Популяция уменьшилась на 84,3-94,7 % между 1980 и 2013 годами, а ареал сократился на 5000 км. Количественные данные полицейских рейдов свидетельствуют о необузданном незаконном отлове вида на его Восточноазиатском миграционном маршруте в Китае. Популяционная модель, предполагающая начальный уровень добычи в размере 2% популяции и ежегодное увеличение на 0,2% в течение периода мониторинга, производит траекторию популяции, соответствующую наблюдаемому снижению. Мы предлагаем, что в снижение численности внес большой вклад отлов сетями, потому что добыча дубровника и других певчих птиц увеличилась в результате экономического роста и процветания в Восточной Азии. Величина и скорость упадка этого вида беспрецедентны среди птиц с сопоставимым размером ареала, за исключением странствующего голубя (Ectopistes migratorius), который исчез в 1914 г. из-за охоты промышленного масштаба. Наши результаты демонстрируют срочную необходимость в совершенствовании мониторинга популяций дубровника, до недавнего времени бывшего обычным и широко распространенным, и уточнении масштаба его добычи повсюду в Восточной Азии.
Темпы исчезновения видов, вызванного человеком, в настоящее время приблизительно в 100-1000 раз превышает естественный темп и, по прогнозам, нарастает (Pimm et al. 1995). Риск полного исчезновения варьирует среди разных видов. Специализированные и редкие виды с маленькими ареалами, населяющие острова или имеющие низкую плотность населения, наиболее уязвимы (Cardillo et al. 2005; Kotiaho et al. 2005), потому что они наиболее чувствительны к природным бедствиям, болезням и беспокойству со стороны людей (McKinney 1997). Случаи сильного снижения численности видов, прежде чрезвычайно многочисленных и с очень большими ареалами вплоть до почти полного исчезновения, по вине человека — редки. Традиционно природоохранная биология сфокусирована на редких видах, поскольку для них риск их вымирания выше (Lindenmayer et al. 2011). В противоположность, обычные виды принято считать находящимися в безопасности. Однако свежие данные свидетельствуют, что у обычных видов потери, и весьма значительные, могут быть недооцененной природоохранной проблемой (Inger et al.2015). Обычно главными факторами, обусловливающими потери биоразнообразия, считаются человеческие действия, связанные с изменениями в использования земли, такие как сельское хозяйство и лесоводство, или изменение климата (Vitousek et al. 1997). Прямое преследование видов играет менее важную роль в промышленно развитых странах, но в развивающемся мире это все еще постоянная угроза (Rosser, Mainka 2002; Warkentin et al. 2009). Особенно в Восточной Азии, где потребление дериватов живой природы в медицинских, ритуальных и пищевых целях внедрено в культуру и широко распространено (Sodhi et al. 2004; Nijman 2010), охота и переэксплуатация являются главными причинами исчезновения видов (Yiming, Wilcove 2005). Воздействие преследования хорошо изучено для загадочных и заметных видов в Восточной Азии (например, Gao, Clark 2014; Liu, Weng 2014), но об эффекте крупномасштабного отлова и потребления многочисленных и широко распространенных наземных видов известно немного (Sodhi et al. 2004; Yong et al. 2015). В большинстве работ эффект отлова изучен только на местном уровне (Liang et al. 2013). Кроме того, во многих странах Восточной Азии наблюдаются диспропорции между человеческим населением и экономическим ростом. Взаимодействия между ростом, процветанием и традиционным укладом потребления живой природы и отдельных видов изучены еще слабо.
Во время миграций и в местах зимовки на ночевку дубровники собираются в огромные стаи. В Китае и в меньшей степени в Таиланде и Камбодже в этот период на ночевках птиц традиционно отлавливают паутинными сетями для употребления в пищу (Glutz von Blotzheim, Bauer 1997). В Китае в 1997 г. вслед за начальным снижением численности, торговля видом как пищевым продуктом была запрещена, однако незаконная торговля на всем миграционном маршруте в Китае сохранилась, и для дубровника и других певчих птиц существует черный рынок. S. Chan (2004) оценил общее количество дубровников, отловленных в китайской провинции Гуандун в 2001 г. в 1 миллион особей. Особе беспокойство вызвало то, что в ближайшие годы из-за возрастающего спроса в Китае, вызванного ростом населения и экономики, масштаб истощительного отлова, скорее всего, увеличится.
Мы рассмотрели опубликованные и неопубликованные данные с 237 участков в разных местах гнездового и зимовочного ареала за 1980-2013 гг. и использовали неопубликованные данные мониторинга, чтобы установить глобальный тренд популяции и географические точки снижения численности. Мы оценили масштаб незаконной торговли видом, основанный на количественной информации о конфискованных птицах в результате полицейских рейдов в Китае. Сравнив прогнозируемые тренды при разных сценариях популяционной модели, мы оценили, насколько вероятный масштаб добычи и потенциальные альтернативные причины соответствуют наблюдаемому снижению численности.
Методы
Обзор литературы
Мы провели поиск изданной и неопубликованной информации относительно локальных трендов дубровника за 1980-2013 гг. В дополнение к стандартному поиску в интернете по сервисам Google Scholar и Web of Science, мы включили в наш поиск web-библиотеки на русском и китайском языках, особенно российскую базу данных http://www.elibrary.ru. Красные книги из всех областей и краев Российской Федерации (доступные онлайн на http://oopt.aari.ru/bio/8672) также были проанализированы. Дополнительно были просмотрены интернет-ресурсы типа вебсайтов и блогов по регионам, входящим в ареал вида. Российским, казахстанским и монгольским орнитологам были разосланы по электронной почте запросы о размерах популяций и плотностях населения дубровника за длительные периоды, в расчете на неопубликованные данные по их регионам. Мы также использовали информацию о природоохранном статусе вида, доступную на
форуме Birdlife International по глобально угрожаемым видам, а также просмотрели онлайн-ресурсы для вида на английском языке по его латинскому названию (включая непризнанный синоним
Ocyris aureola, который широко используется в России) и по названиям в языках других стран. При поиске вводилась комбинация названия вида со словами «динамика, изменение, популяция, численность, сокращение, стабильность, рост.
Где это было возможно, мы извлекали оценки обилия и плотности населения вместе с местоположением и временем оценки. Часто приходилось довольствоваться только поверхностными свидетельствами, предполагающими изменения в популяции. Описательная информация (например, изменения от обычного, многочисленного, или очень обильного к более редкому, очень редкому, почти исчезнувшему, устойчивому или исчезнувшему) отмечалась с указанием года исследования. В общей сложности доступной оказалась информация относительно изменений в популяциях для 237 участков (225 участков в пределах гнездового ареала и 12 — в местах зимовок), которые с достаточной детальностью можно идентифицировать на картах и определить для них географические координаты. Представительная информация собрана для европейской части России, южной Сибири и российского Дальнего Востока. Существование стабильных популяций в регионах, не охваченных нашим анализом, мы сочли маловероятным, поскольку во всем зимовочном ареале вида отмечено снижение численности.
Отрицательные тренды в популяциях упоминались в публикациях чаще, нежели положительные или стабильные тренды, ввиду стремления исследователей привлечь внимание к необходимости охраны. Этот уклон, возможно, вел к переоценке общего масштаба снижения. Поскольку мы анализировали не только источники, непосредственно связанные с дубровником, но и оценки численности по другим видам птиц, а в некоторых регионах — для всех видов, включая сведения о локальных сообществах птиц, регионов, где численность дубровника была стабильной или увеличилась с 1980 по 2013 гг., вряд ли осталось много.
Всю доступную информацию об изменениях численности мы классифицировали по следующим критериям. Если количественные данные (индексы учетов или оценки плотности) были доступны по крайней мере за 2 года, отстоявших не менее 10 лет друг от друга, мы вычисляли различие между этими оценками и изменение численности > 90% классифицировали как очень сильное снижение, уменьшение > 50% — как сильное и > 30% — как умеренное.
Если информация была лишь поверхностной (например, очень обильный в период 1 и редкий в период 2), мы давали качественные категории степени снижения.
Компиляция данных по отлову
После того, как добыча вида в Китае в 1997 г. была признана незаконной, полиция начала проверять рынки, поезда и автотранспорт и искать добытых дубровников. Успешные рейды часто освещались в прессе и на интернет-сайтах. Для получения обзора о величине и географических районах незаконного отлова в Китае мы компилировали сведения из интернета и СМИ о добытых и конфискованных птицах за 2000-2014 гг. Конечно, учету поддаются только те случаи незаконной добычи, которые были выявлены, сообщены в полицию и впоследствии опубликованы в СМИ. Поэтому количество конфискованных птиц занижает величину добычи, а усилия полиции, отчетность, и интерес СМИ изменялись по годам, с общим числом отловленных птиц этот показатель может быть не связан.
Анализ данных мониторинга и исследований
Для количественной оценки изменений в популяции и выявления временных диапазонов, мы проанализировали данные многолетнего мониторинга и повторных учетов на 9 участках по всему ареалу (от восточной Финляндии до Амурской области на российском Дальнем Востоке) (рис. 1). В Финляндии и в Европейской России для оценки численности использовалось картирование территории. В Финляндии, в одной из областей, и на южном берегу озера Байкал,чтобы избежать проблем с «псевдодублированием», мы объединили данные учетов в 3-х и 4-х участках, расположенных друг около друга. На сибирских и дальневосточных участках учеты проводились на пешеходных трансектах с фиксированной шириной полосы, плотность населения рассчитывали как количество поющих самцов на квадратный километр. Эти трансекты охватывали все типы биотопов, учеты повторялись в одно и то же время года (с конца мая до середины июня, в период пика гнездовой активности вида). На двух участках в Финляндии и на одном около Москвы территориальные пары дубровника регистрировали ежегодно с начала 1980-х, после исчезновения дубровников наблюдения на этих участках продолжали спустя 2-5 лет после года последней регистрации вида. На всех остальных участках учеты в течение гнездового периода проводились не ежегодно.
Путем моделирования обилия как функции от года были получены ежегодные индексы размера популяции. Мы использовали обобщенные линейные смешанные модели (GLMMs) со структурой ошибки Пуассона и логарифмическую связь в программном пакете lme4 R (версия 3.0.2). «Сырые» подсчеты поющих самцов или пар использовались как зависимые переменные для участков, где проводилось картирование территории. Все данные по плотности (количество поющих самцов на квадратный километр), полученные путем пересчета данных учетов на линейных трасектах, были преобразованы в численность путем их умножения на различные значения площади.
Для проверки чувствительности итоговых трендов к используемому размеру площади мы использовали 3 различных размера площади: 1, 10 и 100 км². Участок рассматривался как произвольный фактор, а год — как фиксированный фактор. Параметры оценок для года были затем стандартизированы и использовались в качестве индекса размера популяции в данном году по всем участкам. Для контроля сверхдисперсии мы рассматривали случайный эффект дополнительного уровня наблюдения.
Пригодность модели была оценена крайним R² по Nakagawa и Schielzeth (2013), которое интерпретируется как разница, объясняемая фиксированными факторами модели.
Для вычисления популяционного тренда для ареала в целом использовались обобщенная совокупная модель (GAM) с Гауссовской структурной ошибкой и кубический сплайн сглаженного значения k = 3 (в R программном пакете mgcv) (Wood 2006), чтобы сгладить ряд временных ежегодных индексов. Индекс популяции был смоделирован как функция года. Ежегодное изменение обилия в процентах и оценка общего снижения численности популяции за 34-летний период были, в конечном счете, рассчитаны по прогнозируемым модельным значениям.
Определение времени локального исчезновения
Для 86 участков (38,2%), выделенных по литературным данным, в источниках был указан приблизительный год локального исчезновения дубровников. Мы ожидали, что направление исчезновения вида будет с запада на восток ареала, согласно его биогеографическому происхождению в Восточной Палеарктике. Мы использовали ту же модель GAM, описанную выше, чтобы связать время исчезновения участков с известными годами исчезновения, от западных и центральных частей ареала по широте и долготе. Поскольку на время исчезновения, возможно, также влиял биотоп, мы включали растительный покров как замещающую переменную для биотопа как установленный фактор в моделях. Значения для растительного покрова были взяты из Globcover 2009, глобальная классификация растительного покрова — из Olson et al. 2001. Для коррекции возможного снижения численности дубровников в районах плотного человеческого населения, во всех моделях плотность человеческого населения была принята за несглаженную ковариату. Значения с 2000 года были использованы и извлечены из глобальной растровой базы данных с разрешением 0,5° (CIESIN 2000). Модели, содержащие все возможные комбинации переменных, были сравнены путем использования информационного критерия Akaike для маленьких размеров выборки (AICc). Модели с ∆AICc <2 от модели с наиболее низким AICc считали одинаково информативными и имеющими существенную поддержку.
Исследование потенциальных причин популяционного коллапса
Снижение численности может быть вызвано различными факторами, среди которых наиболее заметные — потеря среды обитания или увеличение смертности из-за преследования или загрязнения среды. Поскольку прямого свидетельства, что крах популяции дубровника вызван именно добычей в Китае, мы вычислили популяционную траекторию вида для 4-х различных сценариев, которые отражали вероятные крупномасштабные изменения окружающей среды. Чтобы исследовать траекторию популяции для каждого сценария и проверить, совпадало ли наблюдаемое снижение численности популяции с реалистичными ожиданиями уровня добычи, мы проанализировали жизнеспособность популяции. Сначала мы провели базовое моделирование, которое учитывало особенности жизненного цикла и информацию относительно репродуктивного темпа и смертности в стабильных популяциях. Значения необходимых параметров, кроме оценки смертности первогодков и взрослых птиц, были получены из публикаций о дубровнике. В качестве оценки смертности была использована такая же величина для экологически близкого мигрирующего вида — камышовой овсянки (Emberiza schoeniclus), полученная путем надежных исследований в Европе.
Мы считали, что инбредная депрессия отсутствует, соотношение полов взрослых птиц 1:1, и нет катастрофы популяции (внезапной потери индивидуумов). Затем мы построили контрастную базовую модель с 4 альтернативными сценариями при разных вероятных причинах снижения численности. Первый сценарий описывал добычу дубровников и содержал функцию, моделирующую регулярный отлов. Мы выдвинули гипотезу, что отлов был широко распространен еще до начала снижения численности популяции и увеличился недавно из-за роста потребностей людей. Были приняты три варианта стартовой добычи: 1%, 2% и 5% популяции, при последующем ежегодном увеличении на 0,1%, 0,2% и 0,5% отловленных особей до последнего года моделирования.
Второй сценарий описывает изменения среды обитания: крах популяции связан с нарастающим темпом потери местообитаний. При этом сценарии максимальная плотность популяции, которая может длительно поддерживаться саморегуляцией (представленная доступной средой обитания), уменьшалась ежегодно на 10%, 20%, и 50%.
В третьем сценарии мы предполагали, что смертность росла постепенно из-за загрязнения, что приводило к болезням или отравлению, вплоть до смертельного. Мы моделировали этот сценарий, увеличивая смертность взрослых птиц и первогодков на 5%, 10% и 15%, в соответствии с наблюдаемыми различиями в выживании в стабильных, увеличивающихся или сокращающихся популяциях большого количества видов певчих птиц (Siriwardena et al. 1998).
При четвертом сценарии потеря среды обитания и ее загрязнение произошли одновременно, и мы объединили эффекты сценариев среды обитания и загрязнения, сокращая максимальную плотность популяции, которая может длительно поддерживаться саморегуляцией, и увеличивая смертность при моделировании.
Моделирование популяционной динамики выполнено в программе Vortex 10.0. Это программа детерминированного моделирования, которая может учитывать эффекты демографических, экологических и генетических стохастических событий в популяции в живой природе (Lacy 1993). Моделирование выполнено для гипотетического размера популяции в 100 птиц и проводилось 100 раз для более чем 34 лет, соответствующих периоду, охваченные данными мониторинга.
Результаты
С 1980 по 2013 гг. в 82,5% мест гнездования и зимовки было зарегистрировано очень сильное снижение, в 13.8% — сильное снижение (рис. 1). Никаких свидетельств стабильности популяций после 2000 г. в пределах всего гнездового и зимовочного ареала найдено не было. Очень сильное снижение было также отмечено по всему зимовочному ареалу (например, в Непале и Бангладеше, начиная с года 2000 г., и в Камбодже примерно с 2005 г.) (Рис. 1).
Популяционный тренд для всего ареала, основанный на повторных учетах и ежегодных данных мониторинга на 9 участках, был чувствителен к области, которая использовалась при преобразовании плотностей населения на линейных трансектах в значения обилия (рис. 2).
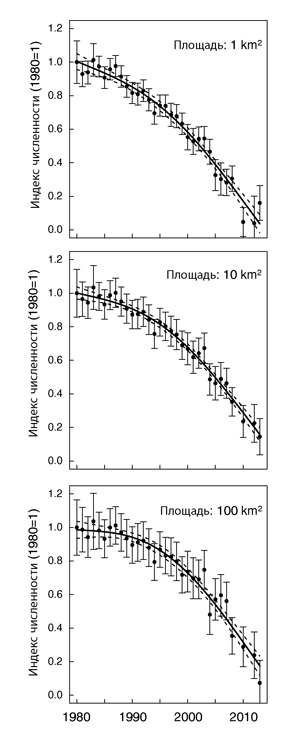
Рисунок 2. Популяционный тренд дубровника для ареала в целом в 1980-2013 гг., рассчитанный по данным мониторинга на 9 участках в разных частях ареала. Смоделированные ежегодные индексы обилия из обобщенной линейной смешанной модели показывают (стандартизированы до 1980 = 1): сплошная линия — сглаженный тренд за полный период; пунктирная линия — предсказанные значения от обобщенной добавочной модели; вертикальные линии — параметрический 95 % доверительный интервал. Три кривые альтернативных трендов сделаны по 3 -м моделям, для иллюстрации чувствительности оценки тренда к изменяющейся по размеру площади, когда преобразование плотности были преобразованы в значения обилия.
Преобразование, базировавшееся на размере площади исследований 100 га (1 км²), привело к предсказанию снижения численности на 94,7% от 1980 г. к 2013 г. (GLMM [ежегодные индексы]: крайнее R² = 0.847; GAM [cглаженная тенденция]: прил. R² = 0.97, P <0.001); снижение, основанное на площади 10 км² было оценено как 84,3% (GLMM: крайнее R ² = 0.744; GAM: прил. R² = 0.98, P <0.001) и как 87,2% для площади 100 км² (GLMM: крайнее R² = 0.744; GAM: прил. R² = 0.95, P <0.001). Однако общий тренд в разных моделях был сходен: примерно до 1987 г. популяция была, скорее устойчивой, после чего началось сильное снижение (Рис. 2). К 2012 г. вид фактически исчез в Европейской России, западной и центральной Сибири и Казахстане: в районах, где они были очень обильными до середины 1990-х, наблюдались только отдельные птицы.
GAM, который связал год исчезновения с широтой и долготой, как переменными, получил самую большую поддержку, и предложенная модель оказалась хорошей.
Модели, содержащие растительный покров, варьирующий в пределах 2 ∆AICc единиц, но без переменной долготы, дали большое увеличение ∆AICc, предлагая, что оно было наиболее влиятельной переменной. Участок лучшей модели (поддерживающий человеческое население фиксированным значением при варьирующем среднем) предположил быстро прогрессирующее исчезновение с северо-запада к юго-востоку гнездового ареала (рис. 3). Судя по всему, вид отступает в восточном направлении примерно на 5000 км менее чем за 25 лет, что эквивалентно отступлению гнездового ареала на 200 км в год.
Масштабный отлов и торговля на всем миграционном маршруте в Китае продолжались и после того, как в 1997 г. они были запрещены. Большинство сообщений о случаев незаконной добычи и потребления вида поступило из южнокитайской провинции Гуандун (рис. 1). Ежегодные публичные отчеты об общем количестве конфискованных птиц показывали цифры от 400 (в 2006 г.) до 119000 особей (в 2001 г.) (среднее 25140 [SD 30 228], n = 14 лет) (рис. 1). Лишь за одну операцию в 2013 г. китайские власти и полиция конфисковали более 2 млн. певчих птиц (не все из них дубровники) (рис. 4), что составляет лишь часть незаконно отловленных птиц. Дубровники были добыты и конфискованы не только в провинции Гуандун, где большинство их употребляется в пищу, но также и на всем китайском миграционном маршруте, до 2500 км от Гуандуна (рис. 1).
Сценарии с разными потенциальными факторами предполагали, что добыча 2% от общей численности в 1980 г. и последующее постепенное увеличение на 0,2% в год до 8,6% в 2013 г. стала бы причиной уменьшения численности, подобной оцененному снижению по данным мониторинга популяции (средний ежегодный темп прироста населения λ = 0.92) (рис. 5).
Уменьшение максимальной плотности популяции, которая может длительно поддерживаться саморегуляцией, моделируемое ухудшением или потерей среды обитания, оказывается при расчете намного менее серьезным, чем наблюдаемое. Увеличение смертности взрослых птиц и первогодков в нашем сценарии среды обитания вело к величине снижения численности, подобной наблюдаемому снижению. Однако, чтобы добиться снижения, сопоставимого с наблюдаемым фактически, необходимо увеличение смертности > 10% (среднее λ = 0.90) (рис. 5). Сценарий, комбинирующий низкую выживаемость и уменьшение максимальной плотности популяции, которая может длительно поддерживаться саморегуляцией, только незначительно отличался от сценария, который учитывал только увеличение смертности и тоже требовал существенного увеличения смертности > 10% (рис. 5).
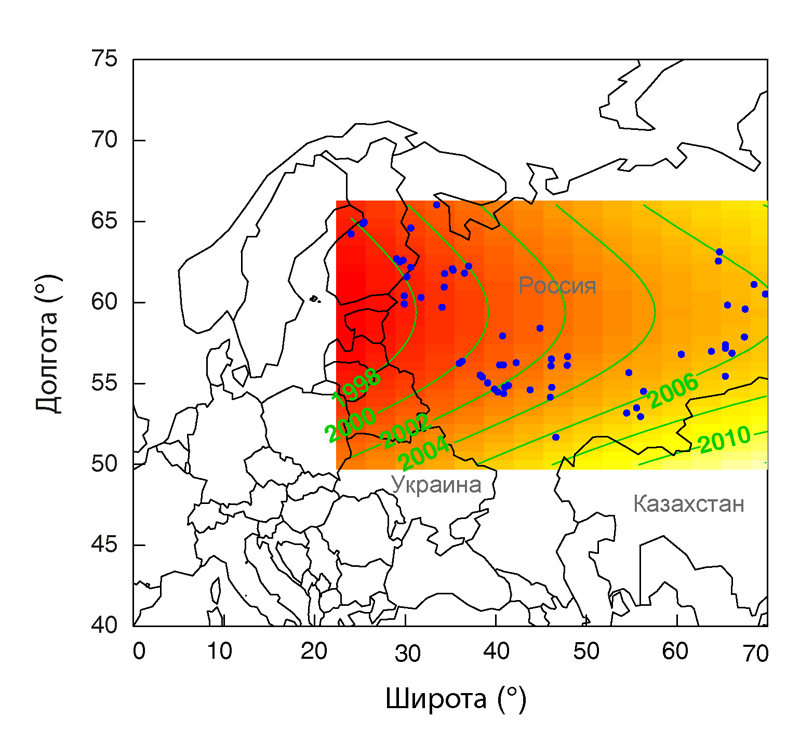
Рисунок 3. Смоделированное время исчезновения дубровника в западных и центральных частях его ареала, как функция широты и долготы (обобщенная совокупная модель). Плотность человеческого населения была включена как ковариата и была фиксированной на участке при переменном среднем (точки — участки, из которых использовались данные относительно года исчезновения для построения модели в параметрической форме). Линии контура отделяют области сходного предсказанного времени исчезновения; более интенсивная штриховка
Обсуждение
Мы выяснили, что мировая популяция ранее очень многочисленного дубровника за последние 25 лет потерпела крах, и что ареал вида сократился на 5000 км в восточном направлении. Наши результаты предполагают, что после официального запрета в 1997 г. отлов этих овсянок в Китае не прекращался, а скорее продолжался, и при этом на очень высоком уровне.
Моделируемые популяционные траектории показали, что наблюдаемого снижения можно добиться при начальной добыче 2% популяции с ежегодным увеличением на 0,2% до финального уровня 8,6% в 2013 г. На протяжении 1980-х размер мировой популяции предположительно составлял 100 миллионов птиц (предположение базировалось на площади ареала и крупномасштабной оценке плотности в 1 поющего самца/га в основных районах гнездования); добыча в 1980 и 2013 гг. должна была бы составлять приблизительно 2 миллиона и 8,6 миллиона птиц соответственно. Эти величины представляются реалистичными, поскольку согласуются с сообщениями о количестве конфискованных птиц.
Следовательно, широкомасштабный отлов по миграционному маршруту и, возможно, также в районах зимовки, мог стать причинной или, по крайней мере, внести большой вклад в наблюдаемое снижение численности.
К местам зимовок дубровники со всего гнездового ареала летят вдоль восточного китайского побережья и поэтому должны пройти описанное «бутылочное горлышко» с чрезвычайно высоким прессом отлова. Спрос, торговля и потребление всех объектов живой природы на всем миграционном маршруте дубровников за прошедшее десятилетие коренным образом возросли (Lau et al. 1996). Главная причина этой тенденции — общий рост уровня жизни, отразившийся в массовом росте внутренних продуктов в Китае (Lau et al. 1996). Шестьдесят процентов китайского людского населения потребляет продукты живой природы, а главная группа потребителей — молодые люди с высокими уровнем образования и доходами (Zhang et al. 2008). С 1992 по 1997 гг. более 10000 туристов (главным образом китайских) посетили город Саншуй (провинция Гуандун, Китай) для участия в ежегодном фестивале продовольствия, где было съедено несколько сотен тысяч дубровников (Chan 2004), и их употребление продолжалось после того, добыча была запрещена в 1997 г. Дубровники и другие птицы используются не для удовлетворения основной потребности в пище возрастающего в числе (и главным образом бедного) населения. Скорее, они стали изысканным блюдом для сравнительно богатых людей (Liang et al. 2013).
До 1980-х добыча птиц имела оправданием пропитание, но в настоящее время целые деревни живут отловом разных видов птиц на продажу в большом количестве дилерам, которые ездят по сельской местности (Liang et al. 2013). Цены на пойманных диких животных быстро растут (Lau et al. 1996). Дубровник в настоящее время продается приблизительно по 8-11 долларов за особь, но сообщалось также о местном уровне цен в 30-40 долларов. Возрастающее процветание китайцев означает, что больше людей могут позволить себе покупать объекты живой природы, несмотря на повышающиеся цены (Liang et al. 2013). Кроме того, улучшение инфраструктуры облегчает транспортировку популярных объектов живой природы на большие расстояния (Lau et al. 1996; Sodhi et al. 2004).
Дальнейшее увеличение спроса последовало в результате недавнего открытия границ между Китаем и соседними государствами (например, Вьетнамом, Индонезией, Лаосом), где процветает масштабная торговля живой природой (35 миллионов особей видов, занесенных в список конвенции CITES в 1998-2007 [Lau et al.1996; Nijman 2010]). Сверхэксплуатация и потребление считаются сейчас наиболее важными факторами, приводящими к угрожаемому состоянию и исчезновению видов в Китае; 53,5% видам птиц, находящихся в опасности, угрожает добыча для продовольствия (Yiming, Wilcove 2005). Поэтому мы пришли к заключению, что в быстром крахе глобальной популяции дубровника важную роль сыграла высокая и, возможно, увеличивающаяся даже после ее запрещения, добыча.
Популяционному коллапсу могут способствовать широкомасштабные изменения в землепользовании. После распада Советского Союза в 1991 г. местах гнездования было заброшено более 30 млн. га пахотных земель (Schierhorn et al. 2013) и резко сократилось поголовье домашнего скота (Dubinin et al. 2011). Эти изменения привели к спонтанной растительной сукцессии в бывших пахотных угодьях, сенокосных лугах и пастбищах на обширных территориях (Dubinin et al. 2011). Однако дубровник — вид ранних стадий сукцессии (Glutz von Blotzheim, Bauer 1997), поэтому после 1991 г. пригодность гнездовых биотопов на какое-то время, похоже, увеличилась. Мы не обнаружили начального увеличения численности популяции после 1991 г. Наша популяционная модель предполагает, что уменьшение размера максимальной плотности популяции, которая может длительно поддерживаться саморегуляцией, вряд ли может объяснить популяционный крах, даже если было потеряно 50 % доступных биотопов. Поэтому мы считаем, что вряд ли потеря среды обитания была ключевым фактором, определившим снижение численности.
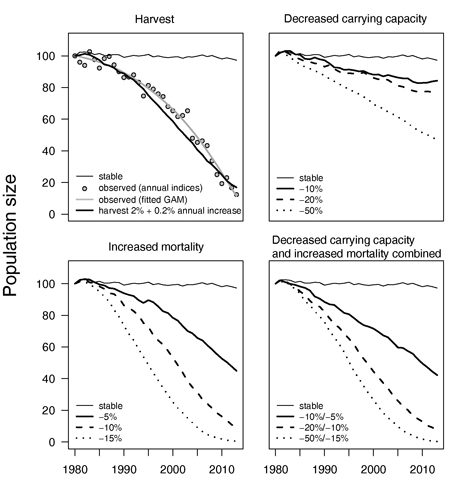
Рисунок 5. Моделируемый размер популяции дубровника более чем за 34 года, базирующийся на 4-х сценариях, влияющих на демографические параметры этой птицы (линии — подсчитанные средние популяционные траектории для 100 воспроизведений популяционной модели, построенные программой Vortex). В квадрате со сценарием добычи соответствующие значения наблюдаемой тенденции и более сглаженная функция обобщенной добавочной модели (GAM) (рис. 2) показаны для сравнения.
Пояснения к рисунку 5. По оси ординат показан размер популяции. Названия сценариев (от верхнего левого по часовой стрелке): добыча; уменьшения размера максимальной плотности популяции, которая может длительно поддерживаться саморегуляцией; комбинация уменьшения размера максимальной плотности популяции, которая может длительно поддерживаться саморегуляцией и возрастания смертности; возрастание смертности. Подписи к графическим фигурам сценария добычи: тонкая черная линия – стабильность; кружок – наблюдаемые значения (ежегодные индексы); жирная серая линия – наблюдаемое (соответствующее модели GAM); жирная черная линия – добыча 2 % + ежегодное увеличение на 0,2 %).
Быстрый крах популяции может также быть вызван болезнями или загрязнением среды. Наша популяционная модель предполагает, что для столь быстрого уменьшения численности, как это реально произошло, необходимо ежегодное увеличение смертности взрослых птиц и первогодков по крайней мере на 10%. Такая смертность могла быть вызвана загрязнением среды, как это было описано для других видов птиц с очень большими популяциями, которые разрушились в течение нескольких лет (например, сипы [Green et al. 2004]). В Китае, где находятся самые важные места миграционных остановок дубровника, использование пестицидов в сельском хозяйстве увеличилось более чем в два раза между 1990 (740000 т) и 2012 (1620000 т [Zhang et al. 2011]). Интенсификация сельского хозяйства также происходила в местах зимовок вида во Вьетнаме и Таиланде, а именно в системах выращивания риса. Пестициды могут затронуть популяции птиц непосредственно (например, через резкую смертность, сублетальный стресс, уменьшение плодовитости [Fry 1995]) или косвенно, через сокращение обилия беспозвоночных, составляющих кормовую базу (например, Hallmann et al. 2014). Однако случаи внезапной смертности, которые убили бы миллионы певчих птиц в местах остановок на миграционном маршруте или в местах зимовок, нам неизвестны.
Наш сценарий с потерей среды обитания показал, что одна только потеря среды обитания не могла привести к наблюдаемому сокращению популяции. Мы не смогли определить, произошло ли увеличение смертности исключительно из-за истощительной добычи, увеличения загрязнения среды или комбинации этих факторов. Однако, масштабы добычи, необходимой для наблюдаемого краха, кажутся реалистичными в свете публично доступных сведений о числе конфискованных птиц. Обобщение свидетельств предполагает, что масштаб отлова певчих птиц в Китае в настоящее время истощителен для их популяций, и этот отлов, по крайней мере частично, вносит вклад в сокращение. Есть виды многочисленные, но с ограниченным ареалом, чьи популяции быстро уменьшились из-за роста торговли или потребления в Азии (van Balen et al.. 2000; Liu, Weng 2014). Однако, крах такого вида с необычно высокой плотностью населения, как дубровник, чей ареал протянулся по всему континенту, по масштабам катастрофы является подобным таковой у странствующего голубя. Странствующий голубь, как полагают, был самой многочисленной птицей в мире в начале 1800-х годов (популяция оценена в 3 млрд. птиц), но исчез в 1914 г. после десятилетий охоты промышленного масштаба (Schorger 1973). Также как это описано здесь для дубровника, популяция голубя разрушились в период быстрого роста населения, экономического роста и увеличения человеческих перемещений (Schorger 1973).
Есть предварительное свидетельство, что из-за отлова в Восточной Азии могла снизиться численность еще одного многочисленного вида певчих птиц. Dale и Hansen (2013) пишут о коллапсе по всей Скандинавии овсянки-ремеза (Emberiza rustica) — вида, близко связанного с дубровником. Многолетнее исследование на озере Байкал в Сибири показало достоверное и сильное сокращение между 1984 и 2007 гг. пяти видов Emberiza и в целом более отрицательные тренды популяций у дальних мигрантов, чем у ближних мигрантов и оседлых птиц (Ananin, 2011). Все вышеупомянутые виды овсянок имеют сходные маршруты миграций и районы зимовок, и также отлавливаются в Китае. Они менее заметны и не образуют таких концентраций на ночевках, как дубровник, и не образуют плотных скоплений повсеместно в местах гнездования. Поэтому они, вероятно, хуже регистрируются, и во многих областях сокращение остается необнаруженным.
Немногое известно о трендах популяциях, природных угрозах и прессе преследования для обычных и широко распространенных таксонов в Азии (Yong et al. 2015). Налицо срочная необходимость усилить мониторинг биоразнообразия в Восточной Палеарктике. Резкое падение численности дубровника — это современный пример того, какой вклад может внести человеческое преследование в крах широко распространенных и очень многочисленных видов. Птицы имеют важные функции в экосистемах (Sekercioglu et al. 2004); такие насекомоядные виды, как дубровник, регулируют численность вредителей сельского хозяйства. Эти экосистемные услуги могут прекратиться, если резкое снижение численности многочисленных видов будет продолжаться или ускорится в ближайшем будущем, с потенциально глубокими и прямыми последствиями для общества.
Благодарности
Let Birds Fly Fund (Китай) предоставил данные о незаконном отлове. A. Symes модерировал дискуссию на форуме BirdLife Internationals’ Globally Threatened Bird Forum, в ходе которой T. Gray, J. W. Duckworth, R. Ayé, М. Zhang, F. Goes, C. Inskipp, H. Sagar Baral и W. Heim прокомментировали статус изученности вида. N. Batbayar, H. Pönkkä и М. Leivo сделали доступными данные дополнительных учетов. Комментарии P. F. Donald и трех анонимных рецензентов сильно улучшили ранний вариант. Работу Johannes Kamp финансировало Германское Правительство, Федеральное Министерство просвещения и исследований в рамках Sustainable Land Management. Kolja Wolanska получила грант Promos mobility grant of the German Academic Exchange Service (DAAD) на полевые исследования на российском Дальнем Востоке в 2013 г.
Цитируемая литература
Ananin AA. 2011. Long-term trends in ‘background’ breeding bird populations of the Barguzinskii mountain ridge. Bulletin of Buryatiya State University 12:93–99 (in Russian).
BirdLife International. 2004. Threatened birds of the world 2004. CD ROM version. BirdLife International, Cambridge, United Kingdom.
BirdLife International. 2014. BirdLife species factsheet: Yellow-breasted Bunting Emberiza aureola. BirdLife International, Cambridge, United Kingdom. Available from http://www.birdlife.org/datazone/speciesfactsheet.php?id=8954 (accessed February 2015).
Cardillo M, Mace GM, Jones KE, Bielby J, Bininda-Emonds OR, Sechrest W, Orme CDL, Purvis A. 2005. Multiple causes of high extinction risk in large mammal species. Science 309:1239–1241. CIESIN (Center for International Earth Science Information Network). 2000. Gridded population of the world, v3. Available from http://sedac.ciesin.columbia.edu/data/set/gpw-v3-populationdensity (accessed February 2015).
Chan S. 2004. A bird to watch – yellow-breasted bunting. BirdingASIA 1:16–17. Dale S, Hansen K. 2013. Population decline in the rustic bunting Emberiza rustica in Norway. Ornis Fennica 90:193–202.
Dubinin M, Luschekina A, Radeloff VC. 2011. Climate, livestock, and vegetation: What drives fire increase in the arid ecosystems of Southern Russia? Ecosystems 14:547–562.
Fry DM. 1995. Reproductive effects in birds exposed to pesticides and industrial chemicals. Environmental Health Perspectives 103:165–171.
Gao Y, Clark SG. 2014. Elephant ivory trade in China: trends and drivers. Biological Conservation 180:23–30.
Glutz von Blotzheim UN, Bauer K. 1997. Handbook of the Birds of Central Europa. Volume 14, Passeriformes (part 5). Aula, Wiesbaden. (In German.)
Green RE, Newton I, Shultz S, Cunningham AA, Gilbert M, Pain DJ, Prakash V. 2004. Diclofenac poisoning as a cause of vulture population declines across the Indian subcontinent. Journal of Applied Ecology 41:793–800.
Hallmann CA, Foppen RD, van Turnhout CA, de Kroon H, Jongejans E. 2014. Declines in insectivorous birds are associated with high neonicotinoid concentrations. Nature 511:341–343.
Inger R, Gregory R, Duffy JP, Stott I, Voříšek P, Gaston KJ. 2015. Common European birds are declining rapidly while less abundant species’ numbers are rising. Ecology Letters 18:28–36.
Kotiaho JS, Kaitala V, Komonen A, Päivinen J. 2005. Predicting the risk of extinction from shared ecological characteristics. Proceedings of the National Academy of Sciences of the United States of America 102:1963–1967.
Lacy RC. 1993. Vortex: a computer simulation model for population viability analysis. Wildlife Research 20:45–65.
Lau M, Ades G, Goodyer N, Zou F. 1996. Wildlife trade in Southern China including Hong Kong andMacao. Biodiversity Working Group of the China Council for International Cooperation on Environment and Development Project, Hong Kong, PR China. Available from http://www.zd.brim.ac.cn/bwg-cciced/english/bwg-cciced/tech-27.htm (accessed February 2015).
Liang W, Cai Y, Yang CC. 2013. Extreme levels of hunting of birds in a remote village of Hainan Island, China. Bird Conservation International 23:45–52.
Lindenmayer DB, Wood JT, McBurney L, MacGregor C, Youngentob K, Banks SC. 2011. How to make a common species rare: a case against conservation complacency. Biological Conservation 144:1663–1672.
Liu Y, Weng Q. 2014. Fauna in decline: plight of the pangolin. Science 345:884–884. McKinneyML. 1997. Extinction vulnerability and selectivity: combining ecological and paleontological views. Annual Review of Ecology and Systematics 28:495–516.
Nakagawa S, Schielzeth H. 2013. A general and simple method for obtaining R2 from generalized linear mixed-effectsmodels. Methods in Ecology and Evolution 4:133–142. Nijman V. 2010. An overview of international wildlife trade from Southeast Asia. Biodiversity and Conservation 19:1101–1114.
Olson D, Dinerstein E, Wikramanayake E, Burgess N, Powell G, Underwood E, D’amico J, Itoua L, Strand H, Morrison J. 2001. Terrestrial ecoregions of the world: a new map of life on earth. Bioscience 51:933–938.
Pimm SL, Russell GJ, Gittleman JL, Brooks TM. 1995. The future of biodiversity. Science 269:347–349. Rogacheva H. 1992. The birds of central Siberia. Husum Druck-und Verlagsgesellschaft, Husum, Germany.
Rosser AM, Mainka SA. 2002. Overexploitation and species extinctions. Conservation Biology 16:584–586.
Schierhorn FD, M¨uller D, Beringer T, Prishchepov AV, Kuemmerle T, Balmann A. 2013. Post-Soviet cropland abandonment and carbon sequestration in European Russia, Ukraine, and Belarus. Global Biogeochemical Cycles 27:1175–1185. Schorger AW. 1973. The passenger pigeon: its natural history and extinction. University of Oklahoma Press, Norman.
Seekercioglu CH, Daily GC, Ehrlich PR. 2004. Ecosystem consequences of bird declines. Proceedings of the National Academy of Sciences 101:18042–18047.
Siriwardena GM, Baillie SR, Wilson JD. 1998. Variation in the survival rates of some British passerines with respect to their population trends on farmland. Bird Study 45:276–292.
Sodhi NS, Koh LP, Brook BW, Ng PK. 2004. Southeast Asian biodiversity: an impending disaster. Trends in Ecology & Evolution 19:654–660.
van Balen S, Dirgayusa I, Adi Putra I, Prins HH. 2000. Status and distribution of the endemic Bali starling Leucopsar rothschildi.
Oryx 34:188–197. Vitousek PM, Mooney HA, Lubchenco J, Melillo JM. 1997. Human domination of Earth’s ecosystems. Science 277:494–499.
Warkentin IG, Bickford D, Sodhi NS, Bradshaw CJ. 2009. Eating frogs to extinction. Conservation Biology 23:1056–1059. Wood SN. 2006. Generalized additive models: an introduction with R. Chapman & Hall/CRC Press, Boca Raton (FL)/London.
Yiming L, Wilcove DS. 2005. Threats to vertebrate species in China and the United States. BioScience 55:147–153.
Yong DL, Liu Y, Low BW, Espanola CP, Choi CY, Kawakami K. 2015. Migratory songbirds in the East Asian-Australasian Flyway: a review from a conservation perspective. Bird Conservation International 25:1–37.
Zhang L, Hua N, Sun S. 2008. Wildlife trade, consumption and conservation awareness in southwest China. Biodiversity and Conservation 17:1493–1516.
Zhang W, Jiang F, Ou J. 2011. Global pesticide consumption and pollution: with China as a focus. Proceedings of the International Academy of Ecology and Environmental Sciences 1:125–144.


 «Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве
«Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве Некоторые пути сохранения биосферы при урбанизации
Некоторые пути сохранения биосферы при урбанизации Бить или не бить? За деньги.
Бить или не бить? За деньги. Биология как идеология. Часть 4
Биология как идеология. Часть 4 Неузнанный Дон-Кихот
Неузнанный Дон-Кихот Дивергентно-конвергентные механизмы в эволюции биоразнообразия
Дивергентно-конвергентные механизмы в эволюции биоразнообразия Современное изучение городской климатологии
Современное изучение городской климатологии Разное про запаховую коммуникацию
Разное про запаховую коммуникацию