

Кудрявцева Н.Н., А.Л.Маркель, Ю.Л.Орлов
В статье дается обзор современной литературы, посвященной анализу генетической детерминации внутривидового агрессивного поведения – одной из наиболее сложных и универсальных форм социального поведения у животных, находящихся на различных ступенях эволюционной лестницы. Рассмотрены разные подходы к его изучению и перспективы, открывающиеся с появлением новых молекулярных технологий. Описывается комплексный подход к исследованию эволюционно-генетических и биологических корней агрессивности и ее проявлений при действии различных средовых и социальных факторов и при наличии определенных генетических предрасположенностей. Подчеркивается большая социальная значимость данной проблемы, решение многих аспектов которой связано с изучением генетико-физиологических механизмов агрессивного поведения как у людей, так и в эксперименте на животных. Обсуждается феноменология агрессии, ее стимулирующие и сдерживающие механизмы. Предполагается, что основным социальным механизмом, сдерживающим проявления агрессии в популяции, являются доминантно-субординантные отношения. Проанализированы нейробиологические детерминанты агрессии; представлено исследование роли наследственных факторов в контроле агрессивного поведения у животных. На примере нейрохимической перестройки мозга в процессе формирования агрессивного типа поведения у самцов мышей под влиянием повторного опыта агрессии в ежедневных межсамцовых конфронтациях показана модель исследования «от поведения – к гену». В обсуждении данных по генетике агрессивности у человека отмечено участие тех же генов, которые были выявлены в экспериментальных исследованиях на животных разных линий, а также нокаутных и трансгенных животных: генов транспортеров обратного захвата, рецепторов, ферментов биосинтеза и катаболизма серотонина и катехоламинов. В заключение рассмотрены перспективы использования транскриптомных исследований, вопросы эпигенетической регуляции агрессивного поведения.
Введение
Содержание
- 1 Введение
- 2 Феноменология агрессии
- 3 Нейробиологические детерминанты агрессии
- 4 Генетика агрессивного поведения: исследование роли наследственных факторов в контроле агрессивного поведения у животных
- 4.1 Межлинейные различия
- 4.2 Cелекция по отдельным формам агрессивного поведения
- 4.3 Изучение типа наследования при скрещивании контрастных по выраженности агрессивности особей
- 4.4 Нокаутные и трансгенные животные
- 4.5 Нейрохимическая перестройка мозга в процессе формирования агрессивного типа поведения у самцов мышей: от поведения – к гену
- 4.6 Генетика агрессивности человека
- 5 Исследование механизмов агрессивного поведения с помощью современных протеомно-транкриптомных технологий
- 6 Нейроинформатика поведения
- 7 Эпигенетическая регуляция агрессивного поведения
- 8 Заключение
Проявление агрессии и агрессивного поведения в человеческом обществе является одной из важнейших социальных проблем. Обычно в качестве причин агрессивных столкновений в человеческом обществе рассматриваются факторы социальной и общественно-политической жизни. Однако большую роль играют личностные характеристики людей, которые при определенных условиях могут приводить к сложной психологической борьбе, а иногда и к физическим столкновениям. Исследования последних десятилетий позволяют сделать заключение, что агрессивное поведение следует рассматривать как некую особенность поведения человека, в основе которой лежит генетическая и социально-биологическая история его становления как вида и как отдельной личности, а также история становления той социальной группы и среды, к которой он принадлежит.
Для исследования агрессивного поведения анализ должен распространяться не только на агрессивное поведение человека, но и на соответствующие формы поведения животных, что оправдывается взглядом на агрессию как на общебиологическое явление, имеющее глубокие эволюционные корни. Агрессия у животных является естественной эволюционно отработанной и адаптивной формой поведения. Агрессивное поведение позволяет животным существовать в условиях ограниченных ресурсов. Оно сопутствует конкуренции за пищу, за партнера при спаривании, за место обитания. У социальных животных агрессия является инструментом формирования иерархического сообщества. В результате доминантная особь имеет приоритет в доступе к пище и к половому партнеру (Lindenfors, Tullberg, 2011).
Таким образом, агрессивное поведение часто является предметом полового отбора. Формирование социальной иерархии предупреждает необходимость постоянных конкурентных агрессивных столкновений, которые чреваты физическими травмами и гибелью членов популяции. Можно полагать, что низкий уровень агрессивности может плохо отразиться на жизнеспособности и воспроизводстве популяции, в то же время высокая агрессивность также оказывает отрицательное воздействие, так как она мешает реализации других важных для выживания функций, например, таких, как добыча корма, воспитание потомства. Высокая агрессивность может быть причиной массового травматизма и гибели животных (Packer et al., 1995). Таким образом, можно полагать, что оптимальный уровень агрессивности у животных находится под контролем стабилизирующего отбора, что подтверждается исследованиями, проведенными на самых разных видах, например, таких, как водомерки (Eldakar, Gallup, 2011) или бабуины (Packer et al., 1995).
Наличие значительной генетической компоненты в детерминации агрессивного поведения, казалось бы, является многообещающим условием для успешного поиска специфических генов агрессивности. Однако такой поиск отнюдь не прост, тем более что получить ответы на главные вопросы по генетике агрессивности, раскрыть нейроэндокринные механизмы регуляции агрессивного поведения практически невозможно, если базироваться на исследованиях, проводимых только на человеке. В настоящем обзоре рассмотрены различные подходы к экспериментальному исследованию механизмов внутривидового агрессивного поведения у лабораторных животных и человека, а также перспективы, открывающиеся с использованием новых молекулярных технологий с акцентом на изучение роли в контроле агрессивного поведения как отдельных генов, так и средовых факторов, действие которых опосредуется через нейрофизиологические механизмы.
Феноменология агрессии
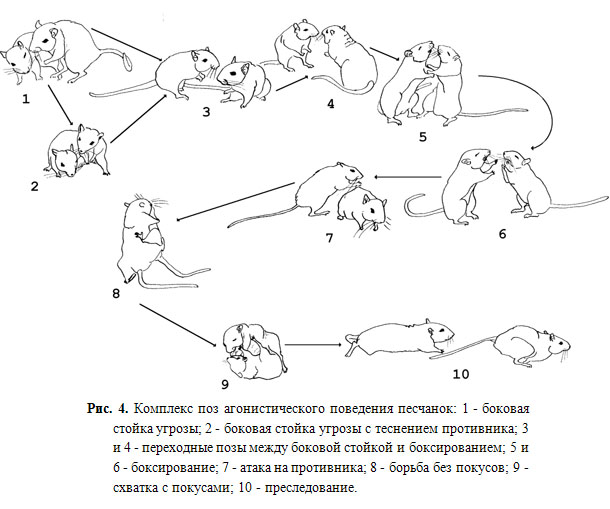
Агонистическое (конкурентное) поведение включает в себя проявления агрессии или подчинения особями, вступающими в конфликтные взаимодействия, и является одной из универсальных форм внутривидового поведения, встречающегося у животных, находящихся на разных ступенях эволюционной лестницы (Scott,1971; Лоренц, 1994; Blancard D.C., Blancard R.J., 2003). Оно демонстрируется самцами при формировании в популяции иерархических отношений или при защите территории, в борьбе за самку, пищу, среду обитания. Агрессия является врожденной формой поведения, развивающейся по инстинктивному типу в провоцирующих условиях среды в ответ на появление или предъявление видоспецифических сигналов. Однако и в природе, и в эксперименте внутривидовая агрессия в явном виде проявляется животными довольно редко в силу существования множества ингибирующих механизмов.
Основным фактором сдерживания внутривидовой агрессии в естественных и лабораторных популяциях являются установленные доминантно-субординантные отношения. Формирование подчиненного типа поведения происходит в результате опыта внутривидовых взаимодействий. Как правило, иерархию в популяции в значительной степени поддерживают подчиненные животные, избегая столкновений с сильным и агрессивным доминантным самцом. Средовое окружение, в котором разворачиваются события, формируя мотивацию поведения, определяет тот или иной тип агонистического внутривидового взаимодействия самцов. Различают территориальную агрессию, демонстрируемую при защите территории обитания, гнезда, потомства, а также агрессию, вызванную страхом, которая демонстрируется животными в ответ на нападение другой особи, когда животное вынуждено защищаться таким образом.
Выделяют также агрессию, возникающую в ответ на болевое воздействие, отсутствие чего-либо или вызванную неожиданными или неприятными условиями: это агрессия, вызванная раздражением или фрустрацией в конфликтной ситуации. Все эти виды агрессии в природе относят к поведению защиты. К агрессии нападения относится конкурентная агрессия, демонстрируемая самцами на общей территории проживания при установлении и/или перемене доминантно-субординантных отношений или при других обстоятельствах. Для характеристики агрессии у людей предложен ряд классификаций (Baron, Richardson, 1994). Исходя из наиболее признанного определения, агрессия – это любая форма поведения, направленная на оскорбление или ограничение возможностей («ущемление»), или причинение вреда любому другому существу, не желающему подобного обращения. Это не только прямая физическая, но и вербальная агрессия, наветы, интриги и т. д.
Агрессия у человека может быть спровоцирована внешними факторами и стимулами, и тогда ее называют импульсивной агрессией, которая рассматривается аналогом таковой у животных. Чрезмерная или патологическая агрессия является проблемой здравоохранения, поскольку повторный опыт агрессии отмечается в структуре многих психоэмоциональных и неврологических расстройств, таких как маниакально-депрессивный психоз, эпилепсия, шизофрения, токсикозы, заболевания мозга, аутизм. В феноменологии агрессивного поведения человека, его психологических и мотивационных составляющих отмечаются важные особенности, которые обусловлены сущностью природы человека и его социума. Независимо от стимула и мотивов, вызвавших проявление агрессивного поведения, механизмы его реализации в основном одинаковы.
В литературе выделяют влияние социального контекста (провокационная среда, территория, маркированная животным, противник), при котором учитываются социальные и средовые стимулы, влияющие на проявление агрессии, провоцирующие или тормозящие ее проявление. Социальными факторами, cтимулирующими агрессию у животных в природе, являются: перенаселенность1, недостаток пищи, половой дисбаланс, нестабильность климатических условий, наличие противника и несформированность доминантно-субординантных отношений. Основным социальным механизмом, сдерживающим проявления агрессии в популяции, являются доминантно-субординантные отношения, которые диктуют правила поведения животных в сообществе на основе полученного социального опыта, негативного (социальные поражения) или позитивного (достижение желаемого), а также наблюдения за поведением других сородичей.
Нейробиологические детерминанты агрессии
Этот аспект исследований направлен на изучение физиологических, нейрохимических и генетических особенностей регуляции поведения, эволюционно обусловленных характеристик, которые способствуют реагированию животного по агрессивному типу в определенных ситуациях. При этом особое внимание уделяется вопросу, почему в одном и том же средовом контексте у одной особи агрессия развивается, а у другой нет, и от чего это зависит. Литературные сведения указывают на множество эндогенных факторов, которые могут влиять на возникновение и характер агрессивного поведения особей, различающихся генетически, а также по психоэмоциональным и физиологическим характеристикам. При исследовании нейрофизиологических систем регуляции используют, как правило, животных одного генотипа. Эксперименты показывают, что повышение активности гамма-аминомасляной кислоты (ГАМК) и серотонергической систем различными методами может привести к снижению агрессии, в то время как активация катехоламинергических систем, наоборот, ее стимулирует (Miczek et al., 2007). Изменение гормонального фона также может как усилить демонстрацию агрессии, так и снизить ее проявление (Archer et al., 1998).
Таким образом, в рамках социо-биологического подхода главными детерминантами агрессии у животных, влияющими на ее проявления, независимо от видовой принадлежности, являются социальная среда, которая, создавая мотивацию, может стимулировать или тормозить проявление агрессии, и биологическая составляющая этого поведения, отвечающая за большую или меньшую генетическую предрасположенность индивида к агрессивной реакции в провоцирующих условиях.

Трубка, используемая для тестирования доминантного статуса самцов лабораторных мышей. см. подробнее рис.1 «Берем одного самца мыши и засовываем его в левое отверстие трубы, а другого — в правое. Самцы шагают навстречу друг другу и где-то по средине встречаются. В какой-то момент один из самцов начинает напирать (виннер), а другой пятиться (стало быть, лузер). Экспериментатор терпеливо ждет и все аккуратно записывает результаты в табличку.
Потом самцов можно генетически модифицировать и посмотреть, как у них потом дело с иерархией обстоит. Короче, у которого самчика проводимость нервных импульсов в синапсах пирамидальных нейронов префронтального кортекса выше, тот и в трубе наглее у того и социальный статус.
Хорошая новость (записывайте, девочки) - the hierarchical status of mice could be changed from dominant to subordinate, or vice versa, by manipulating the strength of synapses in the medial prefrontal cortex«. via progenes
Генетика агрессивного поведения: исследование роли наследственных факторов в контроле агрессивного поведения у животных
Высокие показатели наследуемости агрессивности установлены при исследовании разных видов животных, включая обезьян (Fairbanks et al., 2004), собак (Saetre et al., 2006), мышей (van Oortmerssen, Bakker, 1981), птиц (Drent et al., 2003) и людей (Hudziak et al., 2003), что говорит о том, что значительная доля фенотипической вариации уровня агрессивности в популяциях определяется генетическими факторами. Об этом свидетельствуют межлинейные различия в выраженности агрессивного поведения у лабораторных животных и быстрый эффект, достигаемый при селекции на повышенную и пониженную агрессивность.
Межлинейные различия
Как отмечено выше, о генетически детерминированном характере агрессивного поведения свидетельствует различная выраженность его у линейных животных в одних и тех же условиях. Инбредные линии мышей и крыс отличаются по выраженности агрессии в различных тестах, используемых для оценки агрессивного поведения: после длительной социальной изоляции, после раздражения током, в реакции нападения на пластинку в тесте трубы, а также при демонстрации агрессии в тесте «резидент–интрудер» (Brain, 1979; Куликов, Попова, 1980). Это указывает на влияние генетических факторов, которые участвуют в детерминации разных механизмов, регулирующих агрессивное поведение.
Исследования на мышах и крысах выявили различные варианты взаимосвязи между наследственно обусловленным уровнем тревожности и агрессивностью животного. При исследовании 10 инбредных линий мышей было показано, что самцы тех линий, которые показывали высокий уровень межсамцовой агрессии, имели и больший уровень тревожности, оцениваемый в тесте предпочтения темной или светлой камеры (Guillot, Chapoutier, 1996).
В другом исследовании, напротив, линии мышей (Swiss, CD1) с более высоким уровнем атак имели пониженный уровень тревожности, измеряемой в тесте крестообразного приподнятого лабиринта, по сравнению с менее агрессивными линиями CBA/2 и C57DL/6N (Parmigiani Parmigiani et al., 1999). В популяционных взаимодействиях высокоагрессивные доминанты проявляли более высокий уровень тревожности по сравнению с субординантными мышами (Ferrari et al., 1998). Можно предположить, что тревожность и агрессивность могут быть связаны «ортогональной зависимостью», показанной также и для людей (Barratt, 1972): есть линии животных с высоким уровнем тревожности и агрессивности, с низким уровнем тревожности и высокой агрессивностью, с высоким уровнем тревожности и низкой агрессивностью и низкими уровнями тревожности и агрессивности. Показано, что различия в характере агрессивного поведения мышей линии CBA/Lac с меньшей выраженностью агрессивного поведения и самцов мышей линии C57BL/6J с большей агрессивностью могут быть обусловлены различиями в эмоциональности, двигательной и исследовательской активности, болевой чувствительности, т. е. находиться под влиянием разных неспецифических составляющих их поведения (Кудрявцева, Ситников, 1987). Большое количество атак, наблюдаемое у особей линии C57BL/6J, может быть результатом более частых столкновений вследствие высокой двигательной и исследовательской активности этих мышей, в то время как у мышей линии CBA/Lac отмечены более низкая двигательная активность и агрессивность в новых условиях. На развитие агрессивного поведения у самцов линии CBA/Lac в большей степени может оказывать влияние выраженная эмоциональность, ингибирующая исследовательскую и двигательную активность (Walsh, Cummins, 1976). Взаимосвязанность проявления многих форм поведения и психоэмоциональных состояний животных, которые могут повлиять на характер агрессивного поведения, обнаружена и при селекции животных на выраженность агрессии.
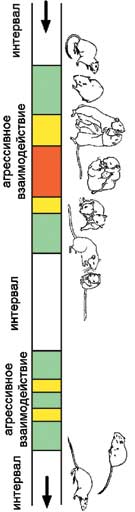
У всех трёх видов песчанок конфликты представляют собой чередование агрессивных взаимодействий и интервалов между ними. Источник Е.Володина. Грызуны в рукопашном бою
Cелекция по отдельным формам агрессивного поведения
Была проведена успешная селекция мышей на низкую и высокую выраженность агрессивного поведения, причем животные отбирались из генетически однородной популяции для того, чтобы получить различия именно в проявлении агрессии, а не других характеристик, влияющих на нее (Lagerspetz K.M.Y., Lagerspetz, K.Y.H., 1974). Аналогичные исследования по селекции животных на высокий и низкий уровень агрессивности были проведены и другими авторами (Beilharz R.G., Beilharz V.G., 1975; Van Oortmerssen, Bakker, 1981; Van Oortmerssen, Bakker, 1981; Cairns , 1981; Sandnabba,1986). Так, дикие домовые (не отличающиеся однородностью генотипа) мыши были селектированы на высокую и низкую агрессию (Benus et al., 1991). Агрессивные мыши с коротким латентным временем первой атаки (SAL – short latency attack) демонстрировали активную стратегию поведения, в то время как неагрессивные мыши (LAL – low latency attack) показывали пассивную стратегию поведения. Полагают, что эти различия в стратегии обусловлены разницей в регуляции гипоталамо-гипофизарно-надпочечниковой системы в условиях стресса (Veenema et al., 2005): низкоагрессивные мыши показали более высокое и продолжительное увеличение уровня кортикостерона в плазме, вызванное стрессом, по сравнению с агрессивными мышами. Межлинейные различия в проявлении агрессивного поведения также достоверно коррелировали с двигательной активностью мышей (Soutwick, Clark, 1968), хотя авторы отмечают исключения для некоторых линий. Обычно селекция мышей на выраженность агрессии приводила к повышению двигательной активности (Lagerspetz, 1969). Складывается впечатление, что драки и активность контролируются одними и теми же факторами (Хайнд, 1975). Считается, что двигательная активность отражает степень возбуждения нервной системы (Крушинский, 1960). Это позволило предположить у агрессивных животных большую возбудимость нервных процессов (Goldsmith et al., 1978; Вальдман, Пошивалов, 1984).
Увеличение интенсивности агрессии связывали как со снижением порога ответа центральной нервной системы на стимуляцию, так и с оптимизацией уровня общей нервной активности, необходимой для осуществления интегрированного поведения. В современных терминах это могло бы называться импульсивностью, которая, по мнению многих исследователей, является необходимой, хотя и далеко не достаточной характеристикой для проявления агрессии. Эмоциональность, которая чаще всего оценивалась в тесте «открытое поле», понимаемая как поведенческая и физиологическая реакция организма на новизну и сопровождаемая развитием страха, находится в обратной зависимости с проявлениями агрессивности у животных (Brain, Nowell, 1969). Мыши, прошедшие селекцию на более высокий уровень агрессивности, имели меньшие показатели эмоциональности, чем низкоагрессивные самцы (Lagerspetz K.M.J., Lagerspetz K.Y.H., 1974).
И, наоборот, селекция крыс на высокую эмоциональную реактивность сопровождалась снижением уровня агрессивности (Annen, Fujita, 1983). Таким образом, можно думать, что эмоциональность влияет на выраженность агрессии у животных, особенно в стрессирующих условиях. Путем многолетней селекции диких серых крыс (Rattus norvegicus), проводимой в Институте цитологии и генетики СО РАН, были получены линии животных, значительно отличающихся по выраженности агрессивного поведения в ответ на приближение человека (тест на перчатку) (Belyaev, Borodin,1982; Плюснина, Оськина, 2000; Плюснина, Соловьева, 2010).
Признак, по которому велась селекция крыс-пасюков, рассматривается как агрессия, вызванная страхом, т. е. как активная защитная реакция (Moyer, 1968). Однако в результате были получены агрессивные крысы, демонстрирующие высокий уровень агрессии не только по отношению к человеку, но и по отношению к партнеру в зоосоциальных взаимодействиях (межсамцовая агрессия, территориальная агрессия и др.) (Плюснина, Соловьева, 2010).
Кроме того, путем селекции были получены «ручные» неагрессивные особи, которые были толерантны или даже «дружелюбны» по отношению к человеку (Belyaev, Borodin, 1982; Плюснина, Оськина, 2000). Показано, что ручные и агрессивные крысы отличаются друг от друга по многим поведенческим и физиологическим характеристикам (Плюснина, Оськина, 2000; Popova et al., 2005; Albert et al., 2008; Plusnina et al., 2008). Таким образом, и в этом случае было показано, что генетическая селекция по одному из параметров агрессивного поведения сопровождается изменением многих нейрофизиологических характеристик животных.
Изучение типа наследования при скрещивании контрастных по выраженности агрессивности особей
Ранние попытки определения типа наследования агрессивности животных характеризуются широким разнообразием результатов. При скрещивании линий, контрастных по агрессивности, у потомков первого поколения обнаружены в одних случаях доминирование низкого (Сiaranello et al., 1974), в других – высокого (Southwick,1970; Kessler et al., 1977) уровня агрессивности, а также наличие аддитивного характера наследования показателей агрессивности (Куликов, Попова, 1980). Во многих исследованиях обнаружены различия между реципрокными гибридами (Roubertoux, Carlier,1988).Были получены данные о влиянии Y-хромосомы на проявление межсамцовой агрессии (Stewart et al., 1980; Maxson et al., 1989). В исследованиях Ciaranello с соавт. (1974) и Kessler с соавт. (1977) отсутствие реципрокного эффекта на проявление агрессии при скрещивании линий мышей, отличающихся по этому признаку, а также характер расщепления его у потомков второго поколения позволили авторам прийти к выводу о моногенном аутосомном контроле поведения нападения («атака»). Другой генетический подход с анализом реципрокных гибридов и рекомбинантных линий обнаружил, что в контроль агрессивного поведения включаются, по крайней мере, два аутосомных локуса, а также материнский эффект (Elefteriou et al., 1974).
При скрещивании двух близкородственных агрессивных и двух близкородственных неагрессивных линий мышей показано, что агрессивность находится под генетическим контролем 4 локусов, два из которых усиливают проявление агрессивности, а два других – подавляют, причем один из локусов, усиливающий агрессивность, находится в Y-хромосоме, остальные – на аутосомах (Selmanoff et al., 1976). По-видимому, разнообразие данных по типам наследования агрессивности обусловлено, в первую очередь, характерным сочетанием родительских линий мышей, взятых для скрещивания и несущих разные генетические пулы, участвующие в определении агрессивности. При диаллельном анализе параметров агрессии, вызванной изоляцией, показано, что латентное время первой атаки, интенсивность и время атак у гибридов первого поколения могут наследоваться как сходно, так и по-разному в зависимости от сочетания родительских линий при скрещивании (Hahn, Haber, 1982), что говорит о множественности генетических факторов, которые их определяют.
В обзорах последнего времени, посвященных анализу данных по генетике агрессивного поведения (Craig, Halton, 2009; Anholt, Mackay, 2012; Pavlov et al., 2012), подчеркивается вывод о многолокусной детерминации агрессивного поведения животных и человека. В косвенной детерминации агрессии могут участвовать гены, вовлеченные в регуляцию других видов поведения: полового, исследовательского, тревожности, других эмоциональных состояний и стресс-реактивности. В настоящее время генетический анализ агрессивности мышей проводится путем поиска локусов количественных признаков (QTL). Так, геномы контрастных по агрессивности линий маркируются множеством полиморфных маркеров, линия агрессивных мышей (например, BALB/cJ) скрещивается с неагрессивной линией A/j и анализ ассоциаций маркированных локусов с агрессивностью проводится на расщепляющихся гибридных популяциях (Dow et al., 2011). Однако этот подход позволяет оперировать лишь с некоторой частью возможной генетической и фенотипической изменчивости, которая ограничена разницей между выбранными родительскими линиями. Бывает трудно установить гены, связанные с агрессивностью и находящиеся в обнаруженном районе хромосомы – локусе количественного признака (QTL).
Протестировать каждый из множества имеющихся в локусе генов часто не представляется возможным. Многообещающим выглядит подход, предусматривающий создание панели рекомбинантных инбредных линий, полученных на основе гибридов от скрещивания нескольких разных линий мышей (Churchill et al., 2004), что должно, казалось бы, обеспечить сужение районов поиска генов на хромосомах (Aylor et al., 2011). Однако пока не удалось найти гены агрессивности таким способом (Mathes et al., 2011).
Нокаутные и трансгенные животные
В дополнение к исследованиям по поиску QTL перспективы изучения роли генов в контроле различных форм поведения вообще и агрессивного, в частности, в последние годы связывались с успехами в области молекулярных технологий, которые позволяют получать ген-модифицированных животных, например, с выключением (нокаутные мыши) или инсерцией (трансгенные мыши) определенных генов, вовлеченных в функционирование нейромедиаторных систем (обзоры: Haller, Kruk, 2006; Miczek et al., 2007), регулирующих агрессивное поведение (Nelson, Trainor, 2007). У мышей обнаружены мутации, с которыми связывают формирование гиперагрессивного поведения. Спонтанно возникающая мутация гена ядерного рецептора NR2E1, получившая название «ярость» (fierce), приводит к гиперагрессивности (Young et al., 2002a), которая может быть подавлена замещением мутантного гена нормальным человеческим гомологом (Abrahams et al., 2005). Также выключение гена нейрональной NO-синтазы приводит к тому, что мыши NO-синтазы приводит к тому, что мыши начинают нападать даже на партнеров, которые демонстрируют полное подчинение и нежелание продолжать схватку. Эти мыши-агрессоры убивают своих сожителей по клетке (Nelson et al., 1995). Интересно, что материнская агрессия по отношению к интрудеру-самцу у таких мышей подавлена (Gammie, Nelson, 1999), в то время как у мышей с повышенной материнской агрессией экспрессия нейрональной NO-синтазы повышена (Gammie et al., 2007). Высказано предположение, что изменение агрессивного поведения при нокауте гена нейрональной NO-синтазы связано с нарушениями серотонергической нейротрансмиссии (Chiavegatto et al., 2001). В случаях с мутациями отдельных генов зачастую выясняется, что эффект этих мутаций зависит от генетического фона.
Так, эффект нокаута гена нейрональной NO-синтазы, который выражается в гиперагрессивном поведении, ослабляется при переносе данной мутации с генотипа мышей C57BL/6J-129 на генотип мышей C57BL/6J (Leroy et al., 2000). Также действие мутации «ярость» значительно ослабляется при переносе ее с генотипа мышей C57BL/6J на B6129F1 (Young et al., 2002a). Эти факты свидетельствуют о наличии геновмодуляторов, которые могут серьезно изменять действие аллелей агрессивности. Манипуляции с отдельными генами подтверждают мнение о том, что агрессивное поведение как фенотипическая характеристика имеет сложную генетическую основу, которая представлена ансамблем взаимодействующих генов. Проявление агрессивного поведения тесно связано с особенностями функционирования серотониновой (5-НТ) системы мозга и, в частности, с состоянием 5-НТ рецепторов.
 Исследования мышей с нокаутом по гену рецепторов серотонина 5-HT 1A выявили снижение агрессивности у мутантных мышей по сравнению с мышами дикого типа (Bell, Hobson, 1993). Мыши с нокаутом по гену серотонинового рецептора 5НТ1В отличаются повышенной агрессивностью (Saudou et al., 1994). При выключении гена MAOA, фермента, разрушающего серотонин, уровень агрессии был повышен на фоне увеличенного уровня серотонина (Cases et al., 1995; Scott et al., 2008; Popova et al., 2001). Нокаутные мыши по гену серотонинового транспортера (Net), обеспечивающего обратный нейрональный захват нейромедиатора из синаптической щели, имеют сниженный уровень агрессии и увеличенный синтез серотонина (Holmes et al., 2002; Kim et al., 2005; Lewejohann et al., 2010).
Исследования мышей с нокаутом по гену рецепторов серотонина 5-HT 1A выявили снижение агрессивности у мутантных мышей по сравнению с мышами дикого типа (Bell, Hobson, 1993). Мыши с нокаутом по гену серотонинового рецептора 5НТ1В отличаются повышенной агрессивностью (Saudou et al., 1994). При выключении гена MAOA, фермента, разрушающего серотонин, уровень агрессии был повышен на фоне увеличенного уровня серотонина (Cases et al., 1995; Scott et al., 2008; Popova et al., 2001). Нокаутные мыши по гену серотонинового транспортера (Net), обеспечивающего обратный нейрональный захват нейромедиатора из синаптической щели, имеют сниженный уровень агрессии и увеличенный синтез серотонина (Holmes et al., 2002; Kim et al., 2005; Lewejohann et al., 2010).
Исследования мышей-нокаутов по генам, ответственным за метаболизм норадреналина, не подтверждают и не отрицают возможность специфического участия норадреналина в механизмах агрессивного поведения. У мышей-нокаутов по гену норадреналинового транспортера (Net) или по гену Мao-а, кодирующему фермент катаболизма катехоламинов отмечены повышенный уровень норадреналина и повышение агрессии по сравнению с диким типом (Сases et al., 1995; Haller et al., 2002). У мышей с выключенным геном дофаминбета-гидроксилазы, фермента биосинтеза норадреналина, отмечено снижение как уровня норадреналина, так и агрессии (Marino et al., 2005). Мыши, нокаутные по гену норадреналинового рецептора альфа-2c, быстрее атаковали интрудера, чем мыши дикого типа, в то время как мыши с повышенной экспрессией этого рецептора имели противоположный поведенческий фенотип (Sallinen et al., 1998). У нокаутов по гену дофаминового транспортера (Dat1) наблюдали повышенный уровень дофамина и усиление агрессивного поведения (Rodriguiz et al., 2004). Нокауты по гену катехол-О-метил трансферазы (Comt), фермента катаболизма катехоламинов, демонстрировали повышенную агрессивность (Gogos et al., 1998).
Имеется много данных о влиянии делеций других генов на агрессивность животных (обзоры: Nelson, Chiavegatto, 2000; Miczek et al., 2007). Агрессия, как и все виды поведения, пластична, и ее проявления зависят от внешних условий. Доказано наличие взаимодействия генотипа и среды на проявление агрессии. Так, макаки, которые воспитывались в неблагоприятных социальных условиях, были более агрессивны, особенно если у них был аллель гена Mao-A, обусловливающий низкую активность фермента катаболизма катехоламинов – моноаминоксидазы типа А (Karere et al., 2009). Внешние факторы могут оказывать влияние на агрессивность также через гормональную систему, включенную в регуляцию стрессорной реакции и репродуктивной функции. Эти внешние стимулы зачастую являются феромонами, действующими на рецепторы вомероназальной системы, которая у животных является рецептивным органом, воспринимающим информацию о родственных связях, сексуальных отношениях, наконец, о близости хищников (Tirindelli et al., 2009; Mucignat-Caretta, 2010; Isogai et al., 2011).
Не умаляя успехов в выяснении роли отдельных генов в регуляции сложных форм поведения при использовании нокаутных животных, все же хотелось бы отметить искусственность изменений, которые возникают в их организме.
Интерпретация различий в поведении, обусловленных искусственными генетическими мутациями, может быть ошибочна, поскольку в процессе индивидуального развития происходит компенсаторное изменение функциональной активности многих нейротрансмиттерных систем, что неизбежно должно повлиять и на характер разных форм индивидуального и социального поведения, в том числе агрессивного. Можно предположить, что использование ген-модифицированных животных для изучения роли отдельных генов и медиаторных систем в естественных механизмах поведения вряд ли станет продуктивным методом, поскольку выключение гена, скорее всего, приводит к развитию «патологического мозга», причем с изменениями, зачастую далекими от тех, которые возникают при различных мультифакториальных патологиях.
Нейрохимическая перестройка мозга в процессе формирования агрессивного типа поведения у самцов мышей: от поведения – к гену
Хорошо известно, что исход межсамцовых агонистических взаимодействий в большой степени определяется предшествующим социальным опытом. Победившее в драке животное более агрессивно в следующем столкновении с незнакомым партнером, а агрессивность мышей, терпящих поражение, наоборот, подавляется (Scott, 1966; Lagerspetz K.M.J. Lagerspetz K.Y.H., 1974; Cairns et al., 1983). Постоянный опыт побед использовался в экспериментах для воспитания агрессивных самцов-бойцов (обзор Scott, 1971), при этом агрессивность особей увеличивалась после одержанной победы, и этот феномен проявлялся у мышей как высоко-, так и низкоагрессивной линий (Cairns et al., 1983). В свою очередь, побежденные мыши хуже тренируются на повышение агрессивности, и с каждым последующим поражением подчиненное поведение становится все более выраженным и стереотипным, причем оно демонстрируется по отношению не только к агрессивному партнеру, но и к неагрессивным особям (Scott, 1966; Leshner et al., 1980; Пошивалов, 1986). Показано, что ингибирующие эффекты однократного поражения сохраняются в течение нескольких дней (Charpentier, 1969), а некоторые параметры поведения (замирание) проявляются и через десятки дней при столкновении даже с неагрессивным партнером (Scholtens, van de Pool, 1987).
[Также у победителей происходят структурные изменения гиппокампа и миндалины. «В эксперименте ученых пару самцов сажали в клетку, разделенную сеткой на две части. Мыши могли видеть, слышать и обонять друг друга, но не могли пустить в ход зубы. Каждый день после обеда сетку убирали, после чего немедленно начиналась драка. Через несколько минут выявлялся победитель и мышей снова отделяли друг от друга. Такие эксперименты ученые повторяли несколько дней, а потом перемешивали грызунов так, чтобы в одной клетке не оказалось двух побежденных или победителей. Через три недели ротации мышей разделяли на две группы: в одной триумфаторов отстраняли от стычек, а в другой грызунов продолжали сталкивать друг с другом.
В параллельных экспериментах ученые оценивали уровень агрессии животных (их помещали в клетку с прозрачной перегородкой и засекали время, которое испытуемые проводили у барьера) и его влияние на поведение: мышей сажали в крестообразный лабиринт, в котором один коридор был закрыт, а второй представлял собой открытую площадку. Чем больше времени мыши предпочитали отсиживаться в темном закрытом пространстве, тем больше их поведение можно было охарактеризовать словами «избегающий риска».
Все тесты показали, что опытные самцы, победившие в нескольких драках, ведут себя более агрессивно. Они чаще подходят к прозрачной загородке и скорее нападают на соперника после поднятия перегородки-сетки. Если же таким мышам не давали драться какое-то время до опыта, они становились еще агрессивнее: время перед атакой уменьшалось почти втрое, а сами драки продолжались дольше. Кроме того, у тех же доминирующих животных одновременно росла тревожность. Самец, уверенно победивший в нескольких драках, в лабиринте избегал открытого пространства и предпочитал по возможности отсиживаться в темноте.
В эксперименте ученые исследовали влияние агрессии на структуру гиппокампа и миндалины — зон мозга, связанных с памятью и эмоциями, соответственно (при этом миндалина может быть вовлечена и в некоторые аспекты памяти — благодаря ей, например, мышь замирает на том месте, где раньше ее ударило током). Для этого ученые отслеживали уровень активации гена c-fos, который начинает работать перед любыми долговременными перестройками в мозге, например при обучении.
Оказалось, что у победивших животных не только росла агрессия, но еще и увеличивалось количество новых клеток гиппокампа, а при возможности продолжать драки менялась активность уже существующих клеток — в них росло количества белка, синтезируемого с гена c-fos. В миндалине же активность c-fos, наоборот, падала. Такие эффекты повторились на четырех разных линиях подопытных мышей, что исключает случайное совпадение и говорит о связи между изменениями в структуре мозга и ростом агрессии вместе с тревожностью.
Результаты работы, в которой участвовали исследователи из Института цитологии и генетики РАН, МФТИ, Лаборатории Колд Спринг Харбор, университета и школы медицины Стони Брук,, опубликованы в журнале Frontiers in Neuroscience».
Источник Чердак
Что хорошо согласуется с этологическими данными о поведении доминантов, действующих по принципу «сила есть — ума не надо», способности подчинённых особей их обойти за счёт «большего ума», в том числе восприятия новых объектов, ситуаций и взаимодействий как вызывающих интерес и исследование, а не тревогу, после чего пользующихся их результатами, чтобы «обводить доминантов вокруг пальца» или как минимум реже встречаться с ним, и «цене» доминаирования, повышенной у социальных видов прямо пропорционально статусу и агрессивности альф. Прим.публикатора]
Последовательный опыт поражений приводит к формированию субмиссивного поведения, которое существенно меняет состояние особей (Raab et al., 1986). Полагают, что именно социальное давление – опыт повторяющихся социальных поражений при столкновении с доминантной особью – является причиной, хотя и не единственной, того, что групповые животные демонстрируют низкую агрессивность по сравнению с изолированными (Cairns et al., 1983). Таким образом, существенным является опыт социального обучения, в результате которого у индивида (особи) формируется представление о возможных негативных и позитивных последствиях агонистического взаимодействия и который во многом может предопределить его дальнейшее поведение. Этот принцип был использован в модели сенсорного контакта (Kudryavtseva,1991), позволяющей формировать у животных агрессивный тип поведения под влиянием повторного опыта побед в ежедневных конфронтациях. Основным методическим приемом в этой модели было постоянное проживание партнеров в условиях сенсорного контакта. Для этого пары самцов мышей одной и той же линии постоянно содержат в небольшой экспериментальной клетке, разделенной прозрачной перегородкой с отверстиями, позволяющей им видеть, слышать и воспринимать запахи друг друга. Один раз в день перегородка убирается для инициации «социального конфликта». В ежедневных конфронтациях с представителем противоположного социального опыта у половины самцов формируется агрессивный тип поведения, у другой половины – субмиссивный. Подробное описание всех этапов методики приводится в нескольких обзорах (Kudryavtseva, 2011; Кудрявцева,2012).
Вскоре было замечено, что повторный опыт агрессии, как и повторный опыт социальных поражений, специфически изменяет многие формы социального и индивидуального поведения в различных ситуациях, а также психоэмоциональное состояние самцов мышей. Ценным оказалось пришедшее понимание того, что длительный позитивный и негативный социальный опыт в условиях хронического конфликта может приводить к развитию различных психоэмоциональных и психоневрологических заболеваний. Среди них экспериментально было подтверждено формирование под влиянием хронического опыта социальных неудач депрессивно-подобного состояния, психогенного иммунодефицита, генерализованной тревожности и др. (Devoino et al., 1993; Kudryavtseva, Avgustinovich, 1998; Августинович и др., 2004; Тендитник и др., 2004).
У самцов с повторным опытом «побед» развивались патологическая неконтролируемая агрессия, злобность, повышенная тревожность, раздражительность, нарушение мотивационного поведения, социального распознавания, а также появление неврологической симптоматики: гиперактивности, дефицита внимания, стереотипий и т. д. (Kudryavtseva, 2006). В первых исследованиях у агрессивных самцов были найдены изменения медиаторной активности мозга на уровне метаболизма, активности ферментов, рецепции в разных отделах мозга, возникающие под влиянием повторного опыта агрессии, сопровождавшегося победами и формирующие дисбаланс в работе медиаторных систем – активации дофаминергической и ингибиции серотонергической систем головного мозга (Kudryavtseva, 2006). Возникло предположение, что эти изменения в моноаминергической активности мозга должны сопровождаться изменением экспрессии определенных генов в структурах мозга как у самцов с повторным опытом агрессии, так и у особей с повторным опытом социальных поражений, что и было выявлено в последующих экспериментах. Так, у агрессивных самцов на фоне активированных дофаминергических систем было найдено увеличение экспрессии дофаминергических генов (Th, Dat1, Snca) в вентральной тегментальной области, содержащей большие скопления дофаминергических нейронов, причем изменения в экспрессии генов тирозингидроксилазы (Th) и транспортера дофамина (Dat1) длительно сохранялись (Кудрявцева, Бакштановская, 1991; Bondar et al., 2009). Была также продемонстрирована корреляционная связь между выраженностью агрессивного поведения и экспрессией дофаминергических генов у агрессивных самцов мышей.
В то же время у этих самцов происходило снижение уровня экспрессии серотонергических генов: транспортера серотонина (Sert), триптофан гидроксилазы (Tph2), моноаминоксидазы (Mao-а) – в ядрах шва среднего мозга, который после периода прекращений конфронтаций возвращался к нормальному состоянию (Smagin et al., 2013). Помимо специфических функциональных генов найдено снижение экспрессии нейротрофического фактора мозга (Bdnf) (Boyarskikh et al., 2013; Smagin et al., 2013). Эти данные свидетельствовали о принципиальной возможности исследовать функциональную или, другими словами, физиологическую регуляцию экспрессии генов у животных с агрессивным или субмиссивным типами поведения, нейрохимическая и гормональная регуляция которых достаточно хорошо изучена. Исследования показали, что существуют динамика и последовательность нейрохимических и молекулярных событий, вовлекающих на определенном этапе конкретные гены в процесс работы медиаторных систем мозга. Создавая экспериментальные условия, при которых меняется тип поведения (инверсия поведения), можно предсказуемо изменять экспрессию определенных генов в структурах мозга. Таким образом, помимо пути от гена к поведению, когда наследственная предрасположенность влияет на характер агонистического поведения, существует и путь от поведения к гену, когда изменение или формирование определенного типа поведения под влиянием средового воздействия приводит к изменению экспрессии определенных генов, в первую очередь, тех, продукты которых вовлечены в регуляцию работы медиаторных систем. Работа с животными выполнялась в Центре генетических ресурсов лабораторных животных ИЦиГ СО РАН (№ RFMEFI61914X0005).
Генетика агрессивности человека
В обзоре Анхольт и Макей (2012) приводятся довольно полные сведения о генетической детерминации агрессивности человека. При этом целесообразно отметить, что речь идет о тех же генах: транспортере серотонина и его рецепторах, ферментах биосинтеза и катаболизма катехоламинов и их рецепторах, которые были выявлены в экспериментальных исследованиях на животных.
Уровень наследуемости агрессивности у людей весьма высок. В Голландии при исследовании близнецов в возрасте 3–10 лет коэффициент наследуемости элементов агрессивного поведения составил с высокой степенью достоверности от 0,51 до 0,72 (Hudziak et al., 2003), а при исследовании взрослых близнецовых пар – от 0,37 до 0,57 (Yeh et al., 2010). Отметим, что в поле зрения исследований агрессивного поведения человека попадают в основном случаи явно выраженных агрессивных наклонностей и проявлений, относящихся к патологической агрессии, так как она зачастую сочетается с наличием тех или иных нейропсихических нарушений. Те виды агрессии, которые носят характер, скорее, не физических, а психологических воздействий, остаются практически вне сферы генетических исследований. Им трудно найти аналоги у животных. Возможно, та разновидность агрессивного поведения животных, которая обозначается как демонстрации, может служить некоторым подобием «психологической» агрессии человека, хотя авторам обзора не приходилось встречать в литературе исследований такой аналогии.
Итак, патологическая агрессия человека обычно является следствием либо травматического поражения мозга, либо нейропсихических нарушений, алкоголизма, наркомании, или результатом нейродегенеративных процессов. Физическая агрессия и импульсивные реакции наблюдаются у 20–65 % пациентов с болезнью Альцгеймера. Генетические факторы сочетанного проявления агрессии с нейродегенерацией остаются неизвестными. Мы полагаем, что так как при нейродегенеративных процессах страдают, прежде всего, высшие центры регуляции поведения, в результате отменяются «социальные табу», ограничивающие проявления инстинктивных императивов, которые при наличии генетической предрасположенности находят выражение в импульсивном агрессивном поведении. Вследствие этого внимание исследователей сосредоточено на генах, которые, с одной стороны, ассоциированы с агрессивным поведением, с другой – подозреваются в участии в нейродегенеративных явлениях. Так, эпизоды агрессии фиксируются чаще у пациентов с болезнью Альцгеймера, гомозиготных по аллелю e4 гена аполипопротеина E (исследованы группы численностью 100–400 пациентов) (Craig et al., 2004а; van der Flier et al., 2007).
Однако наличие такой ассоциации не подтвердилось при изучении более многочисленной когорты пациентов с болезнью Альцгеймера (1120 больных) (Hollingworth et al., 2006). В ряде исследований изучалось участие в ассоциации агрессии с болезнью Альцгеймера полиморфизмов генов триптофан гидроксилазы (Craig et al., 2004b), рецепторов серотонина 5HT-2A и 5HT-2C (Assal et al., 2004; Pritchard et al., 2008) и транспортера серотонина (Sweet et al., 2001; Assal et al., 2004; Pritchard et al., 2007).
Однако результаты получились противоречивыми. Ассоциация агрессии с шизофренией в некоторой степени связана с полиморфизмом гена катехол-О-метил трансферазы – comtVal158met (Strous et al., 1997; Kotler et al., 1999; Tosato et al., 2011). У шизофреников и алкоголиков, гомозиготных по аллелю met, активность этого фермента снижена, у них же повышена агрессивность и отмечается склонность к суициду, который рассматривается как агрессия, направленная на самого себя (Nolan et al., 2000; Nedic et al., 2011). У людей различия в агрессивном поведении часто связывают с полиморфизмом по гену дофаминового транспортера Dat1 (Young et al., 2002b; Chen et al., 2005). Однако эти результаты следует принимать с осторожностью ввиду малых выборок, наличия в проявлении ассоциаций половых различий, а также разницы в историях болезни исследованных пациентов и в связи с различиями их условий жизни и гетерогенностью в манифестации психиатрической симптоматики. Кроме того, интерпретация влияния полиморфизма генов-кандидатов на нейропатологическую агрессию осложняется трудностями установления связей между заболеванием и непосредственными причинами формирования агрессивного фенотипа. Хорошо известный из литературы случай действительной идентификации генов агрессии человека получен в 1993 г. при изучении большой голландской семьи (Brunner et al., 1993). У нескольких мужчин из этой семьи были отмечены задержка умственного развития и склонность к насилию, которая выражалась в импульсивных актах агрессии, поджогах, попытках изнасилования, эксгибиционизме. У этих мужчин была снижена активность моноаминоксидазы А (МАОА), фермента катаболизма биогенных аминов в центральной нервной системе, по причине наличия точковой мутации в восьмом экзоне гена Мao-а, которая приводила к образованию кодона терминации. Последующие работы подтвердили наличие ассоциации между полиморфизмом гена Мao-a и агрессивностью у человека (Manuck et al., 2000; Huang et al., 2004), что аналогично результатам, полученным в эксперименте на животных (Cases et al., 1995; Newman et al., 2005).
 На людях также показано, что влияние отдельных генов на агрессивное поведение сильно зависит от генетического фона. Так, эффект аллелей генов Мao-а и серотонинового транспортера на агрессивное поведение во многом определяется предшествующим опытом, особенно условиями раннего развития и воспитания (Caspi et al., 2002, 2003; Huang et al., 2004; Kim-Cohen et al., 2006; Lazary et al., 2008; Kinnall et al., 2009; Weder et al., 2009; Edwards et al., 2010). Причем хорошие условия воспитания, наличие хорошей родительской заботы блокируют проявление генов агрессивности, в то время как плохие условия и отсутствие родительской заботы способствуют проявлениям агрессивности.
На людях также показано, что влияние отдельных генов на агрессивное поведение сильно зависит от генетического фона. Так, эффект аллелей генов Мao-а и серотонинового транспортера на агрессивное поведение во многом определяется предшествующим опытом, особенно условиями раннего развития и воспитания (Caspi et al., 2002, 2003; Huang et al., 2004; Kim-Cohen et al., 2006; Lazary et al., 2008; Kinnall et al., 2009; Weder et al., 2009; Edwards et al., 2010). Причем хорошие условия воспитания, наличие хорошей родительской заботы блокируют проявление генов агрессивности, в то время как плохие условия и отсутствие родительской заботы способствуют проявлениям агрессивности.
Эта же закономерность получена при исследовании на животных (крысы, обезьяны) и, по-видимому, является для животных универсальной. Ассоциация полиморфизма промоторного района гена серотонинового транспортера с агрессивным поведением описана в нескольких работах (May et al., 2010; Gonda et al., 2011; Takahashi et al., 2011). Однако данные о взаимосвязи этого полиморфизма с суицидальными наклонностями, которые расцениваются как проявление агрессии, направленной на себя, были весьма противоречивыми (Bondy et al., 2006; Roy et al., 2009, Tsai et al., 2011). Была обнаружена ассоциация полиморфизма («короткий аллель–длинный аллель») промоторного района гена серотонинового транспортера (SLC6A4) с агрессивным поведением, а также с различными нейропсихическими нарушениями (Murphy et al., 2008).
В дополнение к вопросу о роли серотонина анализировалось значение таких нейромедиаторов, как дофамин, норадреналин и ГАМК. В результате обнаружены ассоциации между девиантным поведением подростков и полиморфными аллелями дофаминового рецептора, рецептора ГАМК и серотонинового транспортера (Miczek et al., 2002; Marino et al., 2005; Hohmann et al., 2009). Следует отметить, однако, что описанные ассоциации полиморфных аллелей с агрессивным поведением не всегда воспроизводятся при исследовании разных популяций. Выявление влияния полиморфных аллелей генов Мao-а и серотонинового транспортера – это только верхушка айсберга, малая часть той нейрогенетической системы, которая определяет наличие или отсутствие агрессивного поведения или его выраженность. Хорошо известно, что эндокринная система влияет на проявление и выраженность агрессии. Виновником чаще всего объявляется мужской гормон – тестостерон. Данные правительства США говорят о том, что мужчины совершают убийства в 10 раз и уголовные преступления в 5 раз чаще, чем женщины (http://www.ojp.usdoj.gov/bjs/ ).
Аналогичные результаты дает статистика преступлений в других странах (Moffitt et al., 2001). Множество данных свидетельствуют о наличии хотя и небольшой (r=0,14), но статистически достоверной корреляции между уровнем тестостерона в плазме крови и агрессивным поведением как у животных (Archer, 1991), так и у человека (Book et al., 2001). М. Вильсон и М. Дэлэй (Wilson, Daly, 1985) характеризуют агрессивность молодых людей и подростков как «синдром молодого мужчины» (Young male syndrome). Это предполагает наличие связанных с полом генов, влияющих как на синтез тестостерона, так и на уровень агрессивности, однако поиск таких генов на Y-хромосоме с помощью весьма изощренных экспериментов оказался практически безуспешным (Gatewood et al., 2006). Возможно, дело в том, что тестостерон создает лишь определенный гормональный фон, на котором лучше могут больше манифестировать другие гипотетические гены агрессивности. Выявление генетической базы агрессивного поведения человека является сложной задачей в связи с генетической гетерогенностью популяций людей и трудно контролируемыми средовыми влияниями.
Для полного представления о нейрогенетической базе такого универсального свойства живых организмов, как агрессия, необходимы широкие сравнительно-генетические исследования и анализ, основанный на использовании эволюционного подхода для объяснения происхождения и биологического смысла различных проявлений агрессивного поведения. Кроме того, исследование генетики агрессивности человека сосредоточено на анализе крайних форм агрессивности, в то время как генетическая основа всего спектра агрессивно-неагрессивного поведения, которое градуально изменяется от нерешительности к уверенности, от подавленного гнева к явному его проявлению, от угрозы насилия к прямому насилию и т. д., часто выпадает из поля зрения. Фенотипическая вариабельность агрессивных проявлений и множественность модифицирующих факторов, а также субъективность оценок поведения сильно усложняют возможность количественной оценки уровня агрессивности при проведении полногеномного анализа ассоциаций.
Эти практически неразрешимые вопросы фактически не оставляют другого выбора, как искать ключ к решению проблемы генетики агрессивного поведения в очень узком круге имеющихся возможностей («поиск потерянного ключа под фонарем»), который фактически ограничивается изучением биогенных аминов мозга и нейротрофических факторов. Поэтому прогресс в изучении сложной архитектуры генетической детерминации агрессивного поведения и агрессивности видится в области эксперимента на модельных организмах животных, у которых можно провести аккуратную оценку фенотипических проявлений агрессивности в разных, но контролируемых условиях и на выровненном генетическом фоне. Сравнительно-генетические исследования помогут выявлению генов-ортологов и установлению эволюционно консервативных принципов организации генетической системы детерминации агрессивности.
Исследование механизмов агрессивного поведения с помощью современных протеомно-транкриптомных технологий
Mетаболомно-протеомные исследования
Исследования, выявившие изменения активности моноаминергических систем мозга на уровне метаболизма, активности ферментов и рецепции (обзор Kudryavtseva, 2006) как в процессе формирования патологии агрессивного поведения у мышей, так и при селекции на выраженность агрессивного поведения у крыс-пасюков (Popova et al., 2005), позволяют надеяться, что их расширение с помощью новых комплексных метаболомно-протеомных методов, использующих совокупность всех метаболитов, являющихся конечными продуктами синтеза в клетке, ткани, органе и организме, а также белков, вовлеченных в обменные процессы, будет продуктивным в плане понимания сложных механизмов регуляции агрессии. Поскольку формирование различных патологий поведения сопровождается перестройкой работы мозга, то исследование устойчивости и динамики таких состояний может раскрыть многие новые механизмы, позволяя изучать, например, развитие болезни от ее начала до глубокой патологии.
Tранскриптомный анализ
Бурное развитие молекулярных технологий в последние годы, начиная с момента появления метода полимеразной цепной реакции (ПЦР) (Chattopadhyay et al., 1993; Wong et al., 1994; Auboeuf, Vidal, 1997), открыло перспективы в исследовании вовлеченности генов в механизмы регуляции различных сложных форм поведения. Метод позволил давать количественную оценку содержания мРНК в минимальных объемах ткани. Очевидна ценность применения этих методов для исследования роли отдельных генов в механизмах регуляции нейрохимических процессов в разных структурах мозга, величина которых очень небольшая, а функциональные роли их существенно различаются. Это значит, что имеется возможность исследовать взаимосвязь между определенными формами поведения, нейрохимическая регуляция которых определяется в разных структурах мозга, причем как при функциональных физиологических, так и при генетических изменениях, например, связанных с селекцией.
В настоящее время распространено мнение, что физиологические процессы приводят к индукции целого ряда генов, ответственных за запуск долговременных перестроек фенотипа, метаболизма и связей нервных клеток. Показано, что экспрессия некоторых генов может быстро увеличиваться или снижаться, в то время как экспрессия других генов может изменяться градуально (Nichols et al., 2003). Так, были найдены динамические изменения экспрессии гена каппа-опиоидных рецепторов (Spangler et al., 1996; Goloshchapov et al., 2005), проэнкефалина (Crespo et al., 2001), мю-опиоидных рецепторов (Azaryan et al., 1998; Nikulina et al., 1999) в различных экспериментальных ситуациях. Нами было продемонстрировано изменение экспрессии, по крайней мере, 12 генов (из 15 исследованных) в структурах мозга самцов мышей под влиянием повторного опыта агонистических взаимодействий (Филипенко и др., 2001; Philipenko et al., 2001, 2002а, b; Филипенко и др., 2002; Кудрявцева и др., 2004; Goloshchapov et al., 2005; Bondar et al., 2009; Kudryavtseva et al., 2010; Boyarskikh et al., 2013; Smagin et al., 2013).
При выборе генов и структур мозга, в которых предполагалось изменение их экспрессии, мы руководствовались, прежде всего, ролью этих структур в механизмах агонистического поведения, которая была подтверждена изменением каких-либо параметров активности медиаторных систем, а также, в первую очередь, анатомическим строением и связями этих структур (Кудрявцева и др., 2004). Это были функциональные гены, которые предположительно должны вовлекаться в процесс работы медиаторных систем в том случае, когда другие возможности регуляции по механизмам обратной связи на уровне метаболизма и рецепции исчерпаны. Как правило, это должно происходить при формировании патологических состояний, основным критерием развития которых является длительность сохранения изменений, возникающих под влиянием психопатогенного воздействия. Таким образом, не всегда, а только при определенных состояниях организма функциональная активность генов может изменяться под влиянием средовых воздействий.
Подход к изучению генетической базы агрессивности и агрессивного поведения, который начал использоваться сравнительно недавно, связан с исследованием полного транскриптома. В 2014 г. появилась статья К. Малки и соавт. (Malki et al., 2014), которая, по утверждению авторов, является первой попыткой анализа транскриптома префронтальной коры у мышей трех линий, селекционированных по агрессивному поведению, по сравнению с тремя контрольными неагрессивными линиями. Экспрессия генов клеток префронтальной коры исследована с помощью микроматриц, позволяющих оценить более 45 тыс. транскриптов генома мыши. В результате выявлены две основные группы генов, ассоциированных с агрессивным поведением мышей, которые включены в две генные сети.
Таким образом, было выделено два «генных модуля», статистически достоверно связанных с проявлениями агрессивности, которые детерминировали функцию двух основных метаболических путей. Ключевым центром одного из них является ген NF-kB (nuclear factor kappa-light-chain-enhancer of activated cells) – это ген транскрипционного фактора, функция которого имеет особое значение для регуляции работы нейронов, клеток глии и иммунной системы. Ключевыми центрами второго генного пула являются гены MAPKs (mitogen-activated protein kinases) – это mitogen-activated protein kinases) – это -activated protein kinases) – это activated protein kinases) – это protein kinases) – это гены ферментов, которые обеспечивают такие фундаментальные процессы, как пролиферация и дифференцировка клеток, апоптоз, реакция клеток на стресс и другую стимуляцию. Ясно, что функция выявленных генных сетей многообразна, практически универсальна, и нет ничего удивительного, что эти гены и метаболические пути вовлечены в регуляцию агрессивного поведения. В составе выделенных генных сетей авторы идентифицировали 14 генов-кандидатов, которые с большей или меньшей степенью статистической вероятности ассоциированы с агрессивным поведением мышей. Интересно, что среди этих генов нет ни одного из того списка генов, которые были обнаружены ранее в мутантных линиях.
Можно предположить, что различия по агрессивному поведению линий мышей, использованных для анализа транскриптома, связаны с генами, каждый из которых оказывает минорное влияние на агрессивность, и их эффект обнаруживается лишь при совокупном их действии. Влияние одиночных генов в случаях мутации или нокаута выявляется в соответствующих моделях, когда эти гены могут оказывать сильное действие на поведение. Но даже в этих случаях, как было показано выше, эффекты так называемых майор-генов могут сильно модифицироваться средой и/или генетическим окружением. Из всего списка генов-кандидатов в качестве «многообещающего» авторы выделяют ген киназы-2 бета-адренорецепторов (Adrbk2 или Grk3). Интерес авторов к этому гену связан с тем, что мутацию гена Grk3 человека находят у 10 % больных биполярным или маниакальнодепрессивным психозом (МДП) (Parruti et al., 1993). У погибших больных с МДП уровень мРНК гена Grk3 во фронтальной коре снижен в 2,5 раза (Rao et al., 2009).
Перспективы использования транскриптомных исследований
Молекулярные методики полного секвенирования транскриптома – всего набора РНК в клетке с использованием высокопроизводительных инструментальных платформ массового параллельного секвенирования, таких как Illumina и SOLiD, основанные на секвенировании фрагментов кДНК, позволят определить гены, специфично экспрессирующиеся в исследуемой ткани у животных с опытом агрессии. К таким тканям организма относятся, прежде всего, структуры мозга и надпочечников, вырабатывающих гормоны стресса. Повторные эксперименты по сравнению агрессивных и неагрессивных животных, а также животных с различным опытом агрессии позволяют выделить группы дифференциально экспрессирующихся генов. Полнотранскриптомный анализ позволит также выявлять совокупность всех транскриптов, синтезируемых группой клеток, включая мРНК и некодирующие РНК, и охватывает полный спектр изменений экспрессии генов всего организма.
При этом транскриптом может сильно меняться в зависимости от условий окружающей среды, отражая профиль экспрессии генов в данный момент времени. Транскриптомы, полученные из разных отделов мозга с различной специализацией (вентральная тегментальная область, содержащая скопления дофаминергических и опиоидергических нейронов и ответственная за механизмы положительного подкрепления; ядра шва среднего мозга, содержащие скопления серотонергических нейронов, ответственных за ингибирующий контроль агрессивного поведения; миндалина, участвующая в контроле эмоций; гипоталамус, участвующий в регуляции стресс-реактивности) у агрессивных животных, могут ответить на вопрос о механизмах избирательного участия этих отделов и оценить характер специфичности их работы в сравнении с неагрессивными животными, независимо от того, получены ли они в результате селекции или под влиянием средовых воздействий.
Таким образом, применение метода секвенирования всего транскриптома в отделах мозга позволит определить значение наследственно обусловленных факторов и средовых влияний в детерминации проявления агрессивных реакций и отделить одни изменения от других. Анализ метаболических и транскриптомных карт, построенных на основании полученных результатов, позволит выявить узловые точки и основные молекулярные мишени, воздействуя на которые возможно осуществить коррекцию аномального поведения. В связи с развитием новых геномных технологий появляется возможность ответить на главные вопросы, которые ставит перед нами генетика агрессивного поведения:
- какие транскрипционные факторы ответственны за функционирование генов, вовлеченных в регуляцию агрессивности, и как осуществляется взаимосвязь регуляторных процессов на всех уровнях функционирования системы агрессивных реакций?
- какова молекулярная природа нейрохимической избирательности воздействия на функционирование того или иного гена в разных структурах мозга у животных с контрастными типами социального поведения?
- какие гены (major genes) играют ключевую роль в формировании разных нейрофизиологических статусов с противоположными поведенческими, нейрохимическими, физиологическими и психоэмоциональными особенностями, наблюдаемыми у животных одного генотипа?
На эти вопросы поможет ответить нейроинформатика поведения, которая будет анализировать результаты транскриптомных исследований на основе данных профилирования мРНК, выделения дифференциально экспрессирующихся генов, реконструкции генных сетей по экспрессионным данным.
Нейроинформатика поведения
В сферу нейроинформатики входит сбор результатов, полученных в ходе нейробиологических исследований, перевод этих результатов в формат баз данных для их последующего анализа с помощью вычислительных моделей и специализированных компьютерных аналитических программных инструментов. В задачи биоинформатики входит изучение особенностей экспрессии генов в структурах мозга, передачи сигнала между органами и между отдельными клетками (работа нейронов, секреция). Отметим, что задачи компьютерной геномики, смежные с анализом экспрессии генов, и описание регуляторных характеристик геномной ДНК, критичных для определения уровня транскрипции, решены лишь частично. Описание механизмов передачи информационных сигналов, связанных с поведением, должно опираться на реконструкцию сетей взаимодействий между структурами мозга и группами клеток. Необходима реконструкция генных сетей – комплексов взаимодействующих макромолекул в клетке, определяющих пути передачи сигналов и взаимодействия между клетками и органами. В рамках молекулярно-генетических исследований с помощью полногеномного секвенирования (RNA-seq) могут быть выявлены дифференциально экспрессирующиеся гены в отделах мозга у агрессивных и неагрессивных животных, полученных путем селекции или в результате развития патологии агрессивного поведения. Оценка уровней экспрессии этих генов, определенная в масштабе генома, должна подтверждаться протеомными данными и ПЦР в реальном времени.
Полногеномный анализ экспрессии генов на уровне транскрипции в исследуемых областях мозга животных поможет найти новые маркеры, связанные с агрессивным поведением, поскольку технологии микрочипов и в последующем более прогрессивные транскриптомные технологии изменили понимание о взаимосвязи фенотипических проявлений с развитием и поведением животных. Сравнение с доступными данными по гомологичным генам у человека, в том числе геном наследственных заболеваний, и выявление сходства между предполагаемой патологией поведения, развивающейся у животных под влиянием повторного опыта агрессии, и определенным психоневрологическим расстройством у людей могут быть приняты за критерий адекватного моделирования заболевания. В результате объединения классических и современных методов исследования станет возможным поиск генов-маркеров агрессивности и кандидатов для терапии, выделение белковых продуктов, связанных с агрессивным поведением.
Эпигенетическая регуляция агрессивного поведения
Кратко остановимся на возможности эпигенетической регуляции агрессивного поведения (Archer et al., 2012; Provençal et al., 2014). Данные об изменении процессов метилирования ДНК и деацетилирования гистонов, ремоделировании хроматина в клетках мозга появились сравнительно недавно, но их важность трудно переоценить (LaPlant et al., 2010). В опытах на мышах было показано, что снижение материнской заботы о потомстве в период вскармливания приводит к серьезным изменениям поведения взрослых мышей, при этом сильно повышается уровень межсамцовой агрессии (Weaver et al., 2004). Напротив, материнская агрессия самок, испытавших дефицит материнской заботы при вскармливании, снижается, но у них же нарушается материнское поведение относительно собственного потомства (Veenema et al., 2007). Аналогичное влияние недостаточной родительской заботы на повышение агрессивности у мальчиков отмечено в данном обзоре выше. Влияние материнской заботы на агрессивное поведение взрослых потомков связывают с изменением уровня метилирования гена глюкокортикоидных рецепторов в гиппокампе (McGowan et al., 2009).
Гиперметилирование промоторного района приводит к снижению экспрессии гена глюкокортикоидного рецептора, что, в свою очередь, нарушает адекватное функционирование отрицательной обратной связи в системе, регулирующей уровень стрессовых реакций. Повышение стресс–реактивности сопровождается усилением импульсивного, в том числе и агрессивного, поведения. Такого же рода изменения метилирования отмечены для промоторного района гена Мао-а (Pinsonneault et al., 2006), что в случае снижения транскрипционной активности этого гена может приводить, как было отмечено выше, к повышению проявлений агрессивного поведения. Таким образом, обнаружение влияния эпигенетических механизмов на регуляцию агрессивного поведения открывает новое направление исследований, которое должно значительно обогатить наши представления о всех возможных путях и способах изменения проявлений агрессивности, что, несомненно, востребовано нашей жизнью.
Заключение
Таким образом, в обзоре представлены современные данные по изучению генетико-физиологических механизмов агрессивности как общебиологического феномена. Рассмотрены подходы к экспериментальному исследованию механизмов внутривидового агрессивного поведения у лабораторных животных, приведены данные по влиянию генетических факторов на агрессивное поведение человека. На примере исследования механизмов регуляции одной из наиболее сложных и универсальных форм социального поведения – внутривидовой агрессии – рассмотрены различные подходы к ее изучению, использовавшиеся ранее, и перспективы, открывающиеся с появлением новых молекулярных подходов, таких как секвенирование транскриптома. Развитие протеомных и метаболомных технологий позволит исследовать механизмы регуляции сложных форм поведения уже на более высоком уровне организации молекулярно-генетической информации. В целом можно заключить, что основой такого сложного биологического феномена, как агрессивное поведение, является многофакторная система, включающая, с одной стороны, несколько генетических ансамблей, связанных между собой многочисленными регуляторными взаимодействиями, на основе которых выстроена еще более сложная нейроэндокринная физиологическая регуляторная система. С другой стороны, большую роль играет внешний средовой контекст, который включает как физические, так и социально-биологические характеристики среды существования особей и популяций. Причем эндогенные и средовые детерминанты взаимосвязаны и взаимообусловлены. Исследование такого рода систем требует координированных усилий биологов, физиологов, генетиков, специалистов по системному анализу и биоинформатике.
Вавиловский журнал генетики и селекции, 2014, том 18, № 4/3
Примечание
1Влияние этого фактора самого по себе на приматов часто переоценивается и в целом довольно сомнительно.
Рекомендуем прочесть
 Синдром социальности: сравнительное исследование поведения песчанок
Синдром социальности: сравнительное исследование поведения песчанок Два механизма «копирования поведения»
Два механизма «копирования поведения» Социальный контроль поведения млекопитающих: ревизия концепции доминирования
Социальный контроль поведения млекопитающих: ревизия концепции доминирования Механизм доминирования самок у бонобо
Механизм доминирования самок у бонобо Механизм становления отцовской заботы о потомстве
Механизм становления отцовской заботы о потомстве Часть и целое
Часть и целое Об агрессии-2
Об агрессии-2 Об агрессии
Об агрессии
