
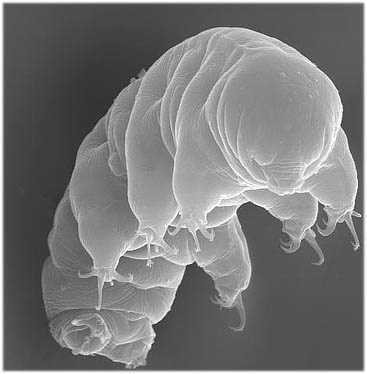
Marina_fr пересказывает недавнюю работу по тихоходкам, авторы которой по отсеквенированному геному смотрели а) есть ли у них Hox-гены, охарактеризованные для более крупной группы Panarthropoda (членистоногие Arthropoda, первичнотрахейные Onychophora, тихоходки Tardigrada), б) есть ли они в транскриптоме, т.е. экспрессируются ли они на разных стадиях развития тихоходки, и где именно. Это привело к интересным выводам о плане строения группы.
«План строения тихоходок возник в результате потери значительной части отделов тела, соответствующих всей груди и почти всему брюшку насекомых. Тихоходки включаются в надтип В настоящее время выполнены работы по сравнению сегментации двух первых групп и экспрессии Hox и других развитийных генов в сегментах их тела. Авторы проанализировали экспрессию Hox-генов у тихоходок и пришли к выводу, что участки тела, гомологичные протяжённым участкам у насекомых и онихофор, у них отсутствует. Общий предок реконструируется ими как обладавший протяжённым телом.
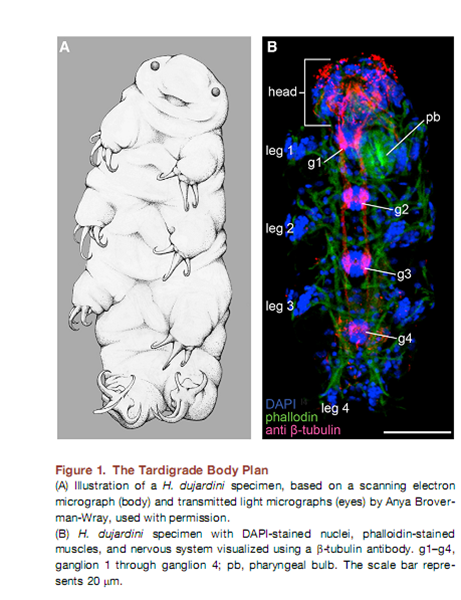 В план строения тихоходок входит голова и четыре сегмента, несущих ноги.
В план строения тихоходок входит голова и четыре сегмента, несущих ноги.
В качестве объекта была выбрана тихоходка Hypsibius dujardini. Авторы работали с её геномом и транскриптомом (где были смешаны разные стадии развития), и в том и в другом случае был идентифицирован тот же набор Hox-генов. Базируясь на филогенетическом анализе и специфических участках, авторы обнаружили гомологи пяти Hox–генов — labial (Hd-lab), Hox3 (Hd-Hox3), Deformed (Hd-Dfd), fushi tarazu (Hd-ftz), и Abdominal-B (последний имел три паралога Hd-Abd-B1–Hd-Abd-B3). Для анцестральных представителей Panarthropoda реконструируется также наличие proboscipedia (pb), Sexcombs reduced (Scr), Antennapedia (Antp), по крайней мере одного гена Ultrabithorax (Ubx) и abdominal-A (abd-A), но они отсутствовали в сборках генома и транскриптома тихоходки.
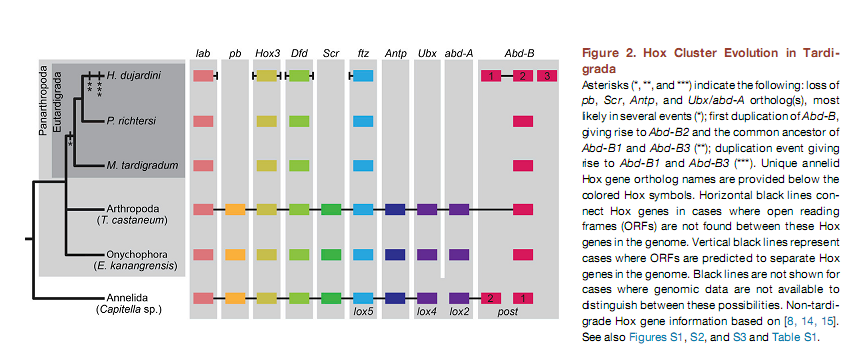
Были изучены также два других вида тихоходок Paramacrobiotus richtersi и Milnesium tardigradum, тоже из класса Eutardigrada, и для них был обнаружен аналогичный набор Hox-генов за исключением того, что Abd-B присутствовал в одном экземпляре. Похоже, что в линии, ведущей к Hypsibius dujardini, произошли дупликации этого гена (не повлиявшие на план строения), тем более, что соответствующий белок у Hypsibius dujardini характеризуется двумя уникальными позициями в гомеодомене. Hd-Abd-B1 наиболее похож на исходный вариант. У тихоходок среди Hox-генов разбросаны белок-кодирующие гены с иной функцией.
Была изучена экспрессия генов в онтогенезе тихоходок. Наблюдались обычные закономерности антериорно-постериорного порядка экспрессии. Через 35 часов после откладки яиц Hd-lab сильнее всего экспрессировался в первом несущем ножки сегменте, через 55 часов наибольшей была его экспрессия в глоточной луковице. Экспрессия Hd-Hox3 была сильнейшей во втором и третьем сегментах, несущих ножки, а через 55 часов – в ганглиях второго сегмента и в ножках второго и третьего сегмента. Во время развития Hd-Dfd обнаруживает локализованную экспрессию в ганглиях третьего сегмента. Hd-ftz экспрессируется в переднем регионе четвёртого сегмента с ножками, причём на 55 часу его сильная экспрессия характерна вблизи ганглиев. Экспрессия Hd-Abd-B1 характерна для задней части четвёртого сегмента, в том числе оснований ножек.
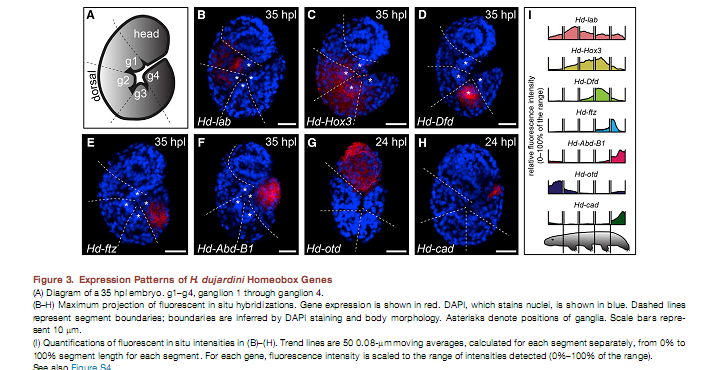
Ранее были изучены передние границы экспрессии аналогичных генов у Arthropoda и Onychophora. У тихоходок они для Hd-Hox3, Hd-Dfd и Hd-ftz такие же. Ось тела у тихоходок гомологична оси головы для этих групп, что хорошо соотносится с идеей, что придатки головы последних произошли от ножек. Похоже, что голова тихоходок гомологична единственному головному сегменту артропод.
У тихоходок была исследована экспрессия головного гена orthodenticle (otd), которая у большинства артропод и онихофор на первых стадиях сегментации ограничена первым сегментом. Hd-otd во время ранних стадий сегментации характеризуется выраженной экспрессией во всей голове тихоходок. Hd-lab экспрессируется во втором сегменте тела тихоходок в отличие от того, что наблюдается у других Panarthropoda. Иначе говоря у тихоходок происходит экспансия передних границ экспрессии этого гена или, наоборот, сдвиг назад этих передних границ у других Panarthropoda. Постериорный маркер caudal (cad) экспрессируется у тихоходок в четвёртом сегменте, несущем ножки.
Присутствует также экспрессия Hd-Abd-B1, что говорит о том, что тихоходки – это всё-таки не просто Panarthropoda, обрезанные с одного конца. У них, однако, потерян регион, характеризующийся у панартропод и аннелид экспрессией Antp, Ubx и abd-A, внутри которого должны располагаться также передние границы экспрессии ftz и Abd-B.
Hox-гены скорее регулируют разметку, чем вызывают образование сегментов. Сомнительно поэтому, чтобы отсутствие pb и Scr у тихоходок соответствовало отсутствию каких-то определённых сегментов. Авторы статьи полагают, что утрата тихоходками генов Antp и Ubx/abd-A была вторичной по отношению к утрате соответствующих сегментов. Как известно, у многих панартропод дополнительный рост тела происходит за счёт добавления постериорных сегментов, что может быть анцестральной особенностью билатерий. Редукция или потеря этой особенности может быть связана с утратой срединных сегментов тихоходками».

Прим.переводчика: Авторы держатся традиционной точки зрения, что ротовые органы насекомых и онихофор гомологичны видоизменённым конечностям, как и ножки тихоходок. Мне представляется, что это требует дополнительных доказательств ввиду специфики их морфологического устройства. Увы, этот момент в статье не освещён.
Рекомендуем прочесть
 Разбор популярных мифов про охрану природы
Разбор популярных мифов про охрану природы Узники совести в Южной Корее: ликбез
Узники совести в Южной Корее: ликбез О фан клубе имени меня, и новостях украинской медицины
О фан клубе имени меня, и новостях украинской медицины Влияние политических взглядов на математические способности
Влияние политических взглядов на математические способности Восприятие расы в США
Восприятие расы в США Урбанизация — не доместикация
Урбанизация — не доместикация Преступники страдают от криминализации социальной среды
Преступники страдают от криминализации социальной среды Экономическая независимость и частная собственность
Экономическая независимость и частная собственность
