

Пресбиорнис
Перескажу важную статью Н.В.Зеленкова про достижения палеорнитологии последних лет, когда было открыто множество ископаемых групп с мозаичной организацией, проливающих свет на происхождение современных макротаксонов птиц. Происхождение их было во многом неясно из-за обширных хиатусов, а эти находки дают нам переходные формы. Которые, однако, не устраняют хиатуса, он уменьшается, но не делается менее чётким (чем они отличаются от переходных форм в обычном понимании этого слова — нет континуума переходов, разрыв сохраняется). Достаточно часто «соединение» нынешних макротаксонов через данные формы хорошо соответствует «соединению«,установленному до их обнаружения методами молекулярной систематики. Что существенно снизило мой прежний скепсис в отношении её методов построения дерева.
Практически все эти группы обладают мозаичной морфологией, у них отмечается наличие устойчивых признаков и комплексов признаков (морфологических модулей), которые встречаются у других (современных) групп в виде примитивных или продвинутых состояний. Специфика таксона здесь определяется именно уникальным сочетанием примитивных и продвинутых черт
Это рассматривается как единственно возможный путь эволюции в силу запрета на параллельную оптимизацию множества систем организма. Наблюдаемая мозаичность, по крайней мере отчасти, — следствие онтогенетической и эволюционной модулярности.
Обзор промежуточных форм в палеонтологической летописи
Содержание
Наиболее интересные переходные формы известны для ветви Galloanserae. Она включает куро- и гусеобразных, а также ряд вымерших форм, отделившихся от бескилевых и сделавшихся новонёбными независимо от других птиц; имеет хорошую поддержку в данных молекулярной систематики.
Родство куро- и гусеобразных впервые постулировал Симонетта, а показал Феликс Янович Дзержинский (1982) на основании функционального анализа челюстного аппарата. Ключевой таксон для понимания их родства между собой — паламедеи Anhimidae, относимые к гусеобразным. Эти довольно длинноногие птицы имеют некоторые черты гусеобразных, но общей организацией, в т.ч. строением челюстного аппарата близки к куриным, почему считались наиболее примитивными из них. Соответственно, общий предок Galloanserae считали наземной птицей, близкой к курам морфологически и экологией. А утиные и полулапчатые гуси из Австралии рассматривались как вторично специализированные к околоводному образу жизни что черепом, что посткраниальным скелетом.

Черношейная паламедея Chauna chavaria

Рогатая паламедея Anhima cornuta
Чтобы проверить данную схему палеонтологией (т.е. понять, какая из морфологий двух сестринских групп — «утиная» или «паламедейная« — примитивней) надо знать как минимум ближайшую к ним сестринскую группу, ещё лучше — несколько последовательных базальных групп. Нынешние семейства курообразных для этого слишком молоды (врозникли в олигоцене) и слишком специализированы для решения вопроса о ранней эволюции курогусей. Поэтому надо приобщать к анализу примитивных представителей обоих отрядов.
«Со стороны« курообразных наиболее примивно сем. Gallinuloidae, известное, в т.ч., несколькими полными скелетами из среднего эоцена северной Америки и Европы. Они демонстрируют удивительную мозаику признаков, характеризующих как куро-, так и гусеобразных.
Первыми они схожи больше всего с современными сорными курами (сем.Megapodidae) и ископаемыми Quercymegapodidae. Причём их немного; что важно, это морфология клюва (поскольку указывает на сходную пищевую специализацию); наличие спинной кости, хорошо развитый апофиз ключицы (гипоклейдиум) и характерное строение стопы (рис.1).
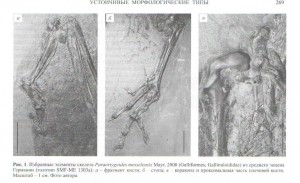 Во всём остальном их скелет или сходен с таковым гусеобразных или (редко) промежуточен по морфологии. Примечательно сходство с гусеобразными в строении черепа (наличие эктэтмоидеума), плечевого пояса и свободной передней конечности (робустные ветви вилочки, удлинённая лопатка с дорзально расположенным мускульным бугром, вогнутый cotyla spapularis, строение биципитального гребня плечевой кости, удлинённый карпометакарпус с узким spatum metacarpale, морфология лучевой кости запястья (рис.1).
Во всём остальном их скелет или сходен с таковым гусеобразных или (редко) промежуточен по морфологии. Примечательно сходство с гусеобразными в строении черепа (наличие эктэтмоидеума), плечевого пояса и свободной передней конечности (робустные ветви вилочки, удлинённая лопатка с дорзально расположенным мускульным бугром, вогнутый cotyla spapularis, строение биципитального гребня плечевой кости, удлинённый карпометакарпус с узким spatum metacarpale, морфология лучевой кости запястья (рис.1).
Часть этих признаков не только характеризуют гусеобразных, но встречаются и у примитивных куриных. Хорошо развитый эктэтмоидеум есть у сорных кур; вогнутый cotyla scapularis характерен для ископаемых семейств Quercymegapodidae, Paraortygidae. Другие черты, общие с гусеобразными, также присутствуют у более примитивных мезозойских птиц; лишь в очень небольшом числе случаев галлинулоиды демонстрируют морфологию, действительно переходную между куро- и гусеобразными (скажем, в строении передней части грудины). Наконец, по третьим признакам они не похожи ни на каких нынешних курогусей (отсутствие spina interna грудины), по другим — сходны лишь с вымершими их представителями (глубокие выемки на грудных позвонках, встречающиеся и у других новонёбных мезозоя).
Следовательно, древнейшие несомненные курообразные по части посткраниальной морфологии сходны или с гусеобразными, или с более высокими (стволовыми) таксонами птиц. Т.е. посткраниальная морфология примитивных Galloanserae была скорее гусеобразного типа (как минимум, строение плечевого пояса и крыла), а то что мы видим у нынешних куриных — недавнее приобретение.
Архаичные ископаемые формы «со стороны« гусеобразных — хорошо известные пресбиорнисы (сем. Presbyornithidae), к настоящему времени подробно изученные (рис.2). Морфологически они тоже мозаичны: продвинутый «утиный» череп сочетается с архаичной посткраниальной морфологией («типа шилоколювок»). Когда были известны лишь фрагменты посткраниального скелета, их относили то ли к фламинговым, то ли к куликам.

Однако после находк полных скелетов выяснилось, что у пресбиорнисов был клюв (и череп) почти как у современных уток – настолько, что кладистический анализ даже реконструирует их как сестринскую группу Anatidae. Это, безусловно, артефакт (известный эффект «умножения синапоморфий»), т.к. вся остальная анатомия этих птиц показывает крайнюю примтивность. Их плечевая кость сходна с таковой примитивных фламинго- и ржанкообразных; туловищные позвонки опистоцельные, что осталось у немногих групп – скажем, у попугаев; грудина с 2мя парами вырезок, как у курообразных (у гусеобразны одна); дистальный тибиотарзус сход с таковым фламинго (рис.2), тарзометатарзус удлинённый, как у длинноногих куликов. И даже квадратная кость, будучи частью челюстного аппарата, у этих птиц удивительно сходна с таковой сорных кур и несходна с квадратной костью прочих гусеобразных.
В анализе филогенетического положения пресбиорнитид есть две возможности. Первое – поверить кладистическому анализу, счесть их сестринской группой утиных. Но полулапчатые гуси, которые в таком случае станут сестринской группой для таксона утиные + пресбиорнисы, имеют много более продвинутую посткраниальную морфологию «утиного» типа. Т.е. если мы признаем эту т.з., то посткраниальная морфология – результат крайней спецализации (или деспециализации, возвращения к более примитивному состоянию).

Полулапчатый гусь Anseranas semipalmata
Второе объяснение экономней – пресбиорнисы сохранили признаки, примитивные для всех новонёбных (скажем, в силу филогенетической инерции). Оно же и лучше, т.к. объясняет большее число фактов меньшим числом предположений – особенно хорошо выраженное сходство пресбиорнисов, с одной стороны, с фламинго и шилоклювками, с другой – с рядом меловых и раннепалеогеновых групп неясного систематического положения, вроде «Graviculidae». Причём эта т.з. не предполагает происхождения гусеобразных от ржанкообразных, как считали Олсон и Федуччиа. Важно, что некоторые черты морфологи пресбиорнисов прослежваются у древнейштх представит елей утиных (п/сем. Romainvillinae).

Шилоклювка Recurvirostra avosetta
Сейчас известно уже много таких мозаичных переходных групп между макротакстонами птиц. Скажем, это Palaeodidae, которые видятся промежуточными между поганками и фламинго, род Turnipax, подтверждающий родство трёхпёрсток и куликов, род Porargornis, демонстрирующй мозаику признаков, указывающую на родство Aegothelidae и Apodidae.
Ископаемое семейство лесных птиц Sandcoleidae сочетает в себе удивительную мозаику признаков дятлов, птиц-мышей, трогонов и некоторых других. Интересны находки ископаемых семейств, демонстрирующих мозаику продвинутых признаков соколообразных (в узком смысле, т.е. только сокола) и попугаев одновременно, что согласуется с нынешними представлениями о достаточно близком родстве этих отрядов. Эоценовое семейство Foratitae характеризуется мозаикой признаков, свойственных современным турако, кукушкам, ястребообразным и ископаемым Idiornithidae из кариамовых. А по форме черепа они очень сходны с современным гоацином – который отнюдь не куриное, а скорей всего, близок к кукушкам-кротофагидам.

Гоацин Opisthocomus hoazin
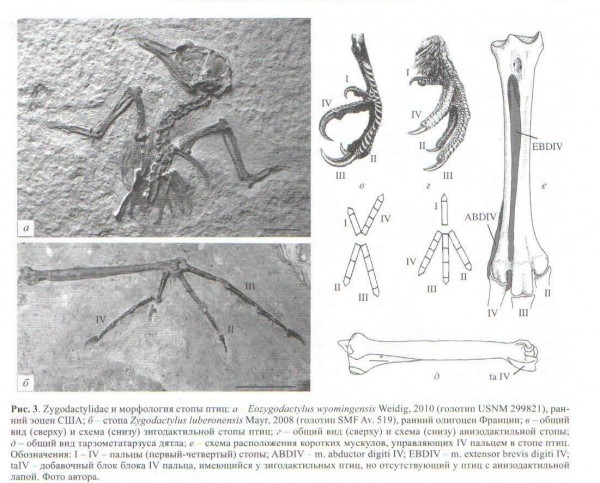
Ещё пример – интереснейшая группа лесных птиц, сем. Zygodactilidae из эоцена-миоцена Европы и Северной Америки, промежуточное междку дятло- и воробьинообразными. Имея ряд примитивных черт, в принципиальных деталях строения скелета они близки ко вторым (никакая другая внешняя группа не показывает более «воробьинообразную» морфологию). В то же время у них зигодактильтная лапа (первый палец вывернут назад), отчасти напоминающая таковую у попугаев и дятлов (рис.3). Это замечательно потому, что у всех современных воробьиных лапа анизодактильная, но по молекулярным данным сестринская группа – зигодактильные попугаи. Зигодактильная стопа была также у нескольких ископаемых семейств переходных между соколо- и попугаеобразными.
 Модули в онтогенезе и эволюции птиц
Модули в онтогенезе и эволюции птиц
Модули – части организма, обладающие структурным и функциональным единством, и эволюционирующие более или менее независимо от других модулей. Эта независимость относительна; совокупность модулей низшего порядка «стягивается» корреляциями в структуру, формирующую модуль высшего порядка. А модуль наивысшего порядка – сам организм, развивающийся как целое и регулирующий свои жизненные отправления, откуда эта подразделённость на модули и их частичная независимость берутся.
Хороший пример такой модульности – устройство стопы у птиц. Недавно показано, что фаланги каждого пальца стопы представляют собой онтогенетический и эволюционный модуль. Экспериментальное изменение длины базальной фаланги одного из пальцев в онтогенезе определяет изменение длин остальных фаланг этого пальца. Переходя от онтогенеза к эволюции, видим ту же зависимость: по длине базальной фаланги можно предсказывать длину дистальных фаланг у самых разных видов современных птиц. С другой стороны, метатарзалии в этот модуль не входят, и изменение их длины никак не влияют на морфологию пальцев.
Подобная модульность и обеспечивает независимую эволюцию фаланг пальцев и плюсны у птиц. В связи с чем интересно, что вышеупомянутые Zygodactylidae демонстрируют разнообразие пропорций фаланг одного из пальцев стопы в пределах одного рода. Что говорит о нестабильности морфогенеза стопы в группе, и может быть предпосылкой к трансформации этого органа в состояние, характеризующее современных воробьиных.
Другой пример модульности в строении стопы – признаки плюсны, связанные с ориентацией наружного (IV) пальца. Плюсневая кость птиц (тарзометатарзус) представлена слившимися дистальными тарзалиями и метатарзалиями. Лишь дистальные части метатарзалий остаются свободными и несут сочленовные поверхности для фаланг пальцев. Расположение этих поверхностей в дистальной части тарзометатарзуса сильно варьирует в разных группах птиц и во многом определяет их экологическую специфику.
В разных группах (кукушки, дятлы, попугаи) независимо 4й палец развернулся назад и возникла т.н. зигодактильная стопа (рис.3в). Её появление связано со сложными перестройками костно-мышечной системы, независимо не встречающимися и, видмо, представляет собой эволюционный модуль. Интересно, что воробьиные вроде бы имеют примитивную анизодактильную лапу, хотя их ближайшие ископаемые и современные родственники зигодактильны. Обратная модификация зигодактильной лапы в анизодактильную затрагивает лишь 1й палец и связанную с ним метатарзалию и, таким образом, является хорошим примером модулярности в эволюции.
Недавнее исследование показало, что по крайне мере у попугаев зигодактильность формируется в онтогенезе так. 4й палец развёртывается назад из-за неравномерности усилий прилагаемых к нему зачатками мускулов. Примитивно у птиц в стопе присутствуют мускулы – анагонисты: m.extensor brevis digiti IV (EBDIV), разгибающий и приводящй 4й палец, и m.adductor digiti IV (ABDIV), отводящий этот палец наружу (рис.3е). У зигодактильных попугаев и дятлов отсутствует EBDIV, поэтому прикладываемое ABDIV усилие не встречает сопротивления, что поворачивает палец назад. Причём деактивация зачатка ABDIV в эмбриогенезе попугая формирует нормальную анизодактильную лапу с 4м пальцем, направленным вперёд.
А вот у воробьиных уникальным образом и ABDIV полностью редуцирован, и EBDIV отсутствует. Предполагается, что этим и объясняется возвращение их стопы к примитивному анизодактильному типу, поскольку в онтогенезе исчезло абдукционное усилие, прироженное к 4му пальцу.
Иными словами, происхождение стопы воробьиных объяснимо перестройками онтогенеза, затрагивающими только этот модуль при сохранении общей конструкции костно-мышечного аппарата стопы практически неизменным. Новейшие открытия в биологии раз0вития показывают, как это могло произойти. Недавно установлено, что процессы морфогенеза костей и мягких тканей конечности, считавшиеся глубоко интегрированными, всё же до некоторой степени автономны. Мускулы конечностей тетрапод формируются в онтогенезе из мигрирующих в дистальные сегменты клеток дермомиотома. Причём судьба мигрирующих клеток (какой именно они дадут мускул) не предопределена заранее, но зависит от в т.ч. от градиентов биохимических факторов, действующих в области их конечной дислокации.
Примером может быть ретиноевая кислота, градиент которой регулирует, в частности, уровень экспрессии Нox-генов, в свою очередь определяющих морфологию пальцев. Эмбриональное расщепление первичного мускульного брюшка на отдельные мускулы происходит под влиянием формирующихся сухожилий (они имеют независимое от мускулов эмбриональное происхождение), а здесь важную роль играет локальное распределение ретиноевой кислоты. В связи с этим важно, что ABDIV располагается на эмбрионально заднем (наружном) крае плюсны, в то время как другой сохранившийся у попугаев плантарный мускул стопы, короткий сгибатель 1го пальца m.flexor hallicus brevis, располагается на переднем (внутреннем) крае.
Поэтому при эмбриональном развитии клетки-предшественники этих двух мускулов (и их конечных сухожилий) окажутся на разных полюсах градиента любых биохимических факторов, уровень экспрессии которых меняется сообразно передне-задней поляризации конечности. А избирательное исчезновение ABDIV у воробьеобразных может быть вызвано изменением параметров биохимической поляризации развивающейся конечности в онтогенезе под влиянием исключительно генетически-обусловленных факторов. Что такое возможно, недавно было показано на конечности парнокопытных млекопитающих. У коровы симметричная морфология дистальных элементов конечности (аутоподия) формируется благодаря нарушению в работе регуляторного модуля, ответственного за передне-заднюю поляризацию.
Неравномерность темпов эволюции разных модулей у птиц была с самого начала истории класса. Скажем, крыло эволюционировало быстро, задняя конечность долго оставалась примитивной. При эволюции макротаксонов, обсуждавшихся выше, быстро формировалась продвинутая морфология челюстного аппарата и черепа в целом, а посткраниальный скелет оставался архаичным.
Макроэволюционные «правила», инадаптивные и эвадаптивные группы
Автор показывает, что современную морфологию макротаксоны птиц обретают постепенно, примитивные признаки очень долго «задерживаются», особенно в посткраниальном скелете (череп и челюстной аппарат обретает продвинутые признаки раньше). В эволюции птиц (как и млекопитающих) наблюдается смена архаичных групп с мозаичной морфологией более прогрессивными, что отсылает нас к концепции инадаптации А.О.Ковалевского.
Надо сказать, что по ряду моментов палеогеновая летопись птиц принципиально сходна с таковой млекопитающих. Тогда были распространены в основном представители вымерших групп, часть которых выделяется или достойна выделения в отдельные отряды. Эоценовые птицы часто демонстрируют адаптации, сохранившиеся у их ныне живущих родственников, особенно в черепе. Это фильтрующий «утиный» клюв пресбиорнисов, вполне современный клюв примитивного ибиса Rhynchaeites и пр. Однако посткраниальной анатомией они радикально отличны от современных форм, почему могут быть причислены к инадаптивным по Ковалевскому.
Однако по Ковалевскому, адаптации таких групп несовершенны, поскольку их «прогрессивная часть» быстро возникает и противоречит всей прочей организации, не подогнана к ней. Соответственно, эти группы быстро вымирают, и сменяются всё более эуадаптивными и гармоничными, т. е. эволюция филума напоминает расчёску, на которой чем ближе к «корню», тем инадаптивнее группы.
Автор показывает, что как минимум для макротаксонов птиц это неверно. Во-первых, адаптации мозаичных групп нельзя считать несовершенными. Длинные ноги пресбиорнисов позволяли им занять другую адаптивную зону «бродячего фильтратора» (а не плавающего, как утиные), где они существовали вполне успешно. Поэтому преобразование инадаптации в эвадаптацию видится как последовательная оптимизация интегрированной системы, которую представляет собой целостный морфололгический тип, присущий тому или иному таксону.
Т.е. мозаичность переходных форм «исправляется» постепенно в эволюции той же самой ветви, и их признаки очень долго прослеживаются у форм современного облика. Поэтому ранние представители эвадаптивных линий сильно отличаются от современных: примитивные утки Romainvillinae сохраняют черты сходства с пресбиорнисами, примитивные фазановые из рода Palaeortyx отличались от большинства современных представителей семейства иными пропорциями задней конечности. Автор считает, что большая часть возникших в конце палеогена семейств, характеризующих современную фауну (фазановые, пастушковые, утиные и пр.) представляют собой результат автохтонной эволюции древних инадаптивных семейств — а не так, что они проявились независимо от них где-то в другом месте, и их вытеснили, как считает Г.Майр.
Соответственно, инадаптивные группы вымирают не в конкуренции с инадаптивными, а в связи с крупными перестройками всего облика биосферы, вызванными, например, климатическими изменениями. Так, многие современные группы появляются в летописи в конце эоцена или в олигоцене, что совпадает с началом глобального похолодания. Дальнейшая эволюция этих эвадаптивных групп представляет собой, с одной стороны, приспособление к климатичяеским и пр. средовым изменениям, с другой — «притирку» к новому таксономическому составу экосистем. Продолжительное существование ряда таксонов на протяжении всего кайнозоя может объясняться постоянством среды. Так, крупные журавлеобразные Eogruidae (включая Ergilornithidae) обитали в аридных ландшафтах Евразии с эоцена по плиоцен. С палеоцена до конца плиоцена жили крупные морские парильщики Pelagornithidae.
На примере ископаемого разнообразия птиц видно, что наличие переходных форм не отрицает хиатусов, плавного континуума переходов не получается. Дискретность наблюдаемого биоразнообразия сохраняется в любом случае, поскольку материалом отбора являются дискретные по своей природе устойчивые фенотипические вариации. Что соответствует эпигенетической концепции эволюции, согласно которой закрепляются эволюционные новшества, исходно бывшие проявлением фенотипической пластичности. Поэтому в происхождении воробьиных бессмысленно искать формы с переходной морфологией стопы. Самое важное, что из ископаемых групп с переходной морфологией очень многие поддерживают филогенетические сближения, сделанные молекулярной систематикой.
Рекомендуем прочесть
 «Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве
«Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве Исследование гендерных стереотипов при направлении на работу
Исследование гендерных стереотипов при направлении на работу Культ книги в РККА: как красноармеец стал главным пропагандистом грамоты в стране
Культ книги в РККА: как красноармеец стал главным пропагандистом грамоты в стране «Кровавые когти и клыки профессора Пинкера». Ч.1.
«Кровавые когти и клыки профессора Пинкера». Ч.1. США против России во имя прибылей ВПК
США против России во имя прибылей ВПК Декоммунизация и антиваксеры
Декоммунизация и антиваксеры Восприятие шумов
Восприятие шумов Эволюция на двух уровнях: о генах и формах
Эволюция на двух уровнях: о генах и формах
