

Два средних поморника тёмной и светлой фазы, в угрожающей прямой позе (слева) и вертикальной прямой позе (справа) соответственно
Введение
Содержание
- 1 Введение
- 2 Примеры и результаты межвидового территориального взаимодействия у тундровых поморников
- 3 Район исследования и население поморников
- 4 Материалы и методика исследования
- 5 Результаты
- 5.1 Внутривидовые демонстрации
- 5.2 Реакция на воздушных хищников
- 5.3 Результаты экспериментов с чучелами птиц
- 5.4 Реакция поморников на представителей не своего вида в природе
- 5.5 Предпочтительная среда обитания и влияние межвидового взаимодействия на распределение гнездовых территорий
- 5.6 Внутривидовая территориальность и район, защищаемый от хищников
- 5.7 Отношения между поморниками и другими хищниками
- 6 Обсуждение
- 7 Литература
Публикуем перевод статьи В.К.Рябицева о внутри- и межвидовой территориальности у поморников (выполнен Кирой Аметовой), исключительно важной по двум причинам. Первая частная: данный номер «Русского орнитологического журнала» давно стал библиографической редкостью. Он отсутствует в большинстве библиотек и электронных хранилищ, где есть номера лишь с 1996 г., и его скан был получен благодаря любезности зав.отделом орнитологии Зоомузея МГУ П.С.Томковича.
Вторая главная: эта стать не столько о территориальности у поморников, сколько о том, почему ритуализованные демонстрации, использованные в социальном общении, не просто стимулы (или не только стимулы), выражающие состояние особи вовне и воздействующие им на оппонента вроде укола, тычка или толчка, но в первую очередь знаки дифференцированных ситуаций внешнего мира. Видовые демонстрации их как бы «именуют», маркируя типологически определённой формой достаточно независимо от мотивационного состояния демонстратора. Это «именование» равно существенно для самой особи и для реципиента сигнала и, как показывает статья, много важней «выражения» вовне внутреннего состояния индивида, даже его изменений в связи с ситуацией, скажем, растущей (снижающейся) агрессии (или страха).
Как именно это показывается? Если коротко, автор последовательно демонстрирует, что
1) мотивационно единые (агрессия в отношении «источника проблемы»), но семантически разные ситуации (охрана территории от конспецификов vs окрикивание хищника, в т.ч. двух других видов поморников крайне сходного облика) маркируются разными демонстрациями, визуальными и акустическими.
2) однотипные ситуации у разных видов поморников маркированы разными демонстрациями (видоспецифическими), несмотря на принадлежность видов к категории «двойников» с крайне сходными обликом, обыденными телодвижениями и экспрессивными реакциями, отражающими общее возбуждение.
3) у всех трёх видов поморников конкретный тип ситуации, мотивируемой одинаково (агрессия) распознаётся по «маркирующим» её формам видовых демонстраций, не другим сопряжённым с нею событиям (скажем, характером локомоции или открытой агрессии), весьма сходным в атаках на хищников и потенциальных захватчиков.
Результаты этой работы, как и многих других, заставляют рассматривать демонстрации видового репертуара как сигналы ситуаций даже там, где вроде бы сами обстоятельства побуждают к иному, как с поморниками. То же относится к изученным В.В.Иваницким (1997) демонстрациям патрулирования у воробьёв. Хотя это верно не для всех сигналов и контекстов общения, важно подчеркнуть, что так бывает, и достаточно часто, сигналы, имеющие внешнего референта, закономерно возникают при совершенствовании коммуникативных систем какого-то из контекстов общения в самых разных филогенетических «веточках».
Где-то это контекст предупреждений о разных классах опасности, где-то — о разных шансах выиграть территориальный конфликт с данным противником, где-то — о разных возможностях сближения с ухаживательными демонстрациями с данной самкой и пр. Но везде идёт эволюционный переход от визуальных или акустических демонстраций, характерная форма которых выражает вовне разные уровни мотивации (в ритуализованном виде, т.е. символически, не напрямую, какк соответствующие действия), к демонстрациям, тем же способом «символизирующим» (или «именующим») альтернативные категории ситуаций вовне, существенные для особей.
***
Примеры и результаты межвидового территориального взаимодействия у тундровых поморников
В. К. Рябицев
Три вида поморников рода Stercorarius – короткохвостый S.pomarinus, средний S.parasiticus и длиннохвостый – часто населяют одну и ту зже местность, так что гнездовые территории сильно перекрываются (в сравнении с другими видами поморников, рода Cataracta). Их ареалы сильно перекрываются на обширных пространствах голарктической тундры.
На гнездовании все три вида строго территориальны, охраняемое пространство большое лишь в некоторых приморских регионах средний поморник бывает колониальным (Andersson and Gotmark, 1980; Cramp and Simmons, 1983; Furness, 1987). Pitelka et al. (1955) пишут о межвидовой агрессии среди поморников Аляски и взаимном исключении территорий. По сведениям Maher (1974) на той же Аляске территории всех трёх видов непересекающиеся. Мною наблюдались частые межвидовые конфликты между средним и длиннохвостым поморниками, а также территориальное разграничение этих видов друг от друга в южной тундре Западной Сибири (Ryabitsev, 1977).
Существуют две основные теории касательно вопроса возникновения межвидовой территориальности.
По предположению Simons (1951) межвидовая территориальность — продукт ошибок в распознавании при агрессии, или взаимной, или преимущественно одного вида по отношению к другому сходному (теория «неадаптивной межвидовой территориальности»). Противоположная ей теория – теория Orians and Willson (1964) – основывается на правиле конкурентного исключения Гаузе (1934). Её следствием оказывается вывод что два вида, сходно использующих ресурсы, должны быть территориально разграничены (теория «адаптивной межвидовой территориальности»).
Развивая положения «адаптивной теории», Cody (1969, 1974) высказал «гипотезу конвергенции признаков». Согласно ей два или более экологически подобных вида в условиях симпатрии сближаются в плане проявлений межвидовой агрессии [и даже становятся внешне похожими, как он предполагал для дятлов — хотя неудачно], что и обеспечивает взаимное исключение территорий.
Обе концепции могут быть применены и к поморникам. Сходство внешнего облика этих видов крайне значимо. Можно предположить, что оно — следствие общего происхождения, и тогда взаимная агрессия будет неадаптивным следствием случайных ошибок. Тем более что ритуализированные демонстрации и, шире, видоспецифические паттерны поведения у разных поморников также схожи (Cramp and Simmons, 1983; Furness, 1987).
Некоторые аргументы поддерживают и «адаптивные теории», особенно связанные с конкуренцией за корм. Питание поморников достаточно полно изучено и описано (Maher, 1974; Dorogoy, 1981;Cramp and Simmons, 1983; Furness, 1987; Flint, 1988 etc.). В условиях тундры средний и длиннохвостый поморники питаются грызунами. Средний поморник в большей степени специализируется на леммингах; он не будет гнездиться в годы их депрессии, и широко перемещается в поисках территорий с обилием леммингов. Длиннохвостый поморник может гнездиться при низкой плотности леммингов, питаясь насекомыми и другими кормами; но лемминг и для него — основной корм.
Поэтому от среднего и длиннохвостого поморника можно ждать пространственной изоляции двух видов во избежание конкуренции за корм. Значимое несовпадение ареалов и разница гнездовых биотопов (Dorogoy, 1981; Furness, 1987) у этих видов могут быть следствием конкуренции между ними. Многие авторы считают, что короткохвостый поморник в тундре питается в основном птицами, однако наши наблюдения и работы других авторов показывают, что при обилии леммингов короткохвостый поморник охотно ловит и ест их. На полуострове Ямал, когда численность грызунов была высока, их останки найдены в 93-100% погадок поморников, тогда как останки птиц только в 8-22% (Osmolovskaya, 1948). То есть короткохвостый поморник может конкурировать со средним и длиннохвостым за корм. При уменьшении обилия грызунов короткохвостый поморник питается главным образом мелкими птицами; однако и другие виды поморников в нелемминговые годы переходят на яйца и птенцов разных видов птиц, т.ч. кормовая конкуренция всех трёх видов интенсивна.
Это будет благоприятствовать «адаптивной» территориальности. Поэтому в обсуждении механизмов межвидовой территориальности у поморников следует ответить на вопрос: способны поморники отличать конспецификов от особей других видов? Если птицы одинаково реагируют на захватчиков своего вида и двух других Stercorarius, то такая агрессия может быть механизмом межвидовой территориальности как по адаптивному, так и по неадаптивному сценариям.
Кроме уже обсуждённых причин, каждая пара поморников может гонять всех прочих — и своего и чужого вида — «на том основании», что они потенциальные хищники, угрожающие их яйцам и птенцам. Все поморники активно преследуют воздушных и наземных хищников, в т.ч. и прочих поморников. Агрессия к потенциальным хищникам [а не только к возможным территориальным захватчикам] также может быть механизмом территориального разделения видов, что надо проверить. Для ответа на все эти вопросы надо исследовать реальные взаимодействия между видами поморников при разной обеспеченности кормом и плотности расселения птиц.

Короткохвостый поморник. Светлая (1) и темная (2) морфы. Из: В.П.Шунтов. Трудная профессия альбатроса
Район исследования и население поморников
Представленные данные в основном получены на орнитологической станции «Яйбари» на севере полуострова Ямал (71°04′ с. ш., 72°20′ в. д.; Рис. 1), на южном пределе подзоны арктических тундр. Эта часть полуострова равнинна, как и вся Западная Сибирь. На изучаемом участке река Venuyieuoyaha характеризуется широкой поймой с болотами и озёрами. Местность за речной долиной равнинная или немного холмистая c мшистыми возвышенностями и мшисто-травянистыми болотами и озёрами, с ручьями текущими через довольно мелкие лощины (см. Рис. 4B).
Рис. 1. Район исследований. Чёрным кружком отмечено место положение полевой станции «Яйбари»
Растительность в основном представлена мхами, лишайниками, злаковыми и ползучими ивами. Кустарниковые ивы 0,1–0,3 м высотой растут преимущественно в нижней части речной долины. Здесь обитают два вида леммингов – основной корм питания поморников. Доминантный вид в этой части полуострова — сибирский лемминг Lemmus sibiricus. Копытный лемминг Dicrostonyx torquatus встречается реже. Полевки отмечены преимущественно в южной части района, одна из них — узкочерепная полёвка Microtus gregalis – редкий вид в окрестностях орнитологической станции. За время работы максимум численности грызунов приходился на 1988-1991 гг., когда преобладал сибирский лемминг.
Динамика населения поморников на станции «Яйбари» приводится в Таблице 1. Плотности разных видов сильно различны, также как направления их изменений. В 1992 году после очень поздней весны и на фоне глубокой депрессии леммингов поморники так и не приступили к гнездованию.

Средний поморник. Темная (1) и светлая (2) морфы. Из: Шунтов, op.cit.
Материалы и методика исследования
Статья базируется преимущественно на материалах, полученных близ станции «Яйбари» в 1988-1993 гг. Работу начинали с конца мая, когда птицы ещё не прилетели, а заканчивали — 5—12 августа, когда у поморников были уже большие или готовые к полёту птенцы. Найденные гнездовые пары и гнёзда отмечались на аэрофотоснимках пробной площадки в 25 км2. Обычно в сезон находили все или почти все гнёзда.
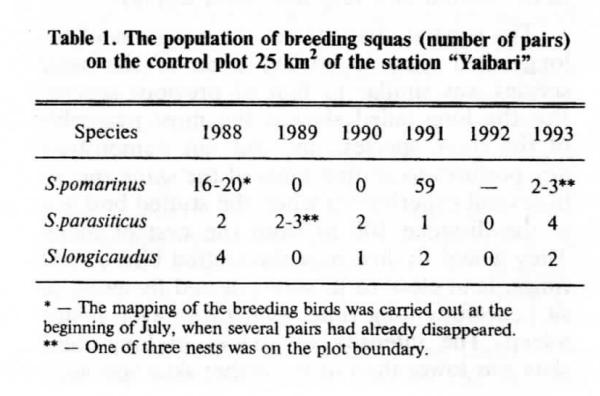
Погодовая динамика численности трёх видов поморников, число гнездящихся пар на контрольной площади в 25 кв.км. * — картирование гнездовых пар проводили в начале июля, когда некоторые из них уже исчезли. ** — одно из трёх гнёзд было на границе участка
С 1990 г. мы отлавливали поморников на гнёздах и метили каждую птицу обычным металлическим кольцом и цветным пластиковым кольцом. Также каждой пойманной птице делали 1-3 небольших высечки на маховых перьях. Указанные метки позволяли нам распознавать кольца с дистанции в 100-200 м, а птиц в полёте — с 300-600 м при использовании бинокля с 12-кратным увеличением. Также полезной для индивидуального распознания были особенности оперения, особенно у наиболее вариабельного среднего поморника с его довольно большим разнообразием в оперении.
Мною был составлен индивидуальный «портрет» каждого среднего и короткохвостого поморника на контрольной площади. Когда я становился свидетелем конфликта или любого другого взаимодействия между поморниками, я зарисовывал ритуализированные демонстрации особей, а сопровождающие их звуки транскрибировал словами.
Всем 3м видам поморников на гнездовые территории подставлялись чучела поморников — дважды за сезон (перед началом гнездования и во время инкубации), на расстоянии в 50-200 м от гнезда (Таблица 2), иной раз ближе или дальше (в Таблице 2 данные не представлены). Все чучела выполнены в позе мирно сидящей птицы. Я старался предъявлять чучело разным парам – чтобы исключить влияние индивидуального восприятия. К сожалению, в моём распоряжении было мало пар короткохвостого и длиннохвостого поморников и потому эксперименты проводились с одними и теми же парами, жившими в годы наблюдений на контрольной площади и не бывших за её пределами. Несколько экспериментов с чучелом было проведено после завершения инкубационного периода, когда уже были птенцы.
В ходе экспериментов с чучелом я зарисовывал ритуализированные демонстрации и транскрибировал сопровождающие их звуки. Наш гость из Великобритании Rose France помогла мне корректно записать звуки в английской транскрипции во время совместных опытов с чучелом летом 1993 года. В дополнение к основным данным за 1988–1993 гг. я использовал наблюдения за поморниками, сделанные в других районах Ямала с 1970 года.
Результаты
Внутривидовые демонстрации
Поморники используют различные звуковые сигналы для внутривидовой коммуникации, и в ответ на появление хищников (Cramp and Simmons, 1983; Flint, 1988). Мои наблюдения не вполне согласуются с написанным в указанных монографиях. В Таблице 3 описаны основные вокализации поморников. Обычно в природе мы можем видеть демонстрации, адресованные другому летящему поморнику своего вида. Они сопровождаются «долгим криком» (Long Call), который у каждого вида свой. Мои наброски этого воздушного обмена демонстрациями очень схожи с рисунками, приводимыми в работе R. Furness (1987).

Длиннохвостый (1) и большой (2) поморники. В.П.Шунтов, op.cit.
«Долгий крик» всегда демонстрируется при появлении незнакомой птицы своего вида на границе гнездовой территории. Эта демонстрация несёт три функции:
1) встреча потенциальных половых партнёров; в силу отсутствия полового диморфизма территориальный самец будет встречать всех конспецификов одной и той же демонстрацией;
2) «объявление» занятости территории; самцы без пары могут воспроизводить эту демонстрацию и без появления другой птицы на их участке; оба партнёра могут адресовать «долгий крик» захватчикам своего вида; иногда он появляется в погонях, сопровождающих охрану территории или адресуется особям, пролетающим над ней, особенно у среднего поморника при высокой плотности;
3) обычно наблюдается перед копуляциией;
«Долгий крик» намного реже издаётся на земле, чем в воздухе. Возможно поэтому рисунки наземного «долгого крика» сильней варьируют от публикации к публикации (Andersson, 1971; Cramp and Simmons, 1983; Furness, 1987). Наиболее обычная демонстрация — вертикальная (прямая) поза (Upright posture).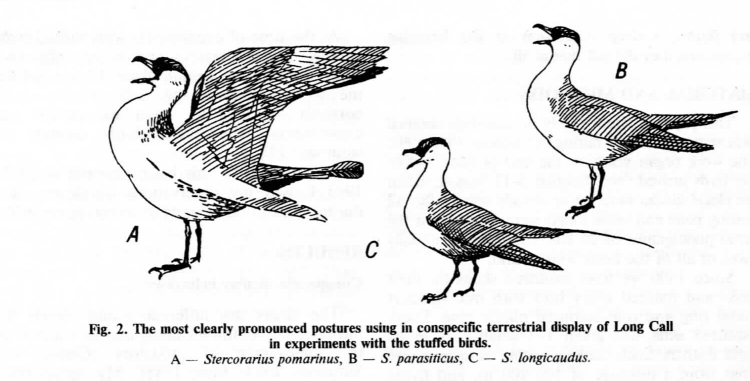
Рис. 2. Наиболее чётко проявляемые позы, используемые при внутривидовых демонстрациях «долгого крика» в экспериментах с чучелом. А – Stercorarius pomarinus, B – S.parasiticus, C – S.longicaudus
Я согласен с теми орнитологами (см. Furness, 1987), которые рассматривают эту позу как комплекс различных, но близких по форме элементов. Но в основном во время «долгого крика» принимается «наклонная» поза. На рис.2 эти позы показаны при наиболее ярком выражении, в опытах с чучелом. раскрытый хвост при сложенных крыльях — обычный, но не постоянный элемент «наклонной позы». Обычно он наблюдается не на начальной стадии исполнения «долгого крика», но после первого, ещё умеренного исполнения данной демонстрации (когда хвост ещё обычной ширины, и грудь не распушена до такой степени), когда чучело по понятной причине «не отвечает на неё» [почему это скорей всего чисто эмоциональная реакция, выражающая рост общего возбуждения вызванный «сбоем коммуникации», но семантически пустая, как сходные элементы в ритуализованной агрессии у большого поморника Cataracta skua, см. знаменитые работы Paton, Caryl, 1986; Paton, 1986]
Обычно поза «поклона» не наблюдалась на начальных этапах демонстрации «долгого крика», но так как чучело не осуществляло ответных демонстраций, то впоследствии указанная поза проявлялась в слабовыраженной форме (при которой хвост удерживается в нормальном положении, а грудные перья распушены не максимально).
Реакция на воздушных хищников
Все поморники активно защищают от воздушных хищников своё гнездо с яйцами или птенцами и прилегающую к нему территорию радиусом 100-400 м. В нашем регионе для поморников опасность с воздуха может исходить от мохноного канюка Buteo lagopus, белохвостого орлана Haliaeetus albicilla, полярной совы Nyctea scandiaca, болотной совы Asio flammeus и чаек Larus heuglini и Larus hyperboreus. Чем больше хищник, тем более интенсивны атаки и «крик атаки» (Attack Call) (Таблица 3). Наиболее выраженное поведение атаки — против орлана-белохвоста. В американской тундре поморники атакуют тех же или близких видов, а также канадских журавлей Grus canadensis (Pitelka et al., 1955). Во время атаки указанных птиц поморники пикируют на них и бьют. Такое поведение было отмечено у всех видов до появления кладок.
Результаты экспериментов с чучелами птиц
Реакция поморников на появление чучела особи своего вида
Во всех экспериментах, проведённых до гнездования и во время инкубации, короткохвостый и средний поморники реагировали на чучело своего вида «наклонной» демонстрацией, полностью выраженной (см. Рис. 2) или её слабой формой с добавлением «долгого крика». Затем они принимали «прямую» позу, а дальше — один или оба владельца территории (гнезда) клевали чучело в спину и шею или тащили за хвост. Если чучело не падало, оба хозяина или кто-то один взлетал и пикировал и бил его сверху. Чем ближе чучело размещали к гнезду, тем раньше хозяева переходили от демонстраций к непосредственной атаке. Атаки останавливаются при падении чучела.
Поморники адресуют «крик атаки» чучелу своего вида только при установке его рядом с птенцами [которых конспецифики могут стащить] или когда чучело закреплено на земле рядом с гнездом так, что не свалишь. Атаки не следовали после «долгого крика» несколько раз, когда опыт ставили на периферии охраняемой территории и на большом расстоянии от гнезда. В этой ситуации поморники довольно скоро теряют интерес к чучелу.
Общий характер реакций длиннохвостых поморников на чучела своего вида аналогичен поведению двух обсуждённых выше. Однако он самый «миролюбивый» (в смысле неагрессивный) из 3-х и потому не демонстрирует никаких поз чучелу, находящемуся на дистанции 100 м и далее. Он несколько раз низко пролетит над чучелом, сядет рядом, осмотрит, а затем просто улетит или же будет мирно сидеть рядом — долго, иногда даже засыпая. Интенсивность атак у длиннохвостого поморника ниже, чем у остальных видов.
Таблица 2. Количество предъявлений чучела и количество проверенных пар поморников (указаны в скобках)
|
Хозяин территории и гнезда |
Поставлено чучело: |
|||||
|
S.pomarinus |
S.parasiticus |
S.longicaudus |
||||
|
предгнезд. п. |
инкубац. п. |
предгнезд. п. |
инкубац. п. |
предгнезд. п. |
инкубац. п. |
|
|
S.pomarinus |
12 (6) |
8 (3) |
8 (5) |
6 (5) |
5 (2) |
7 (4) |
|
S.parasiticus |
– |
5 (2) |
5 (1) |
9 (3) |
– |
5 (1) |
|
S.longicaudus |
– |
5 (1) |
5 (1) |
5 (1) |
5 (2) |
12 (3) |
Таблица 3. Транскрипция издаваемых поморниками звуков
|
Вид |
«Долгий крик» |
Реакция на хищника |
|
|
Тревога |
Атака |
||
|
S.pomarinus |
«hyehhh-hyehh-hyehh-hyeh-hyeh-hyeh…» |
«hyev» или «hyav» |
«ay-ya-ya-ya-ya….» |
|
S.parasiticus |
«myahh, myahh, myahh…» |
спокойное «kyew» или «ke» |
«keh-goh» or «keh-yoh» |
|
S.longicaudus |
«kleh-kleh-kleh-kleeeyaaaa, kleeeyaaaa…» |
резкий «kreh» |
резкий «kreh», долгий приглушённый «kyaw» |
Реакция на чучело другого вида
Короткохвостый и длиннохвостый поморники выказывали агрессию по отношению к чучелу другого вида во всех проведённых опытах. Длиннохвостый поморник адресует «крик атаки» чучелам как среднего, так и короткохвостого поморника, короткохвостый поморник — лишь чучелу среднего. Средний поморник при нападении на чучела других, более мелких поморников, вообще не вокализировал [что однозначно говорит в пользу выражения в этих вокализациях мотивации предупреждения об опасности, но выражении ритуализированным образом, «своей» акустической формой у каждого вида, т.е. символически].
Птицы издавали «крик атаки» только при нападении на более крупных поморников и (также как при реакции на хищников) в случаях наиболее интенсивных атак. Атаки на чучело более мелкого вида поморника менее интенсивны и не сопровождались криками. Если хозяин гнезда слетал (с «криком атаки» или без него) к чучелу другого вида поморников и «не наблюдал ответа», то он либо бил чучело немедленно, или несколько раз низко летал прямо над ним и пикировал, прежде чем атаковать. Реакция хозяина более интенсивна при уменьшении дистанции чучела до гнезда. Иногда после запуска агрессивной реакции или нескольких облётов чучела чужого вида хозяин гнезда приземлялся рядом с ним в «прямой» или «наклонной» позе, сопровождаемых «долгим криком» [ошибка распознавания: отсутствие специфического ответа «хищника» на окрикивание, согласно стохастической модели инстинкта, с небольшой вероятностью, но переключит на восприятие чучела как «захватчика своего вида», т.е. на отпор территориальной агрессии, облик-то всех трёх видов сходен]. Как правило, средний поморник игнорировал чучело длиннохвостого поморника на расстоянии больше 50-100 м от гнезда.
Реакция поморников на представителей не своего вида в природе
При появлении чужаков на расстоянии 200-400 м от гнезда, кто-то из хозяев или оба сразу, как правило, слетали ему навстречу. Если это — представитель более крупного вида, то хозяева прямо сразу издают «крик атаки». В результате подобных преследований чужака таки отгоняли от ближайших окрестностей гнезда (100-400 м.). Если это более мелкий поморник, владельцы гнезда преследуют его молча. Они безразличны к поморникам других видов, пролетающих высоко над гнездом (свыше 100-150 м). Самая слабая реакция была у средних поморников на длиннохвостых: их атакуют только на расстоянии 50-100 м или менее. Я наблюдал несколько раз, как средний поморник встречал «долгим криком» короткохвостого, нарушившему границу его территории. И всякий раз владелец довольно быстро «видел ошибку» и прекращал демонстрации.
Предпочтительная среда обитания и влияние межвидового взаимодействия на распределение гнездовых территорий
На полуострове Ямал длиннохвостый поморник занимает пологие склоны лощин, довольно плоскую или (сильно реже) сырую тундру, но ни в коем случае не ещё более влажные болота. Короткохвостый гнездится на плоских и болотистых участках, как на возвышенностях, так и в пойменной тундре. Оба эти вида на Ямале не достигают высокой плотности, почему не конкурируют за места гнездования. Однако средний поморник влияет на расселение двух, более мелких видов. Он прилетает ранней весной, когда снег местами начинает таять и, при высокой численности леммингов, занимает гнездовые территории в широком спектре биотопов — от болот и крайне сырых тундр до сухих тундр в междуречьях. Он отчётливо избегает лишь большинства форм рельефа ниже уровня затопления в пойме, вершины холмов с голой землёй, песчаные отмели.
В 1988 г. численность средних поморников была высока (см. Таблицу 1), однако обе пары короткохвостого поморника загнездились на плоских болотах – самом предпочтительном месте для них; три пары длинохвостых поморников из четырёх гнездились на склонах лощин. В сезоны 1989 и 1990 гг. наблюдалась депрессия леммингов. Средний поморник тогда не гнездился на контрольной территории; длиннохвостый не гнездился в 1989 г., в 1990 г. одна пара гнездилась в плоской тундре; как и в 1988 г., короткохвостый гнездился на тех же территориях все эти годы.
Карта гнездовых территорий поморников в 1991 г., с высокой численностью леммингов (рис. 3) показывает влияние размещения средних поморников на гнездование других видов. Средний поморник занял практически всю плоскую тундру. Одна меченая пара короткохвостого поморника смогла загнездиться лишь на песчаной отмели с редкой травой, в 2 км от обычного места гнездования; вторая устроила гнездо на дне болотистой лощины с ручьём. Оба места нехарактерны для этого вида поморников на Ямале (Danilov et al., 1984). Одна окольцованная пара, в предыдущие годы гнездившаяся на большом болоте в западной части контрольной площадки, отмечена летом 1991 г., но гнезда не было найдено (возможно, в том году его и не было). Две пары длиннохвостого поморника в 1991 г. на площадке гнездились на сухих склонах лощин (Рис. 3).

Рис. 3. Размещение гнездовых территорий и гнёзд поморников и других видов птиц в 1991 г. 1 – Stercorarius pomarinus, 2 – S.parasiticus, 3 – S.longicaudus, 4 – Nyctea scandiaca, 5 – Asio flammeus, 6 – Buteo lagopus, 7 – поймы и болота.
В позднее лето 1992 г. леммингов не было, средний поморник не гнездился, хотя мечены длиннохвостые и короткохвостые наблюдались. Все три вида гнездились летом 1993 г. (см. Таблицу 1), когда численность леммингов возросла. Средних было лишь три пары, и понятно, что они не мешали другим гнездиться в своих предпочтительных местообитаниях. Все короткохвостые поморники заняли плоские сырые равнины; одна из двух пар длиннохвостого поморника (окольцованная в 1991 г.) гнездилась на верховом болоте, вторая пара – на краю болота, рядом с лощиной, где гнездилась в 1991 г.
Внутривидовая территориальность и район, защищаемый от хищников
Средний поморник строго территориален при высокой численности. Границы территории чётко «обозначены», и ей всегда соответствует район, в котором птицы атакуют воздушных хищников и беспокоятся при появлении человека (1991, Рис. 4А). При низкой плотности пространство, защищаемое от хищн6иков, существенно меньше территории. Но размер территории здесь не бывает таким большим, как у двух других видов.
Короткохвостый поморник распространён всегда спорадически: охраняемая территория очень большая, но территориальные взаимодействия «мягкие». Соседи хорошо знают друг друга, их встречи довольно миролюбивы по сравнению с другими видами, всегда решительно изгоняемыми. Структура территории оставалась практически постоянной из года в год и в течение всего сезона, даже летом 1991 г., когда сменилось место гнездования (Рис. 4В). Из всех поморников короткохвостый сильней всего привязан к выбранной территории; пара остаётся на участке и продолжает защищать даже после потери гнезда (Ryabitsev, 1993).
Характер внутривидовой территориальности у длиннохвостого поморника сходен с предыдущим видом, но есть небольшие отличия. Он более социален и менее связан с территорией в случае неудачного размножения. Границы территорий короткохвостого и длиннохвостого поморников указаны на Рис. 4 (В, С) пунктирном. На деле это не прямые линии [как у среднего поморника], но «нейтральные зоны», контролируемые обоими соседними парами, как и у ряда других видов (Ryabitsev, 1993). Может быть, самый интересный аспект межвидового взаимодействия между поморниками — то, что присутствие многочисленных средних году влияет на расселение длиннохвостых и короткохвостых, но не влияет на внутривидовую территориальность и на положение границ территорий у этих видов. Все внутривидовые взаимодействия между соседями, или между хозяевами и захватчиками, все демонстрации и преследования происходят обычно на высоте 50-100 м над землёй, и выше, чем с поморниками других видов.
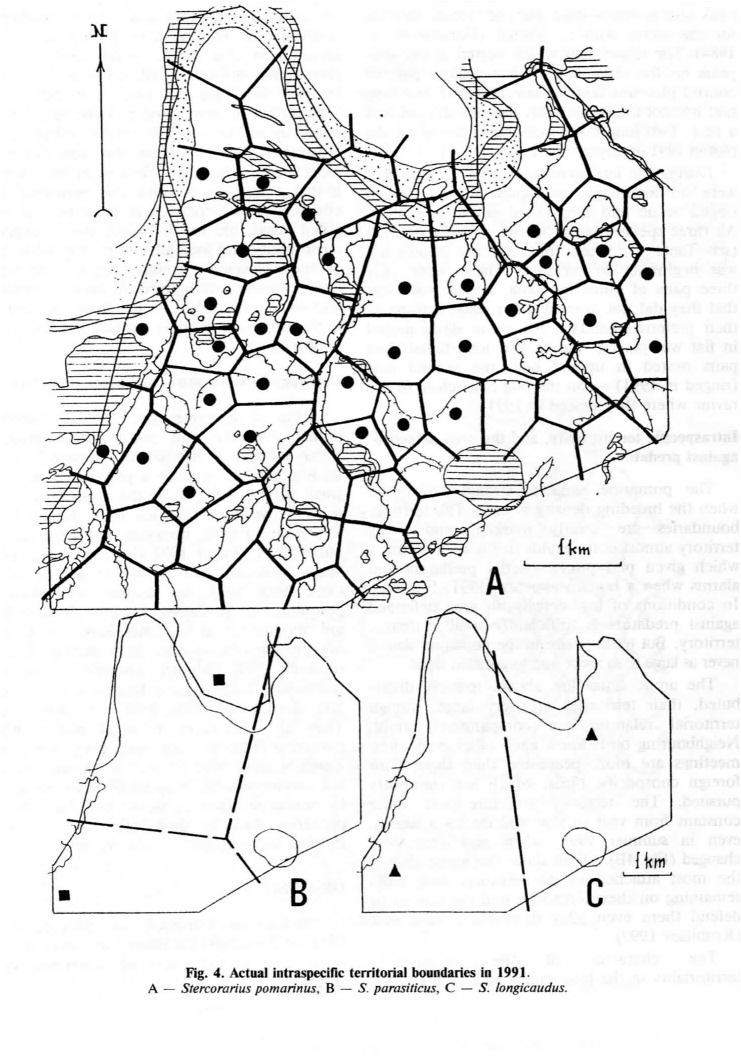
Рис.4. Действующие гнездовые территории трёх видов поморников в 1991 г.
Отношения между поморниками и другими хищниками
Все три вида поморников беспокоятся при появлении человека и пернатых хищников в 200-300 м, а в отдельных случаях и в 500 м от гнезда. Это лишь часть (как правило, малая) территории в обычном смысле этого термина (Нoward, 1920; Nice, 1941; Hinde, 1956). Распределение гнёзд сов, канюков и крупных чаек на контрольной площади в 1991 г. (при самой высокой численности лемминга) приводится на Рис. 3. Все эти виды реагируют друг на друга и на поморников как возможных разорителей гнёзд. Обычно численность этих видов мала и поэтому они не могут мешать друг другу в выборе мест гнездования. Однако летом 1991 г., несмотря на высокую численность среднего поморника, размножение мохноногого канюка, полярной и болотной сов также было очень успешным. Они гнездились на избегаемых средним поморником формах рельефа. Только одна пара чаек Larus heuglini пыталась обосноваться в болотистой тундре, но безуспешно – из-за непрестанных атак со стороны среднего поморника. Скорее всего, в сходной ситуации средний поморник убил белую сову на Аляске (Pitelka et al., 1955).
Обсуждение
Анализ звуков, используемых как при внутри- так и при межвидовых взаимодействиях, показал, что все виды поморников очень хорошо различают «своих» и «чужих»[ а позные демонстрации и вокализации строго видоспецифичны. Больше того — на видах-двойниках южнополярных поморниках рода Cataracta показано, что межвидовые отличия «долгого крика», здесь достаточно небольшие, остаются инвариантными независимо от индивидуальных вариаций в уровне возбуждения разных особей, который варьирует сильно.
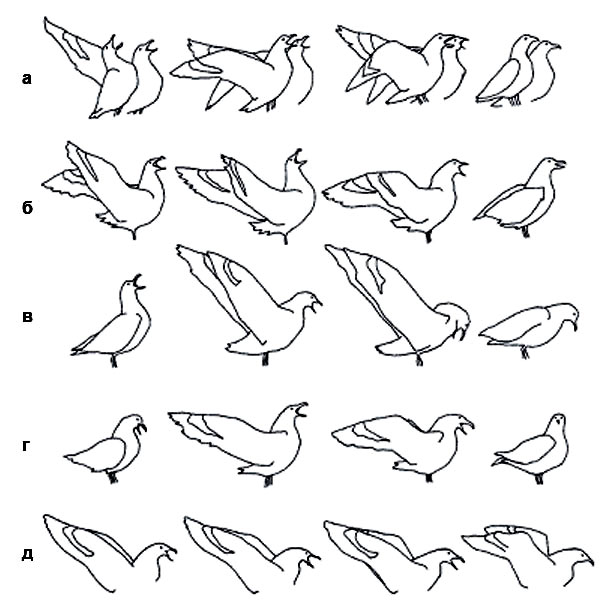
Долгий крик у видов-двойников антарктических поморников: южнополярного …
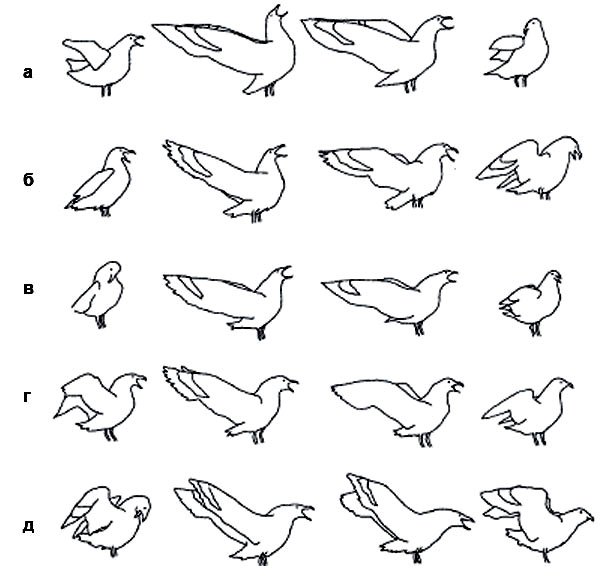
и бурого поморника (а—д — разные птицы, прорисовки по кадрам фильмов). Индивидуальная изменчивость не уничтожает общевидового облика демонстрации.
«Долгие крики» у обоих видов сходны, но между ними есть и четкие, устойчивые различия, как в целом, так и на каждом из пяти этапов: южнополярный поморник отклоняет корпус гораздо дальше назад и (или) вверх. Межиндивидуальная изменчивость «долгих криков» у обоих видов достаточно велика, но никогда не затрагивает видоспецифичных признаков. С другой стороны, стереотипность исполнения не ликвидирует и даже не уменьшает индивидуальную и ситуативную изменчивость — такое просто невозможно. Главное, что не изменяются дифференцирующие характеристики сигналов — те особенности, по которым различаются сигналы близких видов»
«От стимула к символу«. Ч.1.
Между «долгими криками» двух видов поморников сохраняются устойчивые различия: 1–5 — последовательные фазы долгого крика, столбики внизу — частота встречаемости у того и другого вида. (По:. Pietz P., «Condor», 1985, т. 81).

Поморники ведут себя различно в отношении своего вида и всех прочих. Ошибки распознавания редки и не приводят к проявлению «ошибочной агрессии». Эксперименты с чучелом проводились не во всех возможных вариантах и всего лишь с несколькими парами. Однако эти эксперименты убедительно подтверждают предположения, сделанные при наблюдениях за поведением птиц в естественной среде. В некоторых экспериментах демонстрация «долгого крика» и соответствующих поз адресовались особям других видов. Но нельзя утверждать, что эти демонстрации — следствие ошибки в распознавании, надо рассматривать их как побочный результат экспериментов. В первую очередь это реакция на «ненормальную» неподвижность чучела: оно «не реагирует» на беззащитность гнезда (т. к. хозяин улетел) и не стремится атаковать. Во-вторых, поморники, как и представители других видов, иногда издают отрывки песен или демонстрируют отдельные элементы поз, когда вдруг потревожены у или в любой другой критической или нестандартной ситуации (классическая «смещённая активность» – Tingerben, 1952).
Межвидовая агрессия поморников — это полностью защита гнезда от воздушного хищника (а не территориальная агрессия, как к нарушителям «своего» вида). Она точно такая же, как в отношении сов, канюков, крупных чаек и других потенциальных опасностей с воздуха. Таким образом, кажется бессмысленным обсуждать, является ли «межвидовая территориальность» поморников адаптивной или же нет, так как она не несёт характера защиты территории. Территории у разных видов поморников не исключают друг друга, но широко пересекаются, также как у неродственных видов птиц. «Овалы» на Рис. 3 — это не территории (охраняемое пространство от особей своего вида) а пространство, защищаемое от хищника. Это результат картирования мест, в которых появление орнитолога вызывает отчётливое беспокойство хозяев, т. е. «межвидовое территориальное разделение» не более чем ошибка (Pitelka et al., 1955; Maher ,1974; Ryabitsev, 1977).
Межвидовые отношения не влияют напрямую на распределение по биотопам и другие аспекты жизни трёх видов поморников – за исключением сезонов высокой численности среднего поморника; тогда в первую очередь длиннохвостый, а затем и короткохвостый поморники оттесняются в различные неудобные биотопы. Здесь средний поморник выступает в качестве действительного «тирана» для других видов этой экологической группы.
Возможно, что в большинстве случаев выбор длиннохвостым поморником склонов оврага и подобных биотопов — следствие не действительных предпочтений вида, но влияния среднего поморника – прямо или непрямо. Если молодой длиннохвостый поморник под влиянием среднего поморника загнездится на склоне оврага, она в следующие сезоны будет стремиться к таким же биотопам (запечатление местообитаний) и/или вернётся в то же самое место (привязанность к месту).
Трудно определить, как исторически сложилось гнездование полярной совы и мохноногого канюка на обрывах берегов и оврагов. Возможно, средний поморник был одним из факторов, определивших это. Будучи крупной, сильной и агрессивной птицей, не просто активно атакует любого хищника с воздуха, но при высокой численности становится им мощной помехой в корма, когда вся соответствующая местность «разбита» на охраняемые территории пар (и именно в самые обильные кормом годы). Теоретически возможно, что средний поморник может так узурпировать всю охотничью территорию других лемминофагов, что сократит их успех размножения. Однако мы имеем ясные примеры сосуществования всех лемминофагов: в 1991 г. три вида поморников, совы и канюки успешно размножались на выбранных территориях. Такая ситуация стала возможной благодаря высокой численности леммингов в 1991 г. В случае их низкой численности средний поморник либо не гнездится вовсе, либо делает это с малой плотностью расселения. При снижении численности лемминга они покидают свои гнёзда. Поэтому конкуренция за пищу между поморниками и другими хищниками, питающимися леммингами, должна быть весьма редкой.
У меня есть все основания полагать (Ryabitsev et al., 1980; Ryabitsev, 1993), что межвидовая территориальность несвойственна птицам Севера. Я изучаю территориальность у птиц Севера с 1971 г. и не обнаружил ни единого случая, подтверждающего гипотезу Cody о конвергенции признаков [в тропиках она тоже не работает]. Строгая экологическая изоляция невозможна в силу нестабильности и непредсказуемости погодных и кормовых условий, в силу несбалансированности трофических связей в тундровой экосистеме. Связи между поморниками тундры могут быть выявлены при дополнительных исследованиях в этой области.
Литература
Andersson M., Gotmark F. 1980. Social organization and foraging ecology in the Arctic Skua Stercorarius parasiticus: a test of the food defendability hypothesis// Oikos 35: 63-71.
Cody M.L. 1969. Convergent characteristics in sympatric species: a possible relation to interspecific competition and aggression// Condor 71: 223-239.
Cody M.L. 1974. Competition and structure of bird communities. Princeton, Univ. Press
Cramp S., Simmons K.E.L. 1983. The Birds of the Western Palearctic: Handbook of the Birds of the Europe, the Middle East and North Africa. Oxford, London, New York: Oxford Univ. Press. Vol. 3: Waders to Gulfs.
Danilov N.N., Ryzhanovski V.N., Ryabitsev V.K. 1984. Birds of Yamal. Moscow: Nauka (In Russian).
Dorogoy I.V. 1981. Ecology of skuas// Ecology of Mammals and Birds of Vrangel Island. Vladivostok: 38-55 (In Russian).
Hint V.E. 1988. Family Stercorariidae// Birds of the USSR: Gull Birds. Moscow; 10-47 (In Russian).
Furness R.W. 1987. The Skuas. Gallon: T. & A.D.Poyster.
Cause G.F. 1934. The Struggle for Exislance. Baltimore et al.
Hinde R.A. 1956. The biological significance of the territories of birds// Ibis 98: 340-369.
Howard H.E. 1920. Territory in Bird Life. London: Athene um.
Maher W.J. 1970. Ecology of Pomarine, Parasitic, and Long-tailed Jaegers, lake Hazen, Ellesmere Island// Arctic 23: 112-129.
Nice M. 1941. The role of territory in bird life// Amer. Midi. Nat. 26: 441-487.
Orians G.H., Willson M.F. 1964. Interspecific temtories in birds// Ecology 45: 736-745.
Osmolovskaya V.I. 1948. Ecology of the predator birds of Yama! Peninsula// Proc. Inst. Geogr. Acad. Sci. USSR 41: 5-77 (In Russian).
Pitelka F.A., Tomich P.Q., Treichel G.W. 1955. Ecological relations of jaegers and owls as lemming predators near Barrow, Alaska// Ecol. Monogr. 25: 85-117.
Ryabitsev V.K. 1977. Results of studies of interspecific territorial relations in birds on the Southern Yamal// Zoot. J. 56: 232-242 (In Russian).
Ryabitsev V.K. 1993. Territorial Relations nd Communities Dynamics of Birds in Subarctic. Ekaterinburg:Nauka (In Russian).
Ryabitsev V.K., Shutov S.V., Ol’shvang V.N. 1980. Analysis of concrete relations of the willow warbler and Eversmann’s warbler// Soviet J. Ecol. 11: 62-69.
Simmons K.E.L. 1951. Interspecific territorialism// Ibis 93: 407-413.
Tinbergen N. 1952. «Derived» activities: their causation, biological significance, origin, and emansipation during evolution// Qu. Rev. Bioi 27: 1-32.
Публикация статьи частично оплачена Институтом экологии растений и животных Уральского отделения Российской Академии наук.
Рекомендуем прочесть
 Используются ли «понятия» в коммуникации животных?
Используются ли «понятия» в коммуникации животных? Сигнальное поле как регулятор: от «памяти» и «наследственности» до эволюции
Сигнальное поле как регулятор: от «памяти» и «наследственности» до эволюции Разное про запаховую коммуникацию
Разное про запаховую коммуникацию «Жестикуляция» при совместной охоте коралловых рыб
«Жестикуляция» при совместной охоте коралловых рыб Ритуализация «по Благосклонову» и её сопряжение с «тинбергеновской»
Ритуализация «по Благосклонову» и её сопряжение с «тинбергеновской» «Человек» и «животное»: где провести грань?
«Человек» и «животное»: где провести грань? «Запасной ум» и «избыточная социальность» животных: проблемы реконструкции
«Запасной ум» и «избыточная социальность» животных: проблемы реконструкции Грань социального/несоциального у животных
Грань социального/несоциального у животных
