость, агрессивность, более высокий уровень активности вообще, большая шаблонность, стереотипность поведения, тенденция игнорировать небольшие изменения внешней стимуляции, т.е. дефицит пластичности. Новые и незнакомые объекты они обследуют быстро и поверхностно, исследовательское поведение проявляется быстро. «Реактивные» рыбы пугливей, неагрессивны, крайне чувствительны к изменениям внешней среды, их поведение наиболее пластично. Новые объекты обследуют медленно, но более тщательно.
Каждая из стратегий характеризуется своим поведенческим синдромом — коррелирующими между собой состояниями разных поведенческих характеристик (последние выступают признаками, по которым дифференцируются стратегии, а состояния признаком — набором оппозиций, по которым идёт дифференциация. Так, у разных особей трёхиглой колюшки уровни активности, смелости и агрессии коррелируют между собой. Также коррелируют степени пугливости и реактивности.
У млекопитающих и птиц характеристики темперамента, в том числе связанные со смелостью, в значительной степени определяются гормональными характеристиками реакции на стресс и стратегиями копинга (хотя процессы урбанизации «диких» видов и доместикации домашних вносят свои коррективы). То же самое обнаружено у Brachyraphis episcopi и других видов рыб. Большая смелость индивидов сопрягается с относительно слабой гормональной реакцией на стресс. У карпа Cyprinus carpio смелые особи отличались существенно большей интенсивностью метаболизма, чем пугливые. У них были понижены уровни экспрессии гена рецептора кортизола, концентрации лактата и глюкозы в плазме. У японского горбыля Argyosomus japonicus смелые особи тоже имели существенно меньшую концентрацию кортизола, чем пугливые.
У плотвы и ряда других видов рыб выделены альтернативные стратегии, связанные с участием/неучастием особей в покатной миграции. В этот период молодь плотвы чётко дифференцирована на две группировки, резидентов и мигрантов. Им соответствуют особи с низким и высоким содержанием веществ катехоламинэргической системы (ДОФА, дофамин) и кортизола, а промежуточных не бывает. Молодь из «низкой» группы обитает прибрежных биотопах, не выходит на стремнину и не участвует в покатной миграции. Молодь второй группы держится на течении ближе к русловому потоку, стрессируется реоусловиями, для адаптации к ним повышает содержание гормональных веществ и т.д. В конце концов это ведёт к их дифференциации как мигрантов и участию в перемещении по реке.
Однако же здесь, наоборот, смелее оказались рыбы с более интенсивной гормональной реакцией на стресс, они с большей вероятностью выходят в новую среду и пр. Что ещё раз подтверждает сформулированный нами принцип социальной релятивности действия гормонов: если в данном конкретном случае дифференциации жизненных стратегий большие концентрации соответствующих веществ обеспечивают миграцию, т.е. перемещение в неизвестность и новые условия, они будут связаны с большей смелостью, в противоположность видам, все особи которых живут на одном и том же месте всё время. Особи резидентной группы в реоусловиях могут находиться лишь очень короткое время, ибо не выдерживают развивающейся стресс-реакции. Мигранты же справляются за счёт интенсификации синтеза катехолмаинов и стероидов, регулирующих энергетические процессы в организме.
Работа Øverli et al. (2005, 2007) описывает систему различий между выведенными линиями микижи Parasalmo mykiss с высокой и низкой реактивностью на стресс, HR и LR. Критерием отбора была высокая (HR) или низкая (LR) постстрессовая концентрация кортизола в плазме крови. Эти две линии были сильно различны по физиологии и поведению в различных тестах, так что исследовали сделали вывод о различиях в темпераменте. Рыбы HR отличались развитием стресс-индуцированной анорексии, не питаясь в постстрессовом периоде. LR отличались большей смелостью, агрессивностью, обычно занимали верхние ранги во внутригрупповой иерархии. Условный рефлекс на выученный стресс у них затухал сильно медленней, чем у HR. Иными словами, низкая реактивность на стресс здесь была связана с низкой пластичностью, большей смелостью и агрессивностью и наоборот, что согласуется с аналогичными данными по птицам и млекопитающим.
Различия между линиями микижи также захватывали профили активности и метаболизма моноаминэргической системы мозга. HR отличал больший уровень синтеза и метаболизма серотонина, норандреналина и дофамина в постстрессовый период. LR характеризовались повышенной концентрацией метаболитов серотонина 5-HIAA и норандреналина MHPG в гипоталамусе, в их переднем мозгу было выше соотношение 5-HIAA/5-HT. Что в целом соответствует альтернативным стратегиям копинга у млекопитающих (Øverli et al., 2007).
У нас и других млекопитающих в контроле эмоций и формировании индивидуальности важную роль играет лимбическая система, включающая гиппокамп, миндалину, гипоталамус и несколько близлежащих структур. У рыб, по всей видимости, этим занимаются её гомологи — латеральный и медиальный паллиум; показан их контроль, например, над эмоциональным обучением. Сейчас появляются данные о ключевой роли в формировании эмоций и индивидуальных различий в поведении и латеральной асимметрии у рыб ряда структур эпиталамуса, в первую очередь хабенулы. Это один из основных компонентов промежуточного мозга, соединяющего лимбические отделы переднего мозга со средним и задним. У рыб и ряда других позвоночных она резко асимметрична, левые латеральные ядра значительно превышают правые, а мутация, изменяющая эту асимметрию, резко увеличивает смелость у данио-рерио. Развитие эмбрионов этого вида в полной темноте также влияет на асимметрию хабенулы и одновременно на смелость.
Так или иначе хабенула, как и другие названные отделы, от рыб до млекопитающих, «делают» индивидуальность, модулируя дофаминэргические системы мозга. Они, в свою очередь, играют важную роль в формировании тревожности и страха, и связанных с ними характеристик индивидуальности, почему находятся под сильным социальным влиянием (у рыб, правда, его лишь начинают исследовать, а у млекопитающих разбирают уже в подробностях).
Однако и у рыб выраженность этих противоположностей в значительной степени зависит от внешней и внутренней среды обитания популяции (иногда полностью). Так, отмеченные Wilson et al., 1993 различия между смелыми и пугливыми солнечниками, значительные в природе, полностью исчезали при продолжительной изоляции в аквариуме, хотя наблюдались сполна в первые моменты после помещения туда. Повторяющиеся имитации атаки хищника повышали уровень смелости рыбок Brachyrhaphis episcopi, происходящих из популяций как с высоким, так и с низким уровнем хищничества. Обыкновенные окуни из популяций с высоким и низким уровнем хищничества существенно отличались смелостью, но эти различия исчезали при выращивании в среде без хищников.

Пецилия Brachyrhaphis episcopi
Далее, изменение состава группы окуней значительно модифицирует поведение пугливых особей (они более социабельны, т.е. больше зависимы от социальной стимуляции) и в меньшей степени смелых. Пугливые рыбы становились смелей в группе таких же, а смелые в группе смелых, наоборот, снижали уровень смелости, т.е. происходила регрессия к среднему. В целом для окуней как вида корреляция между временем, проводимым в потенциально опасной среде и исследованием незнакомой среды (что и формирует характеристику смелости) оказывается статистически значимой только при учёте поведения других членов группы.
У микижи уровень смелости модифицируется как опытом победы-поражения в агрессивных взаимодействиях, и даже простым наблюдением за поведением смелых или пугливых особей своего вида (Frost et al., 2007). После нападения воздушного хищника самцы гуппи проявляют большую смелость при нахождении в группе самцов, а не самок (у этого вида различия по пугливости-смелости входят в структуру полового диморфизма). И даже созданные отбором различия стратегий копинга у линий микижи с высокой и низкой реактивностью на стресс спонтанно реверсируют при смене среды. После переселения из Великобритании в Норвегию рыбы HR демонстрировали большую смелость, чем LR и стали доминировать над ними во взаимодействиях. Самое важное, что различия гормональных профилей между линиями не изменились (Ruiz-Gomez et al., 2008). По-видимому, повлияла процедура транспортировки и другие условия содержания.
Поэтому альтернатива пугливости-смелости и связанные ними стратегии копинга связаны больше с дихотомией ролей в популяционной системе, чем с чисто индивидуальными различиями.
Напротив, различия в способности рыб к обучению, видимо, отражают чисто индивидуальную вариацию. Так, особи сёмги и гуппи существенно различались по скорости обучения двустороннему избеганию в челночной камере. Безусловным стимулом здесь служил удар тока, условным — включение лампочки, надо было выучить, что в ответ на них «следует» переместиться в противоположный отсек камеры.
В то время как часть особей быстро выработали правильное избегание, другие были к этому неспособны и быстро переходили к пассивно-оборонительному поведению, больше всего похожему на выученную беспомощность. Третьи вырабатывали оборонительную реакцию, но нестойкую и быстро затормаживающуюся. Четвёртые же демонстрировали быстрое обучение, где устойчивый условный рефлекс появлялся без предшествующего периода неустойчивых. Однотипные паттерны индивидуальных различий найдены у других видов рыб и, видимо, универсальны.
Скорость обучения гуппи в данных экспериментах значительно связана с уровнем смелости, причём менее смелые рыбки учатся быстрее. Что хорошо согласуется с двухпроцессной теорией избегания А.Ю.Жуйкова (1995). В соответствии с ней общая скорость обучения определяется скоростью выработки реакций «выученного страха» (conditioned fear) и потом инструментальной реакции бегства в противоположный отсек. По-видимому, трусливых особей отличает легкая выработка выученного страха в сочетании с интенсивной двигательной реакцией в ответ на аверсивный стимул, почему они сразу боятся и сильно бегут. Хотя в популяции есть все возможные сочетания. К слову, эта модель позволила закрыть тему «Есть ли инсайт у рыб?»; то, что раньше считалось инсайтом, сполна объясняется этой более простой моделью, т.ч. её следует предпочесть в соответствии с каноном Конвей Ллойда-Моргана[8].
Таким образом, «чисто индивидуальной» вариацией выступает трейд-офф между скоростью и тщательностью обучения, описанный у разных видов рыб (и, видимо, других позвоночных). Выраженность обоих параметров образует непрерывный градиент, вдоль которого первое убывает, а второе растёт, но не концентрируется у противоположных полюсов.
А там, где есть система из двух или нескольких альтернативных стратегий, образующие их признаки и качества особей всегда жёстко «пристёгнуты» к участию в максимизации репродуктивного выхода популяции, долгосрочной устойчивости её пространственно-этологической структуры и пр. через стабилизирующий отбор. Почему и должны рассматриваться как характеристики не самой индивидуальности животного, а роли его как носителя определённой стратегии (семафоронта) в надындивидуальной системе.
Заключение
Вышеприведённый анализ (хорошо согласующийся с рядом других аргументов) показывает необходимость не индивидо-, а системоцентрического взгляда на процессы естественного отбора в популяции, где объектом отбора являются не индивиды, а поведенческие модели (стратегии), перспективные и не очень, которые с разной точность и разным качеством могут реализовываться разными индивидах, сталкивающимися друг с другом в конкурентной борьбе.
Эта реализации и столкновения служат для «статистического испытания» поведенческих моделей и отбора лучших, с последующим тиражированием во всей популяции. Люди же, в смысле отдельные индивиды, здесь существенны лишь как семафоронты, носители более или менее перспективных моделей, способные делать выбор — запускать или нет их реализацию в соответствующей жизненной ситуации. В этих условиях социальное родство ничуть не хуже, а по ряду параметров и лучше биологического, ибо «хоть какое-то» исполнение перспективной модели будет достигнуто за счёт обучения, подражания и собственного интеллекта исполняющего, что уже даёт этому последнему выигрыш перед теми, кто реализует «неперспективную модель».
А дальше включается эффект Болдуина и «подгоняет» физиологию с биологией индивидов-исполнителей «лучшей» модели так, чтобы эта последняя срабатывала всё эффективней и эффективней.
Примечания
[1] Hogg J.T., 1984. Mating in bighorn sheep: multiple copulative male strategies// Science. V.225. №4661. P.526-529.
[2] См. «Сила есть, ума не надо, и наоборот»; Про альтернативные стратегии разрешения проблемной ситуации; Популяционная регуляция после истребления у «социальных» и «несоциальных» видов грызунов;
[3] См. подробнее «Про биологический смысл мастурбации у приматов»
[4] См. «Ещё про знаки и символы в коммуникации животных»
[5]Вид включает 3 морфы: светлую (7% гнездящейся популяции), среднюю (57%) и темную (36%), со всеми возможными переходами. Анализируя данные за 32 года, Briggs C.W. et al. (2011) пытались найти связанные с ними различия в выживаемости, успешности гнездования, общем количестве молодых за год, возможности вовлечения молодых в гнездящуюся популяцию и репродуктивном успехе в течение всей жизни. Очень искали и жали этих различий, но не нашли, и сделали вывод, что ни апостатичный отбор, ни гетерозиготность несущественны как детерминанты индивидуального полиморфизма.
[6]Здесь и далее данные по индивидуальности рыб, где не приведены ссылка, пересказаны по обзору: Будаев С.В., Михеев В.Н., Павлов Д.С., 2015. Индивидуальные различия в поведении и механизмы экологической дифференциации на примере рыб»// Журнал общей биологии. Т.76. №1. С.26-47. См. также пересказ публикаций С.В.Будаева «Про индивидуальность поведения рыб».
[7] Калька с английского здесь особенно неудачна, т.к. дезориентирует. Имеется в виду то, что человек или животное делают, чтобы справиться со стрессом, на ум же приходит неуместное здесь «копирование»
[8] См. «Безопасная бритва Оккама»
Источник Этология.ру
Рекомендуем прочесть
 Итоги реконструкции выборов в Учредительное собрание
Итоги реконструкции выборов в Учредительное собрание Биология как идеология. Часть 3
Биология как идеология. Часть 3 Некоторые пути сохранения биосферы при урбанизации
Некоторые пути сохранения биосферы при урбанизации Механизм становления отцовской заботы о потомстве
Механизм становления отцовской заботы о потомстве- Биология как идеология. Часть 4
 «Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве
«Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве Современное изучение городской климатологии
Современное изучение городской климатологии Дивергентно-конвергентные механизмы в эволюции биоразнообразия
Дивергентно-конвергентные механизмы в эволюции биоразнообразия
