Гиляров А.М.
![]()
В современном языке слово «экология», предложенное Э. Геккелем в 1866 г. для обозначения раздела биологии, изучающего взаимоотношения организмов и среды, стало своего рода расхожим штампом, бездумно использующимся в столь многих значениях, что первоначальный смысл его кажется почти утраченным. Данное обстоятельство вовсе не способствует формированию в обществе уважительного отношения к экологии как науке, и, видимо, не случайно свою статью, публикуемую в академическом журнале, профессиональный эколог А.Ф. Алимов назвал «Об экологии всерьез» [1], а другой профессиональный эколог Г.А. Заварзин в «Лекциях по природоведческой микробиологии» специально разъяснил, что избегает слова «экология», ассоциирующегося у него с «экодемагогией» [2]. Полностью разделяя обеспокоенность коллег относительно спекуляций на термине «экология», автор все же не может без него обойтись. Речь пойдет об экологии как о вполне серьезной науке, исследующей живой облик биосферы, пытающейся его всесторонне охарактеризовать и понять, какими механизмами он поддерживается.
Как отдельная, самостоятельная научная дисциплина экология сложилась в 20-х годах XX в., когда появились люди, называющие себя экологами, были основаны экологические научные общества и стали выходить соответствующие специализированные журналы. Сначала экология была преимущественно эмпирической и описательной наукой, пытающейся собрать и как-то упорядочить огромное количество данных, касающихся распределения и динамики численности различных организмов, а также существования различных надорганизменных систем — популяций, сообществ, экосистем и всей биосферы.
Очевидно, однако, что определенная теоретическая или, выражаясь более осторожно, концептуальная база экологии всегда существовала, поскольку даже «простое» описание наблюдаемого не может не быть «теоретически нагруженным». (Как справедливо заметил в свое время К. Поппер, невозможно «просто наблюдать», надо по крайней мере решить заранее, за чем наблюдать и как [3].) При этом сами экологи соглашаются с тем, что хорошей теории — достаточно универсальной и вместе с тем делающей более или менее надежные прогнозы — в их науке никогда не было.
На этом фоне удивительным кажется тот прогресс, который наблюдался в экологии за минувшее столетие и проявился не только в увеличении объема накопленной эмпирической информации, но и в достижении более глубокого понимания того, как формируются и поддерживаются определенные структуры (паттерны) в устройстве популяций, сообществ и экосистем. Возникает вопрос: благодаря чему это понимание стало возможно, или как экология могла развиваться без «хорошей» теории, не будучи при этом заблокированной огромным количеством новых эмпирических данных. Можно дать два ответа, причем не исключающих, а дополняющих друг друга.
Во-первых, мягкость и гетерогенность экологической теории, хотя и свидетельствовали о ее слабости, или, по меньшей мере, незрелости, в то же время имели положительное значение, позволяя охватить и «переварить» множество сырых данных и гипотез ad hoc (порой весьма противоречивых) [4]. Уточним также, что требования экологов к теоретическому обоснованию поднимаемых вопросов были совсем не завышенными, а вместо настоящей теории они нередко довольствовались определенными традициями сбора и обработки материала, сложившимися в той или иной конкретной области экологии или даже в отдельной научной школе.
Во-вторых, в самом теоретическом базисе экологии на протяжении XX в. произошли серьезные изменения, и если в 20-х годах важная роль еще принадлежала аналогиям, заимствованным из других наук, в частности, физической химии или экспериментальной эмбриологии, то к концу столетия на первое место стали выходить собственно экологические принципы, следующие из таких элементарных, свойственных всем организмам, свойств, как размножение, экспоненциальный рост численности и наличие ограничений, накладываемых на этот рост средой и структурно-функциональными особенностями организмов. Сдвиг акцентов отразился и на современном операциональном определении экологии как науки о «взаимодействиях, определяющих (де факто -«ограничивающих». -А.Г.) обилие и распространение организмов» [5, 6].
Называние видимого, панглосская парадигма и детерминизм.
Содержание
Прослеживая становление экологии как самостоятельной науки, мы отмечаем в ней несколько корней, уходящих в научные традиции XIX и даже XVIII в., в частности, такую своеобразную форму упорядочивания знаний о живой природе, как естественная история. Французский философ М. Фуко метко охарактеризовал естественную историю как «называние видимого» [7]. Действительно, описание и классификация огромного разнообразия видов растений и животных происходили именно в рамках парадигмы, или, используя терминологию Фуко, — «эпистемы» естественной истории. Отметим, что это «называние видимого» признавалось абсолютно самодостаточной задачей, хотя уже со второй половины XVIII в., начиная с работ И. Канта, в европейском естествознании стала быстро развиваться другая эпистема, ориентированная на распознание скрытого. В частности, в рамках этой новой эпистемы в начале XIX в. появились работы по географии растений одного из крупнейших предтечей экологии — А. Гумбольдта [8].
Экология, как и естественная история, в течение долгого времени преимущественно описывала «видимое» и также имела дело с огромным разнообразием организмов и разнообразием биотопов. Экспериментальные работы на «модельных» видах, например, на мучных жуках Tribolium или ветвистоусых ракообразных Daphnia, хотя и проводились, но касались они только некоторых проблем динамики популяций и никогда не заменяли исследований, имевших дело с множеством разных видов.
Минимальные объекты экологов — это целые организмы, рассматриваемые порой как своего рода «черные ящики», для которых важно, что «на входе» и что «на выходе» (то есть что и в каком количестве организм потребляет и выделяет), а не то, что «внутри» (предмет других наук). Любопытная параллель: классическая естественная история, как подчеркивает Фуко, повышенное внимание уделяла именно внешнему облику организма, а не тому, что требует вскрытия для понимания устройства, поэтому именно с изучением царства растений связаны основные успехи естественной истории.
В качестве примера настоящего рудимента естественной истории, существующего и в современной экологии, можно привести выделение и называние растительных сообществ в соответствии с методикой, предложенной в конце 1920-х годов швейцарским исследователем Ж. Браун-Бланке [9]. Суть этого подхода в том, чтобы с помощью специальных приемов (граничащих, по Браун-Бланке, с искусством), которые должны передаваться от учителя к ученику на совместных экскурсиях в мир природы, уметь распознать идеальные ассоциации — совокупности устойчиво встречающихся видов, и дать им названия, аналогичные тем, что используются в классической таксономии организмов. При этом практически провозглашается отказ от попыток изучить и понять причинную обусловленность пространственно-временного распределения отдельных видов (например, зависимость их от типа почв, увлажнения, присутствия конкурентов и т.п.). Удивительно, что данный подход, ориентированный не на объяснение природы, а исключительно на ее описание, причем в рамках незыблемых канонов, до сих пор практикуется некоторыми фитоценологами [10,11].
Своими мировоззренческими корнями естественная история уходит в креационистскую доктрину. Разнообразие животных и растений трактовалось в ее рамках как результат продуманных действий Творца, уготовившего каждой группе организмов свою роль. Например, хищники должны сдерживать рост травоядных животных, чтобы те в свою очередь не смогли съесть все растения. Также «сверху» задавалось и природное равновесие, на поддержание которого специально направлена деятельность тех или иных организмов. Мысль о том, что все в природе имеет определенное предназначение и (или) является адаптацией, как ни странно, прослеживается порой и в современной биологии. По отношению к строению и функционированию организмов подобный подход Гулд и Левонтин [12, 13] метко назвали «панглосской парадигмой» в честь известного персонажа Вольтера, но с не меньшими основаниями это язвительное, но по сути справедливое определение может быть отнесено и ко многим рассуждениям в экологии, касающимся адаптивности тех или иных особенностей устройства надорганизменных систем (например, о «биоцено-логической роли» хищников и паразитов).
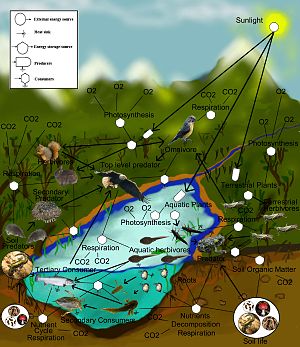
Mathematical models developed in theoretical ecology predict complex food webs are less stable than simple webs. Источник.
Путаница с адаптивностью нередко возникает из-за того, что ее ищут не там, где она есть. К примеру, сейчас описано перемещение готовых продуктов фотосинтеза между деревьями разных видов по мицелию соединяющего их гриба, причем преимущественно в сторону того дерева, которому не хватает ресурсов (например, испытывающего затенение) [14]. Сторонники «панглосской парадигмы», скорее всего, будут трактовать такое проявление взаимопомощи между разными видами растений (конкурирующих за одни и те же ресурсы) как «адаптацию» лесного сообщества, направленную на поддержание своей целостности. Но гораздо проще объяснить это как побочный результат вполне «эгоистичной» эволюции гриба, для которого выгодно образовывать микоризу с разными деревьями для обеспечения своего выживания и размножения.
К естественной истории, а точнее — к мифологическому мышлению, элементы которого еще сильно сказывались в XVIII в. и не изжиты в настоящее время [15], восходит и традиция жесткого детерминизма, дающая о себе знать прежде всего в учебной и популярной литературе по экологии. Характерный пример — один из так называемых «законов Коммонера», сводящийся к утверждению, что «всё связано со всем». Но любая экологическая система — от популяции до биосферы в целом — не могла бы существовать и уж тем более эволюционировать, если бы была устроена таким жестким образом. Функционировать и развиваться подобные системы могут только благодаря вариабельности их элементов, стохастичности многих событий и определенной (хотя и ограниченной) автономии организмов от окружающей среды.
Подвижное равновесие, «физическая биология» и органицизм.
В начале XX в. в кругах исследователей, стоящих у истоков экологии, весьма популярны были умозрительные построения Г. Спенсера [16-18], касающиеся общих принципов организации систем, энергетического подхода, а также подвижного равновесия, возникающего в результате взаимодействия разнонаправленных процессов. Наиболее фундаментальными процессами считались эволюция, понимаемая как процесс интеграции вещества, сопровождаемый рассеиванием движения, и противоположная ей «дезинтеграция», трактуемая как поглощение движения, сопровождаемое деструкцией вещества. В использованной терминологии очевидно влияние успехов термодинамики. Неудивительно также, что идея подвижного равновесия нашла свое применение прежде всего в химии, где получила название принципа Ле-Шателье.
Возникшая в конце XIX в. физическая химия также широко использовала термодинамику и энергетический подход. Один из основателей новой науки — В.Ф. Оствальд — надеялся даже с его помощью придать химии надлежащую строгость, а в начале XX в. другой исследователь — А. Лотка — под влиянием работ Оствальда предпринимает попытку упорядочить биологические знания согласно принципам физики. В 1925 г. он публикует книгу «Элементы физической биологии» [19], содержание которой находится на стыке физики, химии, биологии и философии. Фактически это вариант общей теории систем в понимании А.А. Богданова или в современной трактовке А.Л. Тахтаджяна [20]. Своим объектом физическая биология, по мысли ее автора, принципиально отличалась от тогда весьма уже популярной биофизики: если биофизика имела дело с отдельными организмами, то физическая биология должна была анализировать более крупные и целостные системы, включавшие комплексы организмов, связанные со средой.
Встроенность живых организмов (в том числе человека) в некую большую целостную систему также особо подчеркивалась Лоткой. Сравнивая развитие такой большой системы со спектаклем, он замечает, что «наше внимание должно концентрироваться не только на персонажах пьесы, но и распространяться на сцену, из которой они появились, на которой исполняют свои роли и с которой вскоре вновь соединятся» [19, с. 184]. Очень важное место Лотка уделял эволюции, но трактовалась она не в биологических терминах, а скорее, термодинамических, как история системы, претерпевающей необратимые изменения. Биологи же, по его мнению, неоправданно много внимания уделяли частным аспектам эволюции (например, филогенетическим линиям), упуская из виду закономерности, применимые только к системе в целом. (В последнее время такое биосферное понимание эволюции активно развивает Г.А. Заварзин [2], используя при этом много новой эмпирической информации.) Хотя сам Лотка слово «экология» практически не употреблял, несколько влиятельных американских ученых, связывающих себя с экологией (в частности, Р. Перль и Ч. Эдамс), быстро оценили значение его работ для становления новой науки [18]. С позиций сегодняшнего дня значимость этих исследований для дальнейшего развития экологии несомненна.

Во-первых, Лотка одним из первых использовал математические модели для изучения популяционных процессов, в частности, исследовал стабильное возрастное распределение, устанавливающееся в экспоненциально растущей популяции, и предложил систему дифференциальных уравнений, описывающих динамику взаимодействующих популяций (связанных отношениями «хищник-жертва» или конкуренцией за общие ресурсы). Именно с работ Лотки и независимо выполненных исследований итальянского математика В. Вольтерры началось активное использование математических моделей для выявления универсальных закономерностей динамики численности. Заметим, что многочисленные ссылки на Лотку в экологической литературе второй половины XX в. в большинстве случаев касались именно уравнений «хищник — жертва», хотя по сути это были только небольшие детали в развиваемой им системе представлений об устройстве и эволюции надорганизменных образований.
Во-вторых, Лотка в своей книге уделил много внимания циклам биогенных элементов и их взаимодействиям, перекликаясь здесь с идеями Вернадского (хотя оба исследователя развивали свои концепции самостоятельно и ограничивались лишь формальными ссылками на работы друг друга). Геохимическая деятельность человека рассматривалась Лоткой как естественный процесс, направленность которого совпадала с общей направленностью эволюции на увеличение массы вовлеченного в биогенный круговорот вещества и возрастание скорости самого круговорота. Надо подчеркнуть, что оказав огромное влияние на развитие в экологии популяционного подхода (то есть изучение динамики численности организмов), работы Лотки практически не имели значения для становления экосистемного подхода, ориентированного на изучение потоков вещества и энергии. Формирование последнего происходило в другом русле и связано было прежде всего с успехами гидробиологии и лимнологии, в том числе и достигнутыми в 1930-х годах российскими исследователями (Л.Л. Россолимо, Г.Г. Винбергом, СИ. Кузнецовым, B.C. Ивлевым и др.).
В-третьих, книга Лотки и неожиданный (по крайней мере, для самого автора) интерес к ней экологов свидетельствуют, что по сути своей экология очень близка к общей теории систем. Недавно на это справедливо указал В.А. Красилов [21], подчеркнувший, что уже с момента определения новой науки Геккелем ее объектами стали системные взаимоотношения, а не материальные тела, формы движения или процессы.
Помимо идей, пришедших в экологию из физической химии и общей теории систем, важное влияние на ее формирование всегда оказывали представления, развиваемые в рамках самой биологии при изучении живого организма. Использование последнего как некой модели при исследовании других объектов, не являющихся организмами в узком смысле слова, — это основа методологии органицизма. В качестве таких уподобляемых организму объектов чаще всего выступали сообщества и экосистемы (реже — популяции), а в конце XX в., в рамках развиваемой Д. Лавлоком концепции Геи, — вся биосфера. Особенно популярны идеи органицизма были в фитоценологии [22], где уже в начале века Ф. Клементс при описании закономерной смены растительных сообществ подчеркивал, что сообщество рождается, развивается, взрослеет и отмирает. Причем если на начальных стадиях сукцессии возможны значительные колебания в видовом составе и численном соотношении разных видов, то к зрелому состоянию (климаксу) все сообщества определенной местности становятся сходными, демонстрируя тем самым эквифинальность — достижение одного конечного состояния, несмотря на возможные нарушения в начале.
До этого эквифинальность была известна из работ по экспериментальной эмбриологии (механике развития), но достижения последней были в 1920-1930-х годов очень популярны среди биологов, в том числе и тех, кто стоял у истоков экологии. Детерминизм развития рассматривался Клементсом как очень важная черта, сближающая растительное сообщество с отдельным организмом. В некоторых фитоценологических школах, в том числе в отечественных, такая жесткая детерминистская позиция сохранялась довольно долго, что на самом деле мешало проникновению в экологию идей о стохастичности многих процессов.
Борьба за существование, индивидуалистический подход и «трейдоффы».
Из приведенного краткого обзора видно, что первоначально идеи, вокруг которых формировалась теоретическая база экологии, были заимствованы из других наук. Подвижное равновесие и принцип Ле-Шателье перешли в экологию из общей теории систем и физической химии. Выделение группировок растительности основывалось на классической таксономии, уходящей корнями в естественную историю, а развитие сообщества рассматривалось по аналогии с онтогенезом отдельного организма — объектом «механики развития». Все эти параллели были полезны для накопления и упорядочивания эмпирического материала. Но вместе с тем создается ощущение, что глубинные процессы, определяющие живой облик биосферы, оставались в тени. Взаимоотношения организмов (одного или разных видов), хоть и признавались очень важными, практически почти не изучались.
Кроме того, образ природы, рисуемый экологией первой трети XX в., был в высшей степени статичным, соответствующим старым, но не стареющим представлениям о всеобщем природном равновесии. Эволюционное начало в ней практически отсутствовало. И это при том, что Геккель, предлагая термин «экология», находился под сильнейшим впечатлением от «Происхождения видов» и даже надеялся, что предметом новой науки станут «все те запутанные взаимоотношения, которые Дарвин условно обозначил как борьбу за существование» [23].
Свою неудовлетворенность отрывом экологии от эволюционных идей, и конкретно от дарвиновских представлениях об отборе, выразил в 1930 г. молодой, но уже известный английский эколог Ч. Элтон [24]. Он подчеркнул, что если для вида существует оптимальный уровень плотности популяции, то естественный отбор должен способствовать формированию у этого вида особенностей, позволяющих поддерживать данный уровень. Однако, рассуждая дальше, автор приходит к заключению, что практически такое невозможно, поскольку оптимальный уровень плотности должен все время изменяться, чтобы быть «оптимальным» (ведь меняются плотности популяций других видов, с которыми данному виду приходится взаимодействовать, не говоря об абиотических условиях).
Противоречия, указанные Элтоном, впоследствии отчасти были разрешены или, по крайней мере, переформулированы в понятиях, позволяющих приблизить их разрешение. Так, уже в 50-х годах. Д. Лэк задался вопросом: каким образом у птиц формируется оптимальный размер кладки, то есть то количество откладываемых яиц, которое обеспечивает стабильное существование популяции, компенсируя смертность в последующем возрасте. А вышедшая в 1993 г. книга Э. Чарнова об инвариантах жизненных циклов [25] открывалась разбором таблицы, в которой приведены данные для разных видов птиц по годовой плодовитости (числу отложенных яиц в расчете на одну самку) и смертности взрослых. Из таблицы следовало, что обе величины, широко варьирующие от вида к виду, теснейшим образом связаны между собой, а один и тот же результат — поддержание из поколения в поколение более или менее стабильной численности — может достигаться как при крайне низкой плодовитости (например, у альбатроса), так и очень высокой (некоторые воробьиные). Вопрос об оптимальной плотности популяции постепенно заменился вопросом об оптимальной плодовитости, а этот признак уже гораздо легче трактовать как результат классического дарвиновского естественного отбора.
В 1934 г. в США публикуется книга, само название которой — «Борьба за существование» — являлось ссылкой на Дарвина [26]. Впрочем, автор — молодой московский зоолог Г.Ф. Гаузе — акцентировал внимание вовсе не на эволюции и даже не на эволюционных последствиях экологических взаимодействий. Его задачей было проанализировать элементарные процессы, протекающие в растущих популяциях как одного вида, так и разных, находящихся в отношениях конкуренции или «хищник-жертва». Подход Гаузе базировался на сочетании математических моделей, которые тогда уже были предложены другими исследователями (но изложены в книге в более понятной для биологов форме), и данных, полученных автором в ходе проведения изящных лабораторных экспериментов с бактериями, дрожжами и простейшими. С позиций сегодняшнего дня самый главный результат этих исследований — наглядная демонстрация того, что основные процессы, определяющие существование и взаимодействие популяций, не настолько сложны, чтобы их нельзя было изучать с помощью простых математических и лабораторных моделей.
Для экологии это был настоящий триумф редукционизма — методологии, до этого уже нашедшей успешное применение во многих областях естествознания. Вопрос, однако, в том, были ли сами экологи готовы безоговорочно принять новый подход? Видимо, скорее, нет, чем да. Об этом свидетельствует хотя бы то, что многочисленные ссылки на книгу Гаузе в течение нескольких последующих десятилетий касались исключительно принципа конкурентного исключения, то есть утверждения, что два вида не могут устойчиво сосуществовать, если занимают одну и ту же экологическую нишу1.
1Вокруг этого принципа было много дискуссий среди экологов-зоологов, ведущих наблюдения в природе. Некоторые авторы не без оснований считали его тавтологией и указывали на то, что он не может быть фальсифицирован, ведь для любых видов, в том числе сосуществующих, всегда можно найти какие-то различия и, соответственно, «списать» на них сосуществование [27].
Принцип этот, бесспорно, сыграл важную роль в сборе и трактовке материала по экологии родственных видов [28], но все же имел достаточно частный характер. Гораздо важнее была сама методология, дающая возможность практически изучать элементарные (именно поэтому основополагающие и универсальные) механизмы динамики популяций. Существенно и то, что использованный подход не сводился к апостериорному описанию результатов наблюдений, а предполагал анализ происходящих процессов, опирающийся на некоторые априорно принимаемые модели, например, предположение об экспоненциальном или логистическом росте популяции.
Работы Гаузе, выполненные в 30-х годах, не были изолированным явлением в науке того времени. Самым непосредственным образом они продолжали сделанное предшественниками, среди которых были и теоретики (Перль, Вольтерра, Лотка), и экспериментаторы (Чепмэн), и натуралисты, ведущие наблюдения в природе (А.Н. Формозов, Элтон). В последующие годы экспериментальные исследования с растущими популяциями (чаще всего мелких членистоногих) также время от времени проводились, а результаты некоторых даже попали в учебники, однако в целом использованная Гаузе методология имела ограниченное распространение. Подавляющее большинство работ в экологии были направлены на описание наблюдаемого, без серьезных попыток причинно-следственного анализа той или иной конкретной ситуации.
Меняться ситуация начала только в конце 70-х-начале 80-х годов, когда регулярно стали появляться работы, направленные на изучение механизмов ограничения роста популяций. При этом анализ динамики численности организмов в природе все чаще предполагал подробные наблюдения за изменениями не столько самой численности, сколько рождаемости и смертности, то есть популяционных показателей, непосредственно реагирующих на внешние факторы. В ходе экспериментальных работ удалось также выяснить, что конечный результат взаимодействия разных видов нередко может быть предсказан на основе их характеристик, полученных при изолированном культивировании, причем скорость популяционного роста не является решающим показателем2.
2Данное обстоятельство следует подчеркнуть особо, поскольку в популяционной генетике в качестве показателя приспособленности, как правило, использовался мальтузианский параметр (r), и тогда становилось непонятным, как вообще могут существовать виды, размножающиеся медленнее, чем их сородичи, живущие рядом.
Так, прогнозируя исход конкуренции между разными видами планктонных водорослей [29], исследователи использовали величину пороговой концентрации лимитирующего ресурса, то есть той минимальной его концентрации, которой достаточно для поддержания популяции (сохранения равенства рождаемости и смертности).
Редукционистский подход, столь ярко проявившийся в работах Гаузе 30-х годов, но окончательно утвердившийся в экологии только в 80-е годы, на самом деле возникал в разных ее областях и раньше, но не получал развития, поскольку не укладывался в господствующую парадигму. Так, фактически всю первую половину XX в. специалисты, изучающие растительный покров, рассматривали его как совокупность отдельных сообществ, а каждое сообщество — как высоко интегрированное целостное образование, порой сравниваемое с организмом. Такая позиция, в частности, особо активно отстаивалась одним из лидеров тогдашней американской экологии Ф. Клеменсом.
На этом фоне совершенно еретической выглядела позиция другого американского эколога — Г. Глизона [30], который в 20-е годы выдвинул так называемую индивидуалистическую концепцию, противопоставив ее трактовке сообщества как аналога организма или таксономического вида. Основное внимание исследователей, по мысли Глизона, должно быть направлено на распределение особей (индивидуумов) того или иного вида и выявление определяющих его факторов. Сообщество же рассматривалось им как результат совпадения в распределении ряда видов, сходно реагирующих на среду. Хотя идеи Глизона и были опубликованы в известных изданиях, они не были поддержаны современниками, а сам он вскоре отошел от экологии и долгие годы весьма успешно трудился на поприще систематики растений.
Ситуация, сложившаяся в 20-30-е годы в российской фитоценологии, чем-то напоминала ту, что наблюдалась в США. Доминирующими также были представления о сообществах (фитоценозах) как о дискретных целостных системах с детерминированным развитием. Такие идеи активно развивались, в частности, в трудах бесспорного лидера отечественной фитоценологии В.Н. Сукачева.
Был в России и свой очень яркий «еретик» -Л.Г. Раменский. В 1924 г. в Воронеже он опубликовал ставшую классической статью «Основные закономерности растительного покрова» [31], в которой четко сформулировал тезисы принципиально нового подхода. Растительный покров для Раменского — это не совокупность сообществ, а «многими факторами обусловленная текучая непрерывность, сложно отзывающаяся на смену условий в пространстве и во времени» [31, с. 13]. Всем видам присуща экологическая индивидуальность, то есть свое место на градиенте зависимости обилия от условий, меняющихся в пространстве и времени. Раменский выражал решительное несогласие с доминирующими представлениями о расчленении растительности на неподвижные единицы (ассоциации, сообщества), подчеркивая, что «все течет, не считаясь ни с какими условными границами … устойчивы же не группировки, а только законы сочетаемости растений, они и подлежат изучению» [31, с. 20, выделено Раменским].

Северный сосновый лес после пожара и начало восстановления
В истории фитоценологии суть расхождений во взглядах Сукачева и Раменского (или Клементса и Глизона) сводят к антагонизму между «органицизмом и континуализмом», то есть представлениями о растительном покрове как о наборе четко различающихся целостных сообществ или как о непрерывных переходах, связанных с различиями в зависимости отдельных видов от тех или иных факторов среды. Маурер [32], обсуждая разницу между позициями Клементса и Глизона, делает основной упор на детерминизм в первом случае и стохастизм во втором. Указанные противопоставления справедливы, но еще важнее то, что фактически это было противостояние интегративного (холистического) и редукционистского подходов в экологии.
И если сторонники интегратизма (холизма) основной своей задачей ставили изучение неких целостных систем, которые сначала нужно было еще выделить, договорившись о принимаемых критериях, то редукционисты, будучи менее отягощены умозрительными представлениями, пытались непосредственно обратиться к более элементарным (а главное — более реальным) объектам — локальным популяциям разных видов. Однако отношение к редукционизму биологов вообще и экологов в частности всегда было довольно настороженным, и неудивительно, что призывы Раменского изучать не сообщества, а закономерности пространственно-временного распределения видов, не нашли особенного отклика среди современников.
В отличие от Глизона Раменский не оставил экологию, а продолжая изучать проблему экологической индивидуальности, в конце 30-х годов [33] выдвинул идею о существовании среди растений трех жизненных типов. Виоленты (силовики) очень эффективно захватывают жизненно важные ресурсы и держат их под контролем, патиенты (терпеливцы) могут произрастать в условиях постоянной нехватки ресурса (например, влаги или света), а эксплеренты (заполняющие), характеризуясь высокой скоростью роста, способны быстро занять освободившееся пространство, но не могут его долго удерживать, поскольку вытесняются более сильными конкурентами.
Представления Раменского о трех жизненных типах растений были известны отечественным фитоценологам, но не получали особого распространения вплоть до 70-х годов, когда фактически были переоткрыты английским экологом Ф. Граймом [34], предложившим различать у растений три жизненных стратегии, в зависимости от возможности существовать при разной комбинации стресса, под которым понималась прежде всего нехватка ресурса (света, влаги, элементов минерального питания и т.п.), и нарушений — неизбирательное поедание животными-фитофагами или вытаптывание. Стратегия конкурентов (виолентов, по Раменскому), позволяющая эффективно захватывать и удерживать ресурсы, может проявиться только в условиях обилия ресурса (отсутствие стресса) и слабого нарушения. Стресс-толеранты (патиенты) способны существовать при нехватке ресурса, но не выносят сильного нарушения, а рудералы (эксплеренты) требуют обилия ресурсов, но могут противостоять довольно сильным нарушениям. Очень важным в концепции Грайма было то, что выделенные стратегии рассматривались как альтернативные: усиление признаков, свойственных одной из них, автоматически ведет к ослаблению признаков, свойственных двум другим.
Система Грайма оказалась востребованной потому, что предложена была в то время, когда эволюционная идея стала активно проникать в экологию, в частности, большой популярностью пользовалось введенное несколько раньше Макартуром и Уилсоном представление об г- и К-отборе. Суть последнего сводилась к тому, что более или менее стабильное существование популяций в природе может у разных организмов достигаться по-разному: или за счет большого количества производимых потомков, которые в дальнейшем подвержены очень высокой смертности (r-отбор), или за счет небольшого количества потомков, но лучше защищенных и, соответственно, характеризующихся невысокой смертностью (К-отбор).
Идея об альтернативных способах поддержания организмами своего существования из поколения в поколение для биологов, в общем, была очевидна уже очень давно. Еще в 60-х годах XIX в. ее четко сформулировал (причем в экономических терминах) Г. Спенсер [6]. Но только начиная с работы Макартура и Уилсона эта идея стала практически использоваться для упорядочивания данных по большим совокупностям разных видов. В английском научном языке распространился даже новый термин «trade-off» (трейдофф), обозначающий отрицательную связь между двумя характеристиками организма (например, скоростью соматического роста и интенсивностью размножения), возникающую за счет перераспределения одних и тех же ресурсов на разные цели. Особо много внимания «трейдоффам» стали уделять в 80-90-е годы в связи с бурным развитием исследований по эволюции жизненного цикла, под которым фактически принимали определенный способ выживания, свойственный какому-либо виду или группе видов.
Среди рассматриваемых характеристик жизненного цикла чаще всего фигурируют те, что в конечном итоге сказываются на скорости популяционного роста — возрасте достижения половозрелости, числе продуцируемых потомков, скорости соматического роста, средней массе тела и др. Различные оптимизационные модели, примененные в этих исследованиях, исходили из того, что средняя скорость популяционного роста (г) должна быть неотрицательной величиной [35, 36].
Представления о «трейдоффах» легли в основу ряда гипотез, объясняющих механизмы конкурентного вытеснения или сосуществования видов. Так, моделями, подтвержденными некоторыми экспериментальными данными, было показано [37], что разные штаммы бактерий, конкурирующих за общий ресурс и воздействующих друг на друга специальными ингибиторами роста, сосуществуют только благодаря отрицательной зависимости между способностью вырабатывать токсин (ингибитор роста другого штамма) и скоростью собственного популяционного роста. Образование токсина, а также средств защиты от него требует определенных ресурсов, которые могут быть направлены на рост и деление клеток. Именно поэтому возможны разные стратегии выживания, позволяющие конкурентам жить бок о бок. Аналогичным образом, согласно некоторым моделям, сосуществование многих видов деревьев в тропическом лесу возможно благодаря обратной связи между средней продолжительностью жизни разных видов и средней годовой продукцией семян
Таким образом, было показано, что механизмы формирования сообществ в значительной мере могут быть поняты на пути типичного редукционистского объяснения, через механизм сосуществования видов. Важно и признание того, что любой ныне существующий в природе вид тем или иным способом хорошо приспособлен к окружающей среде. Данный тезис кажется абсолютно очевидным, но он служит отправной точкой при изучении соотношения затрат на совершенствование какой-либо функции и получаемых от этого выгод. «Баланс» подводится в ходе эволюции самим естественным отбором, а итоговым показателем оказывается средняя скорость популяционного роста, которая должна сохранять неотрицательные значения.
Современность: алгоритмы объяснения и макроскопический подход.
К концу XX в. экология стала способна не только описывать (в том числе количественно) исследуемые структуры и процессы, но и объяснять их, причем объяснение касалось как непосредственных механизмов, лежащих в основе тех или иных явлений, так и возможностей их эволюционного происхождения. От описания статики экологи перешли к анализу динамики, в том числе к тем скрытым процессам, которые поддерживают стационарное состояние популяций, экосистем и биосферы.
Очевидно, однако, что успехи экологии как объясняющей науки не основывались на одной обобщающей теории, которая бы охватывала и связывала в единое целое все разнообразие изучаемых структур и процессов.Такой теории нет, а если бы она появилась (например, в рамках общей теории систем и термодинамики неравновесных процессов), то вряд ли произвела революцию в экологии. Специалистам, посвятившим себя этой науке, как правило, неинтересны общие вопросы, подразумевающие столь же общие ответы. Гораздо большую привлекательность для них представляет распутывание конкретных ситуаций, конечно, если при этом не требуется все время сочинять ad hoc гипотезы, а результаты анализа выстраиваются в цельную непротиворечивую картину.
Но что действительно постепенно выработалось в экологии, так это некий набор методологий, позволяющих анализировать цепочки причинно-следственных отношений и вскрывать не всегда очевидные механизмы взаимодействий организмов как между собой, так и с неживыми компонентами среды. В основе этих методологий лежат базовые понятия современного естествознания: законы сохранения, принципы термодинамики, закономерности структурно-функциональной организации живых организмов (при особом внимании к процессам обеспечения их энергией и нужными химическими элементами), а также некоторые более специфические для экологии правила.
К последним относится, например, закон экспоненциального роста, согласно которому численность любой популяции растет (или убывает) экспоненциально (то есть с постоянной удельной скоростью г), «пока окружающая среда по отношению к каждой особи в этой популяции остается неизменной» [38]. Признание экспоненциального роста как некоего универсального принципа является неотъемлемой частью дарвиновской концепции происхождения видов, и отсюда естественно возникает связь популяционной экологии с современным дарвинизмом. Впрочем, не меньшее значение имело и чисто эмпирическое правило, согласно которому экспоненциальный рост ограничен во времени уже хотя бы потому, что рано или поздно обязательно возникнет нехватка ресурсов.
Очень важная особенность современного этапа развития экологии — это растущее осознание того, что любая выявленная упорядоченность структуры (паттерн) строго привязана к определенному масштабу пространства-времени и, соответственно, может существенным образом изменяться или вообще никак не обнаруживаться при использовании иного масштаба. По отношению к живому покрову Земли подавляющее большинство исследований в экологии оставались локальными, если не сказать точечными. Появление аэрофотосъемки, а позднее и методов обследования с помощью космических аппаратов произвело настоящую революцию в получении интегрированных оценок многих экосистемных процессов (например, первичной продукции океана, рассчитанной на основе дистанционного определения содержания хлорофилла по всей акватории). Что касается популяционной экологии, имеющей дело с совокупностями организмов, то здесь переход к другому масштабу пространства и времени происходил за счет накопления и статистической обработки данных, полученных многими исследователями на разных видах во многих местах, разбросанных по огромной территории.
Анализ таких больших наборов данных позволяет выявить определенные закономерности, никак не проявляющиеся в локально исследуемых популяциях и сообществах. Так, для всех млекопитающих Северной Америки обнаружена четкая картина распределения числа видов по отдельным размерным группам (массе тела взрослых особей) [39]. Также на основании множества данных из разных опубликованных источников была построена зависимость плотности популяций разных видов млекопитающих в естественных местах обитания от массы тела взрослых особей. Выяснилось, что по мере увеличения массы тела (W) плотность популяции убывает пропорционально массе в степени -0.75 [40]. Поскольку обмен животных пропорционален массе тела в степени +0.75, то из комбинации обеих зависимостей следовало, что потребление энергии популяциями млекопитающих разного размера оказывается одним и тем же, не зависящим от массы тела. Данный удивительный вывод, получивший название «правила энергетической эквивалентности«, подтвержден теперь и для растений [41].
Эти и другие примеры закономерностей, выявляемых только для больших совокупностей данных, относящихся к множеству видов и огромным территориям, составляет предмет специального направления экологии, получившего название «макроэкология» [39]. В развитие макроэкологии определенный вклад внесли и отечественные исследователи. Так, А.И. Азовскому [42] на примере морского прибрежного бентоса удалось показать, как увеличивается число видов из разных размерных групп при расширении обследуемой территории. Выяснилось, что чем мельче организмы, тем выше для них локальное разнообразие, но тем медленнее оно возрастает по мере увеличения охватываемого исследованиями пространства, вплоть до включающего все известное видовое разнообразие данной группы.
Другой важный результат получен Л.В. Полищуком [43], обобщившим данные по жизненным характеристикам млекопитающих России и прилегающих стран (территории бывшего СССР) и показавшим, что степень их уязвимости (вероятности вымирания, оцененной по факту включения в «Красную книгу СССР») может быть надежно предсказана по среднему числу потомков, произведенных самкой за год.
Таким образом, в современной экологии мы видим радикальное расширение горизонта исследований при сохранении основного его теоретического базиса. Признаков смены парадигм в строгом смысле этого слова пока нет, но это не есть признак застоя. Наши знания о том, как устроен живой покров Земли, становятся все более фундаментальными.
Работа выполнена при поддержке гранта РФФИ № 04-04-49623.
ЛИТЕРАТУРА
- Алимов Л.Ф. Об экологии всерьез // Вестник РАН. № 12.
- Заварзин Г.А. Лекции по природоведческой микробиологии. М.: Наука, 2003.
- Popper К. Evolution and the tree of knowledge / In: Popper K. Objective knowledge. An evolutionary approach. Oxford: Clarendon Press, 1979.
- The changing place of theory in 20th century ecology: from universal laws to array of methodologies // Oikos. 2001. V. 92. № 2.
- Krebs C. Ecology: The experimental analysis of distribution and abundance. N.Y.: Harper and Row,
- Бигон М., Харпер Дж., Таунсенд К. Экология: особи, популяции и сообщества. М.: Мир, 1989.
- Foucault М. Les mots et les choses. Une archeologie des sciences humaines. Paris: Gallimard,
- Nicolson M. Alexander von Humboldt, Humboldtian science and the origins of the study of vegetation // History of Science. V. № 68.
- Nicolson M. National styles, divergent classifications: a comparative case study from the history of French and American plant ecology // Knowledge and Society: Studies in the Sociology of Science Past and Present. V. 8.
- Миркин Б.М. Очевиден ли вклад дарвинизма в экологию? // Журн. общ. биологии. 2003. № 5.
- Гиляров A.M. Дарвинизм как средство ограничения экологического плюрализма // Журн. общ. биологии. 2003. № 5.
- Gould S.J., Lewontin R.C. The spandrels of San Marco and the Panglossian paradigm: a critique of adaptionist programme // Proc. R. Soc. London. V. 205.
- Gould S.J. The structure of evolutionary theory. Cambridge, Mass.: Belknap Press,
- Simard S.W., Perry DA., Jones M.D., et al. Net transfer of carbon between ectomycorrhizal tree species in the field // Nature. V. 388.
- Гиляров A.M. Мифологическое в экологии // Природа. 1992. №2.
- Spencer H. First principles. N.Y.: Thomas Crowell,
- Spencer H. The principles of biology. London: Williams andNorgate,
- E. Modeling Nature: Episodes in the History of Population Ecology. Chicago: Chicago University Press, 1985.
- Lotka A.J. Elements of physical biology. Baltimore: Williams and Wilkins,
- Тахтаджян A. Principia tectologica. Принципы организации и трансформации сложных систем: эволюционный подход. СПб.: Изд-во СПХФА, 2001.
- Красилов В.А. Охрана природы: принципы, проблемы, приоритеты. М.: Институт охраны природы и заповедного дела, 1992.
- Миркин Б.М. Теоретические основы современной фитоценологии. М: Наука, 1985.
- Новиков Г.А. Очерк истории экологии животных. Л.: Наука, 1980.
- Elton С. Animal ecology and evolution. Oxford: Clarendon Press,
- Charnov E. Life history invariants: Some explorations of symmetry in evolutionary ecology. Oxford: Oxford Press.,1993.
- Gause G.F. The struggle for existence. Baltimore: Williams and Wilkins,
- Peters R.H. A critique for ecology. Cambridge, N.Y.: Cambridge University Press, 1991.
- Галл Я.М. Популяционная экология и эволюционная теория: историко-методологические проблемы // Экология и эволюционная теория. Под ред. Я.М. Галла. Л.: Наука, 1984.
- Tilman D. Resource competition and community structure. Princeton (N.J.): Princeton Univer. Press, 1982.
Вестник РАН, 2005. Т.75. №3. С.214-223.
Рекомендуем прочесть
 «Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве
«Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве Основные концепции и методы восстановления природных лесов Восточной Европы
Основные концепции и методы восстановления природных лесов Восточной Европы Курносые древесные лягушки защищаются от хищников слуховыми иллюзиями
Курносые древесные лягушки защищаются от хищников слуховыми иллюзиями Птицы и муравьи не мешают друг другу есть гусениц
Птицы и муравьи не мешают друг другу есть гусениц Плейстоценовый перепромысел: роль человека в массовых вымираниях мегафауны
Плейстоценовый перепромысел: роль человека в массовых вымираниях мегафауны Роль крымских татар в кампаниях Богдана Хмельницкого
Роль крымских татар в кампаниях Богдана Хмельницкого Как сыграл «фактор Пиночета» на президентских выборах в Чили
Как сыграл «фактор Пиночета» на президентских выборах в Чили История из американской жизни
История из американской жизни
