

Итальянский воробей
Marina-fr, wolf_kitses
Распространение и изменчивость
Содержание
Итальянский воробей Passer italiae – вид с доказанным гибридным происхождением (Fuglione, Rippa, 2013). Анализ 14 разных локусов микросателлитов в я-ДНК и генов мт-ДНК показывает, что его геном — это не целостность, а равномерная смесь родительских генофондов, взятых соответственно от испанского воробья Passer hispanioleus и домового Passer domesticus.
Родители – испанский или черногрудый воробей Passer hispaniolensis, с каштановой «шапочкой» и большим, но нечётко очерченным «галстуком», и домовый – с серой «шапочкой» и «галстуком» небольшим, но чётким. При этом морфотип итальянского воробья чётко стабилизирован и однороден по всему ареалу, есть только слабые клины изменчивости, ничуть не более выраженные, чем у других видов. Такого всплеска изменчивости фенотипа, какая бывает в гибридных зонах между видами, тут и в помине нет (также как единый и стабилизированный морфотип контрастирует со смешанностью генома P.italiae). Т.е. фенотип italiae соединяет черты той и другой формы (с некоторыми промежуточными признаками), но не в любом сочетании. Скажем, нет сероголовых самцов, но с «галстуками», как у испанского.
Итальянский воробей на севере контактирует с домовым, образуя узкую гибридную зону, стабильную во времени и пространстве, как между серой и чёрной воронами (частью проходящей в том же регионе). На юге Италии он живёт симпатрично с испанским, и с ним не скрещивается. Однако от Неаполя до Сицилии доля фенов испанского воробья в окраске прогрессивно растёт, так что благодаря этой клине воробьи становятся всё более похожи на испанских — однако границы видовых различий так и не переходят!
Вокруг же Италии – в Швейцарии, Болгарии, в Тунисе, на Мальте – мы видим «естественный эксперимент» по синтезу этой формы, когда расселяющийся на север, в ареал домового, черногрудый воробей активно гибридизирует с ним. Учащение гибридизации между родительскими видами насыщает их популяции фенотипом italiae, как бы показывая начальные стадии его образования. Одновременно с фенотипом italiae, который довольно-таки редок, в этих популяциях наблюдается настоящий всплеск изменчивости, можно найти все мыслимые переходы в окраске и морфологии от домового воробья к испанскому.

Домовый воробей, самец
«Гибридность особенно хорошо заметна у тех самцов, которые являются крайними (яркими) типами гибридизации, т.е. когда фенотипически представлены признаки обоих родительских видов. Первый тип – сероголовые гибридные самцы – имеющие серый верх головы (лоб, темя, затылок), как у домового воробья, а горло, зоб, грудь, боковые части тела и плечи у них чёрные, как у испанского воробья. Второй тип – коричневоголовые гибридные самцы – у которых верх головы (лоб, темя, затылок) коричневый, как у испанского воробья, а горло и зоб – чёрные, как у домового воробья. Второй тип … называют «итальянским» воробьём. В природе существуют также и всевозможные комбинации родительских признаков. Множество гибридов (самцов и самок) демонстрируют промежуточные признаки между конечными вариантами. К ним исследователи не всегда относятся с надлежащим вниманием, так что они часто остаются незамеченными».
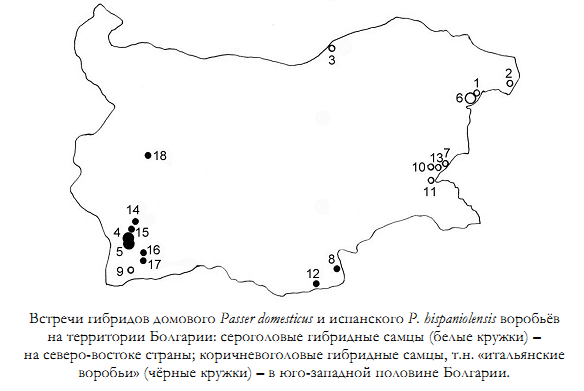
Нанкинов Д.Н., 2013. Гибридизация домового Passer domesticus (Linnaeus, 1758) и испанского Passer hispaniolensis (Temminck, 1820) воробьёв и о так называемом итальянском воробье Passer italiae (Vieillot, 1817) в Болгарии// Русский орнитологический журнал. Т. 22. Экспресс-выпуск 923. С. 2631-2641
Эти фенотипы и преобладают, как это и положено в гибридной зоне; т.е. тут наблюдаются как бы начальные этапы «гибридогенного синтеза», когда соответствующий фенотип ещё не стабилизировался.
Детали биологии родительских видов: системные отличия и их нивелирование
Местообитания
Другой вариант такого «ресинтеза»: в урбанизированных ландшафтах Магриба он оставляет привычку жить в крупных колониях, строя открытые грушевидные гнёзда на деревьях, и гнездится поодиночке в зданиях, под застрехами и пр.
Т.е. как домовый: В.В. Иваницкий обосновал[1], что это «вынужденное сближение» биотопов и мест размещения гнёзд обоих видов подтолкнуло гибридизацию, давшую начало italiae. Поскольку в ином случае оба вида сильно разделены и ландшафтно, и общей организацией процесса ухаживания, почему, как правило, не проявляют интереса друг к другу. Испанский — вид с/х ландшафтов, образует самые крупные стаи и колонии, территориальные связи предельно лабильны: появляется массами, гнездится, потом может исчезнуть.
Так, в Болгарии эти «волны жизни» вызваны социальными трансформациями.
«Пeрвопричиной гибридизации между домовым и испанским воробьями в Болгарии стали существенные изменения среды в местах их обитания, создавшие условия для агрессивной экспансии испанского воробья и частичного разрушения репродуктивной изоляции между этими видами птиц. В середине ХХ века такими существенными изменениями среды, вызвавшими вторжение испанского воробья в Северную Болгарию (а также в Румынию и Сербию), было укрупнение сельскохозяйственных земель в Болгарии (создание колхозов), распашка степей, возникновение огромных площадей, засеянных зерновыми, создание полезащитных лесных полос и орошение ранее безводных земель.
Правда, после 1990 года на протяжении около 15 лет этот процесс резко замедлился и даже остановился, так как колхозы и крупное сельское хозяйство в Болгарии были уничтожены. Огромные площади сельскохозяйственных земель запустели. Сразу произошло резкое падение численности испанского воробья и даже исчезновение его из многих населённых пунктов северной и западной Болгарии. В последние годы в некоторых районах страны снова началось укрупнение хозяйств, большие поля стали засевать зерновыми культурами и подсолнечником. Численность испанского воробья постепенно стала возрастать, и он возвращается в свои старые места обитания».
Нанкинов Д.Н., op.cit.
Рисунок из цитируемой статьи Д.Н.Нанкинова
Жизненная стратегия и социальность в колониях
Домовый более чем консервативен: из 457 птиц, отлавливаемых на ночёвке, встречено через год повторно в среднем 20.6% (18.9-26.1). У испанских воробьёв, меченных в 2-х разных точках ареала у гнёзд на протяжении 3 и 4 лет соответственно (n=255 и 3484 птиц), средняя доля повторных встреч через год составила всего 0.8% и 0.3% (0-1.5% и 0-0.9%)[2].
И стаи, и колонии испанских воробьёв намного больше, чем у домовых, однако первый вид выглядит сильнее менее «социальным», чем второй. Территориальность самцов и пар в колониях предельно редуцирована. Практически их не интересует ничего, кроме мутовки ветвей, в которой потом разместится гнездо. Именно на неё (а не на партнёра) ориентирован процесс образования пары, большую часть которого самец пробует пробиться к мутовке, чтобы продолжить гнездостроение, а самка пытается ему помешать и сама утвердиться там.
Соответственно, персональные связи соседей в огромной колонии испанского воробья практически отсутствуют, как и регулярные территориальные контакты из-за пространства. Напротив, у домового воробья территориальное поведение весьма развито, обычны персонализованные связи и регулярные агонистические взаимодействия соседей на границах территорий, формирующие определённую иерархию. Последние у этого вида (но не испанского) «означиваются» специальными демонстрациями «патрулирования».
«Патрулирование границ» разных видов воробьёв
«…демонстрации патрулирования разных видов воробьёв рода Passer связаны с территориальными отношениями и демаркацией границ между охраняемыми пространствами соседних особей[3] (Иваницкий, 1997). В ряду видов воробьёв показана строгая положительная связь между (а) выделенностью специфической категории проблемных ситуаций патрулирования границ из общей канвы социальных взаимодействий (так что оформляются специальные взаимодействия патрулирования, разрешающие именно эту проблему) и развитостью специфических демонстраций, употребляемых именно в них и, надо думать, необходимых для их эффективного завершения (б).
Сравнительный анализ репертуаров разных видов показывает, что появление специфических демонстраций патрулирования чётко связано с бόльшей выделенностью ситуаций патрулирования границ в системе социального поведения вида. Форма этих демонстраций у разных видов различна, хотя в основе всех их лежат одни и те же телодвижения, выражающие общий мотивационный конфликт «нападение-бегство». Степень «оформленности» и «знаковости» демонстраций патрулирования, их обособленность от других демонстраций видового репертуара строго коррелирует с выделенностью взаимодействий патрулирования границ в социальной жизни вида, с их значимостью для взаимоотношений самцов и самок в связи с подбором оптимального полового партнёра (Иваницкий, 1997).
У одних видов воробьёв патрулирование происходит целенаправленно, занимает относительно много времени, это обычный, часто повторяющийся вид активности. Одновременное патрулирование границы соседними самцами рождает специфические взаимодействия, чётко фиксированные во времени и привязанные именно к данному месту. Их главная особенность — отчётливое противостояние оппонентов друг другу, связанное с сильным мотивационным конфликтом.
Для сходного случая ритуализированной агрессии грызунов (песчанок п/сем Gerbillinae) показано, что сильный и продолжительный конфликт типа «нападение-бегство», «приближение-избегание» психологически истощает животное и должен быть «снят» обменом ритуализованными демонстрациями. После (и в результате) этого «снятия» возникает поведенческая асимметрия между противниками, которая представляет собой специфический результат процесса коммуникации, выигрыш одной особи и проигрыш другой. По характеру асимметрии легко определяется, кто именно выиграл и проиграл (Попов, 1986; Громов, 2005). Подобное завершение «конфликта интересов», вызвавшего столкновение особей, исключает повторение взаимодействия на определённый период времени (= последействие социальной коммуникации), так как оппоненты, прежде «равные» в статусе, дифференцировались на выигравшего и проигравшего способом, «общепризнанным» в данной системе-социум – через организованный обмен специфическими демонстрациями, соответствующий «правилам игры» (Попов, 1986; Гольцман и др., 1994; Goltsman, Volodin, 1997).
Аналогично разрешаются территориальные конфликты у тетерева Lyrurus tetrix (Лысенко, 1985, 1987) другие агонистические и брачные взаимодействия у птиц вообще (Hurd, Enquist, 2001; Фридман, 2007, 2008а). Подобное «снятие» должно производиться и здесь.
И, действительно, у «патрулирующих» видов появляются специфические демонстрации, используемые именно для этой цели: они маркируют ситуацию конфликта и делают явственной поведенческую асимметрию, созданную взаимодействием. У индийских воробьёв Passer indicus сюда относятся позы К-Л рис.11 в работе В.В.Иваницкого (1997: 91), у рыжих воробьёв P.rutilans позы Д и З рис.14 (ibid.: 93), саксаульных P.ammodendri – позы Г-Ж рис.13 (idid.: 92), полевых P.montanus – позы П и Р рис.12 (ibid.: 92). Также к числу патрулирующих видов В.В.Иваницкий относит домовых воробьёв Passer domesticus (ibid.: 97).
Непатрулирующими являются такие виды, как испанский воробей Passer hispaniolensis, каменный воробей Petronia petronia, короткопалый воробей Carpospiza brachydactyla, снежный воробей Montifringilla nivalis, монгольский земляной воробей Pyrgilauda davidiana (Иваницкий, 1997: 97 и рис.10). Показательно отсутствие у них специфических демонстраций: хотя особи так или иначе вступают в конфликты друг с другом, в том числе и в пограничные, «невыделенность» патрулирования как отдельного аспекта социальной жизни вида ведёт к «неозначенности» его специфическими сигналами.
Важно обратить внимание на видоспецифичность форм демонстраций, связанных с патрулированием и пограничными конфликтами у видов первой группы, но не второй. Хотя ситуация патрулирования, маркируемая соответствующими демонстрациями, у разных видов вполне однотипна, их внешний облик во всех этих случаях различается, демонстрации патрулирования видоспецифичны, а межвидовые различия в форме сигнала устойчивы и независимых от динамики мотивационного состояния индивидов (которое неустойчиво и в конфликтной ситуации сильно колеблется).
Мотивационный конфликт при патрулировании у воробьёв наиболее силён у видов, где патрулирование «выделено» в отдельную область социальных взаимодействий, играющую специфическую роль в социальной жизни особей. Это домовый Passer domesticus, индийский P.indicus, саксаульный P.ammodendri, рыжий P.rutilans и полевой P.montanus воробьи. Патрулирование здесь обособлено от других форм активности, пограничные конфликты фиксированы, имеют чёткие начало и конец.
Следовательно, здесь патрулирование образует особый контекст общения, в противоположность второй группе «непатрулирующих видов»: испанский Passer hispaniolensis, каменный Petronia petronia, короткопалый Carpospiza brachydactyla, снежный Montifringilla nivalis, монгольский земляной воробьи Pyrgilauda davidiana. Здесь патрулирование как особый вид активности отсутствует, стычки за территорию мимолётны и не связаны со специфической ситуацией противостояния, заслуживающей выделения в отдельную категорию «пограничных конфликтов».
В первой группе всегда присутствуют специфические демонстрации, жёстко связанные с пограничным конфликтом, они сугубо видоспецифические и разные у разных видов. В агрессивных взаимодействиях между самцами или самками индийского воробья соперники, сблизившись на границе, демонстрируют друг другу «поклоны», отсутствующие у «непатрулирующих» испанских воробьёв. «Непатрулирующим» видам также свойственны пограничные стычки, интенсивные и частые, но их поведенческий репертуар никогда не содержит ритуализированных демонстраций, тем более специфических для вида или ситуации. Так, самцы испанского воробья просто гоняются друг за другом, при длительных конфликтах они время от времени присаживаются в повседневной позе или малоинтенсивной позе №1 (Иваницкий, 1997: 88-97)».
Фридман В.С., 2013. От стимула к символу. Т.1. М.: изд-во URSS.
Гнездостроение
У испанского воробья этим занимается преимущественно самец, а брачное поведение и строительство гнезда сливаются в единый комплекс (андролатеральное гнездостроение). Ещё до образования пары он строит гнездовую платформу плоской или чашевидной формы; доставка и укладка гнездового материала чередуется с пением и брачными демонстрациями. Холостому самцу приходится всё время чинить платформу или строить её заново из—за расхищения материала соседями. Но даже если этого нет, самцы останавливают строительство на стадии платформы или чашечки, посвящая дальнейшее время саморекламированию или бесцельным манипуляциям с гнездовым материалом. Лишь после образования пары самец возобновляет строительство и быстро доводит его до конца. Однако не сразу; хоть он и испытывает сильное побуждение строить, в «ту самую мутовку» его не пускает самка, оккупирующая её и не дающая самцу приближаться слишком близко. Самец в сущности ловит моменты, когда самка покидает колонию, чтобы возобновить строительство, и не сопровождает её. Вернувшись, самка тут же сгоняет самца и усаживается на платформу.
Прочерк — нет демонстрации, гомологичной таковой у близких видов. Здесь и далее иллюстрации см. Иваницкий, 1997.
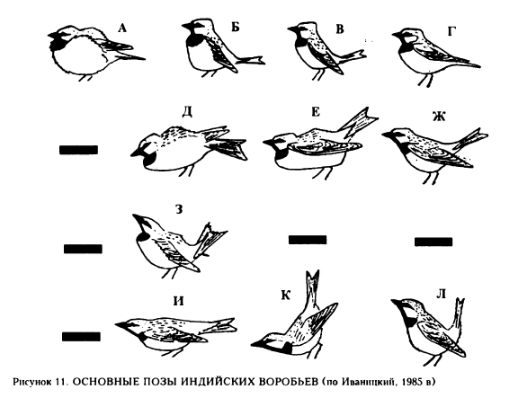
У домового воробья демонстрации практически однотипны
Вскоре, однако, она делается терпимей, и в паре устанавливается чёткий режим пребывания у гнезда. Когда подлетает самец, самка сваливает и даёт ему строить; когда подлетает она, самец улетает за материалом. В этот период гнездо под присмотром обоих, что останавливает расхищение и позволяет достроить.
У домовых воробьёв гнездостроение билатеральное. Холостые самцы в основном рекламируют и чистят убежища. Хотя они и носят туда гнездовой материал, и сооружают плоскую или чашевидную гнездовую платформу, очень скоро бросают это дело и начинают петь, тем более что их ухаживание ориентировано на самку. По-настоящему гнездо начинает строить самка после образования пары, самец лишь к ней присоединяется, хотя через некоторое время уже носит материал интенсивней её. Самка же сокращает своё участие.
Так, из 6 пар домового воробья, строящих гнездо, материал приносили только 4, с частотой 6.3±1.3 в час, самки 3.8±1.0 в час. (27 ч наблюдений). У испанского воробья из 18 самцов носили материал все, со средней частотой 8.4±1.5 в час, самки — 2.7±1.8 (98 ч наблюдений).
При совместном строительстве партнёры избегают делать одно и то же, почему дифференцируют виды активности, а если делают, то один трудится много интенсивней другого. Всё это — проявления принципа стимуляции подобного подобным М.Е.Гольцмана. См. Иваницкий В.В., 1997. С.64-70.
Резюме по различиям биологии и среды обитания
Иными словами, «в принципе» испанский воробей не менее, а может, и более агрессивен, что легко видеть по постоянным стычкам самых разных птиц в колонии и уровню агрессивности партнёров при образовании пары. Однако «на базе» этой агрессии не формируется долговременно-устойчивых социальных связей; поэтому она остаётся «чистым возбуждением», «неопределённым» в том смысле, что неритуализованным, не выраженным в чётко дифференцированных формах демонстраций (как неопределённая материя, materia non signata). Структура сообществ домового воробья во многом противоположна — персонализованные контакты и долговременные социальные связи на базе территориальности и/или агрессивного доминирования, поддерживаемые типологически определёнными демонстрациями. См. Иваницкий В.В., 1997, С.22-25.
Однако же там, где сельхозландшафт густо насыщен микроурбанотерриториями (каменные здания и участки застройки, приближенной к городской, мелкие предприятия, эти различия нивелируются. Оба вида живут примерно в одной среде, гнезда размещают «по типу домового», а уменьшение колоний и рассредоточение гнёзд способствует индивидуализации поведения у испанского. Снижаются и различия в сроках гнездования: исходно перелётный испанский на Балканах появляется на колониях, когда домовые уже насиживают, и смешанные пары возникнут лишь после гибели гнёзд или с не нашедшими пару (во втором и третьем циклах размножения их появление более вероятно). Однако на Апеннинах и в Северной Африке испанский воробей урбанизируется и делается оседлым; также оседлость увеличивается к югу ареала (и гибридизация с ним, создавшая italiae, шла с юга).
Образование пар
Будучи наименее социальным видом, но живущим в крупных скоплениях, испанские воробьи предсказуемо «самые эмоциональные» — характеризуются наибольшим уровнем возбуждения при исполнении гомологичных с прочими видами демонстраций. У большинства воробьёв во время ухаживания крылья отводятся немного в стороны (включая кистевые сгибы) и удерживаются в этом положении неподвижно или трепещут.
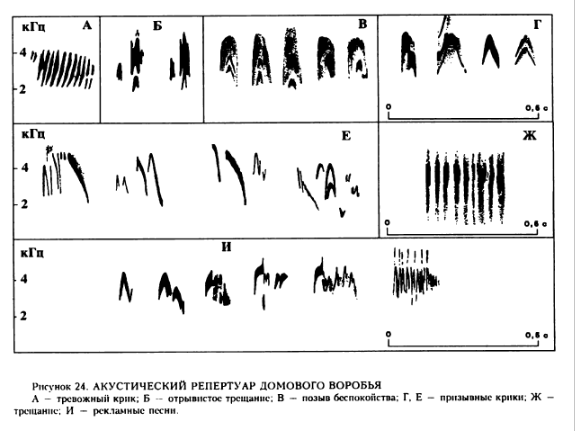
Из наших видов воробьёв именно испанские сильней всего отводят крылья в стороны и трепещут ими; вообще у них брачные демонстрации наиболее интенсивное, а ухаживание — самое «суматошное» с максимумом насилия, в сравнении со спокойным и мирным домовым. См. Иваницкий, 1997, С.89. То же верно для акустической вокализации: характерная особенность многих вариантов песни испанских воробьёв — присутствие особых концентраций энергии в диапазоне 6-8 кГц, отчего их песни приобретают очень характерное «заливистое» или «истеричное» звучание. Даже среди голосов десятков индийских воробьёв в колонии голос пение единственного самца испанского воробья выделяется вполне отчётливо. Помимо различий в структуре рекламных песен, существенны различия в системе позывов; у домового воробья их существенно больше, они лучше дифференцированы и отличаются акустической формой, что согласуется с «большей социальностью» этого вида. См. Иваницкий, 1997. С.108.
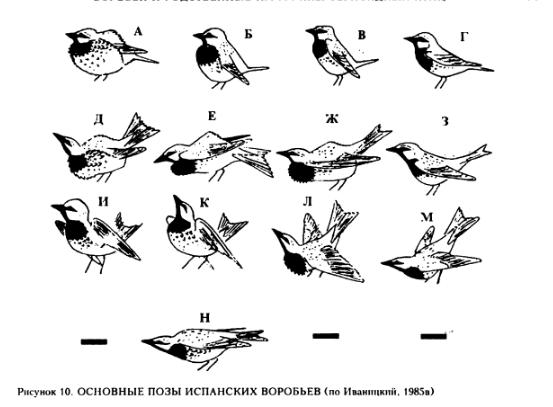
 Образование пар этого вида характеризуется явным антагонизмом партнёров. При виде самки самец залезает в искомую мутовку, где интенсивно демонстрирует в позах Д-М (рис.10). Самка подлетает немедля и пробует проникнуть в ту же мутовку. Она пытается вытеснить оттуда самца всеми доступными способами, в том числе поколачивая его клювом по голове — сперва легко, но по мере растущего возбуждения, со всей дурацкой мочи. Однако это лишь усиливает демонстрации самца, не отвечающего самке, а лишь интенсифицирующего их, и как бы впадающего в настоящий экстаз. См. здесь и далее Иваницкий, 1997, С.34-36.
Образование пар этого вида характеризуется явным антагонизмом партнёров. При виде самки самец залезает в искомую мутовку, где интенсивно демонстрирует в позах Д-М (рис.10). Самка подлетает немедля и пробует проникнуть в ту же мутовку. Она пытается вытеснить оттуда самца всеми доступными способами, в том числе поколачивая его клювом по голове — сперва легко, но по мере растущего возбуждения, со всей дурацкой мочи. Однако это лишь усиливает демонстрации самца, не отвечающего самке, а лишь интенсифицирующего их, и как бы впадающего в настоящий экстаз. См. здесь и далее Иваницкий, 1997, С.34-36.
Если поколачивания не помогли, самка хватает самца клювом за хвост и выбрасывает из мутовки, тут же занимая его место. Самец возвращается и начинает столь же интенсивно демонстрировать около сидящей в мутовке самки, но не пробует её вытеснить. Образование пары можно считать состоявшимся, когда самка получает право относительно свободного доступа к месту гнездостроения.
Всего у испанского воробья отмечено 12 случаев, когда самки выбрасывали самцов из мутовок, в 7 из 10 отслеженных эпизодов образования пары, причём в 4 случаях самки удерживали самцов на весу по нескольку секунд (до 12). Т.е. у этого вида самка утверждает себя в роли доминанта с первых часов семейной жизни, у других видов это происходит постепенно и полностью самка начинает рулить с начала насиживания. Единственное исключение — копуляция; в момент садки самец агрессивен, интенсивно клюёт самку в спину, затылок и темя. Домовый воробей демонстрирует около убежищ и существенно менее интенсивно; важной частью ухаживания обоих видов являются коллективные демонстрации самцов.
Гибридогенный вид между Сциллой и Харибдой
Так или иначе, в сегодняшних зонах контакта и гибридизации родительских видов вокруг ареала italiae встречаются все варианты гибридов, в т.ч. А скрещивание в лаборатории родительских видов показывает пониженную фертильность гибридных самок (т.е. гетерогаметного пола, как и должно быть по правилу Холдейна), хотя у нынешних итальянских воробьёв нормальная. Возникает вопрос: как эта форма восстановила репродуктивную способность до «средне-воробьиной нормы» и как это связано (если связано) со стабилизацией фенотипа, установлением единообразия?
Надо сказать, итальянский воробей – единственный пример гомоплоидных гибридов[4] у птиц, обособившихся до видового уровня (и один из немногих примеров гибридогенных видов у птиц, поскольку у них не встречаются аллополиплоидные гибриды). Скажем, у млекопитающих их существенно больше. Почему? Ведь зон контакта с гибридизацией (в т.ч. долговременно устойчивых, где гибридные фенотипы настолько типичны, что систематики давали им имена) у птиц существенно больше?
Дело в том, что при формировании гомоплоидная гибридная форма должна как бы пройти между Сциллой и Харибдой. Если родительские формы достаточно близки, ей крайне сложно образоваться и обособиться, если достаточно далеки – ей сложно размножаться «в себе» и поддерживать численность, делающую популяцию жизнеспособной.
Когда формы настолько близки, что дают гибридов, плодовитых и жизнеспособных не менее чем родители, механизм «распознавания своих и чужих» «растаскивает» особей промежуточного фенотипа, уклоняющихся в сторону одного из родителей, между популяционными системами этих последних. «Чистых гибридов» оказывается слишком мало, они разрознены и вкраплены в группировки родительских форм, они не могут обособиться и создать свою популяцию, по уровню целостности и распознаванию «своих» и «чужих» сравнимую с родительским. Тем более что в гибридных зонах у птиц численность особей всех фенотипических классов, как правило, на порядок ниже, чем за пределами зоны в тех же местообитаниях. Зачастую гибриды отвергаются как половые партнёры особями обоих родительских фенотипов даже в случаях, когда те «не различают» (или не отвергают) друг друга, и плодят этих самых гибридов.
Так, предшествующие исследования выявили асимметричную реакцию оленух в эструсе на рёв самцов разных видов. Большинство самок благородного оленя (но не все) предпочитают рёв своего вида перед брачным сигналом самцов пятнистого, а вот самки пятнистого предпочтения не выказывают (думаю, потому, что более мощный и низкий рёв второго вида их слишком сильно неспецифически возбуждает).
Однако в опытах с двумя проигрывателями, где самки в эструсе должны выбрать между брачным криком своего вида и гибридов, оба вида выбирали «своих» и отвергли гибридов. Чем опровергли рабочую гипотезу авторов, что вышеназванная симметрия сохранится и здесь, оленухи первого вида предпочтут «своих» гибридам, а второму опять всё будет по барабану. Так получается в том числе потому, что стереотипность воспроизведения акустических форм сигналов у гибридов снижена в сравнении с обоими видами (а значит, и точность распознавания). У млеков, как и у птиц, «притяжение к своим» и «распознавание чужих» — разные процессы, и второй осуществляется сильно более точно.
Поэтому при репродуктивно полноценных гибридах между формами (достигшими в обособлении видового статус, но «защищёнными» от полной интерградации, типичной для подвидов, только распознаванием «своих и чужих» на уровне популяции, без возможности отбора против гибридов) последние больше «подпитывают» популяции родительских видов и «поглощаются» ими. В первую очередь это верно для особей с гибридным индексом ближе к 0 или 1, т.е. к одному из родителей, а полностью промежуточных слишком мало, да и они направленно «исключаются» из группировок обоих. Поэтому не выходит наоборот – чтобы в гибридной зоне обособлялись гибриды и создали «третью», гибридогенную форму; такое бывает лишь если родители не достигли видового уровня, но тогда в зоне вторичного контакта просто возникнет ещё один подвид.
Если же формы дивергировали друг от друга настолько, то их гибриды – с заметно сниженной фертильностью или т.д. приспособленностью, то непонятно, за счёт чего они восстановят то и/или другое, чтобы гибридная популяция была жизнеспособной при размножении «в себе» (хотя бы на уровне родителей). Т.е. будущая гибридогенная форма оказывается перед «вилкой»: в первом случае она не может возникнуть, во втором – возникнув, не очень может размножиться.
Как смог итальянский воробей выкрутиться?
Как итальянский воробей вышел из этой ловушки? Как известно, у итальянского воробья то же хромосомное число, что и у родительских видов, и основным сценарием его происхождения является образование гибридов с их последующей частичной репродуктивной изоляцией. Исходная плодовитость этих гибридов, а в соответствии с правилом Холдейна гетерогаметного пола в особенности (у птиц это самки) проблематична. В работе Eroukhmanoff et al., 2013 показано, что половина самок стерильна, но эта половина – 3 птицы из 6. Более того, 5 из них были гибридами самки испанского воробья и самца городского (из них как раз 3 – стерильны), 1 плодовитая самка – гибридом самки городского воробья и самца испанского.
Между тем, подавляющее большинство исследованных в Elgvin et al., 2011 итальянских воробьёв имеют по митохондриальному гену ND2 генотип городского воробья, хотя и с резко отличающейся частотой полиморфизмов. В другой работе тех же авторов, где была использована более обширная выборка, лишь один из 41 итальянских воробьёв имел митохондриом испанского. Это означает, что они ведут свой род именно от самок, появившихся в результате скрещиваний второго типа. Про их плодовитость мы, практически, ничего не знаем, кроме того, что они могут быть плодовиты.
В этих скрещиваниях самки (они гетерогаметны) должны были унаследовать свою Z-хромосому от отца, то есть от испанского воробья вместе с «испанскими» аллелями её генов. В дальнейшем рекомбинация у гомогаметного пола должна была нарушить это единообразие, но по крайней мере для одного из блоков сцепления Z-хромосомы этого не произошло. Это блок, содержащий ген CHD1Z, аллели которого отличаются двумя фиксированными заменами у испанского и городского воробьёв. У итальянского воробья найден исключительно аллель, характеризующийся теми же фиксированными заменами, что для испанского и обычными для последнего вида полиморфизмами. Укажу, что для каждого вида Eglvin et al., 2011 были исследованы 14 самцов и 2 самки, что, конечно, не вполне достаточно для окончательного суждения.
Тем не менее, автор данного текста может предположить, что именно комбинация «митохондриом итальянского воробья – содержащий CHD1Z блок сцепления испанского воробья» находилась под положительным отбором, обеспечивая: 1) достаточную плодовитость гибридных самок, 2) ограниченное скрещивание с родительскими формами.
Существует и другой Z-хромосомный ген, для которого у родительских форм фиксированы разные варианты – это PLAA. Оба присутствуют у итальянского воробья, но преобладает вариант, характерный для воробья городского (из определённых аллелей 19 как у городского воробья, 8 – как у испанского). Не исключено, что по нему тоже шёл положительный отбор, но в последующих поколениях.
Утверждения Eglvin et al., 2011 о возможном положительном отборе CHD1Z и PLAA базируются на статистическом анализе. Более уязвимым выглядит его предположение о том, что отбирались именно эти конкретные гены, а не какие-то ещё, содержащиеся в том же блоке сцепления. Итальянский воробей – явно недавний вид, что подтверждается высоким уровнем его аутосомного полиморфизма, связанного с тем, что им был «унаследован» полиморфизм обоих родителей (Eglvin et al., 2011). Характерно также наличие у него короткого абортивного сперматогенеза и относительно высокого уровня андрогенов зимой, что говорит о недавнем происхождении от более южного вида (см. Fulgione, Rippa, 2013). В связи с небольшим числом актов рекомбинации, прошедших в Z- хромосоме с момента образования вида, степень сцепления близлежащих аллелей может быть весьма значительна. Тем не менее, Eglvin et al., 2011 отмечают, что CHD1Z связан с ремоделингом хроматина, и в этом плане интересно, что по распределению гетерохроматиновых блоков итальянский воробей хорошо отличается от городского, но не от испанского (см. Fulgione, Rippa, 2013). Это может быть связано с иным состоянием CHD1Z, но является ли это непосредственным или побочным эффектом отбора – сказать трудно. В то же время, отличия по Z-хромосоме у итальянского воробья есть от обоих родительских видов.
В заключение следует отметить, что предполагаемый автором этого текста механизм приобретения несовместимости с родительскими видами (сочетание митохондриома одного с Z-хромосомным блоком другого) может работать исключительно для группы с гетерогаметностью самок и невозможен, например, у млекопитающих, для которых известно больше гибридных видов гомоплоидного происхождения.
Что также важно в этой истории — ещё раз удалось показать, что для видообразования стабилизация морфотипа, отделение его хиатусом от морфотипов родительских видов (с одновременным «отщеплением» его популяционной системы от таковой родительской формы) намного важнее формирования отдельной генеалогической линии с защищённым геномом (от скрещивания с другими линиями).
Примечания
[1]См. Этологические зарисовки// Природа. 1988. №9.
[2]Иваницкий В.В., 1997. Воробьи и родственные им группы зерноядных птиц: поведение, экология, эволюция. М.: Т-во научных изданий КМК.
[3]Все нижеследующее — моя собственная интерпретация данных В.В.Иваницкого (1997), за которую автор не несёт ответственности. Он, скорей, вообще не касается темы сигнальности демонстраций, их возможных «значений» и пр., в соответствии с теоретической позицией, соответствующей неспецифической концепции коммуникации (Иваницкий, 1982, 1989). Я специально использовал эти данные, чтобы показать критически настроенному читателю, как любое тщательное сравнительно-этологическое исследование даёт достаточные основания для утверждения «сигнальности» и / или «знаковости» демонстраций, даже если теоретическая позиция толкает автора отрицать первое и второе.
[4]Т.е. с сохранением у гибрида числа хромосом в кариотипе, характерного для родительских видов, гибридогенная форм «рождается» комбинацией частей геномов родителей без изменения уровня плоидности ; антоним — аллополиплоидный гибрид, где число хромосом у потомства от межвидовых скрещиваний кратно меняется. См. «Homoploid speciation – what is it, and why does it matter?»








