

Н. А. Колчанов, В. В. Суслов, М. П. Пономаренко
В организации регуляторных районов генов, активных сайтов белков и в генных сетях присутствуют эволюционно-консервативные модули, задающие пространства возможностей, ограничивающие изменчивость и канализирующие эволюцию. Это позволяет по-новому взглянуть на закон гомологических рядов Н. И. Вавилова и другие случаи параллельной эволюции. Модули самоподдерживаются за счет дупликаций и конвергентной молекулярной эволюции. В то же время, выводя из-под отбора часть мутаций, модули способствуют их накоплению в квазинейтральном режиме, подготавливая дальнейшую эволюцию и обеспечивая ее неповторимость.
Введение
Содержание
Сравнительно-геномные исследования вновь подняли вопрос о темпах молекулярной и морфологической эволюции. Во-первых, полногеномные исследования выявили низкий уровень молекулярной дивергенции в целом по геномам в сочетании с высоким уровнем дивергенции по генам отдельных систем. Так, несмотря на большие морфологические различия между человеком и шимпанзе, полногеномные сравнения дают оценки различий в пределах 1–2 % — не выше, чем между шимпанзе и другими человекообразными обезьянами (Glazko et al., 2005; CSAC, 2005).
В то же время сравнение генов-ортологов позволяет выявить консервативные и быстро адаптивно эволюционирующие функциональные группы генов. Так, у млекопитающих быстро адаптивно эволюционируют гены иммунитета, гены, связанные с репродукцией, гены ольфакторных рецепторов и гены модификации хроматина (CSAC, 2005; Arbiza et al., 2006). Во-вторых, полногеномные исследования заставили пересмотреть монофилию таксонов, выстроенную на эмбриональном и морфобиохимическом сходстве. Так, у эукариот окончательно оставлена монофилия Coelomata, оспорено единство ракообразных и их родство с насекомыми (Шаталкин, 2003; Cook et al., 2005; Regier et al., 2005), зато «восстановлен в правах» фагоцителлоподобный общий предок Metazoa (первичность билатеральной организации по сравнению с радиальной и ранее обособление Acoela) (Garcia-Fernàndez, 2005; Ogishima, Tanaka, 2007; Ryan et al., 2007).
У прокариот поставлена под сомнение монофилия такой консервативной группы, как цианобактерии (Захаров-Гезехус, 2008). С учетом этих данных картина эволюции больше соответствует не классическому дереву с длинным стволом и ветвями разной длины, а «кусту», ветвящемуся у корня. Большинство веток «куста» эволюционируют параллельно, что и объясняет сравнительно-анатомическое и эмбриологическое сходство, а также низкий уровень молекулярной дивергенции в целом по геномам.
Классическое определение параллельной эволюции подразумевает сходный ответ общей генетической основы на сходные векторы отбора (Грант, 1991). Однако трудно предположить общий вектор отбора на протяжении миллионов лет для такой морфологически разнообразной и эволюционно пластичной группы, как ракообразные. Морфологически сходные, но генетически отдаленные линии цианобактерий населяют такие разные биотопы, как горячие источники, моря и пресные водоемы.
В чем причина широкого распространения параллелизмов? В статье рассмотрена молекулярно-генетическая организация генных сетей с точки зрения существования эволюционно-консервативных модулей. Самоподдерживаясь за счет дупликаций, молекулярной конвергенции, координационных замен, они формируют пространство возможностей, канализирующее эволюцию.
Закон гомологических рядов Н. И. Вавилова
Широкое распространение параллелизмов заставляет вспомнить закон гомологических рядов Н. И. Вавилова (1920, 1935):
1) виды и роды характеризуются сходными рядами наследственной изменчивости с такой правильностью, что, зная ряд форм в пределах одного вида, можно предсказать параллельные формы других видов и родов;
2) семейства в общем характеризуются определенным циклом изменчивости, проходящим через все роды и виды.
Для формальной записи параллельных (гомологических) рядов изменчивости был предложен радикал вида — запись признаков в порядке констанности их проявления. Радикалы, по Н.И.Вавилову, возникают на базе общего пула генов-гомологов, дополненного общим сходством физиологического пути от гена до признака (здесь Н.И.Вавилов ссылается на Ю. А. Филипченко, 1925 и А.А.Заварзина, 1923, 2000).
Морфотип любой популяции можно описать радикалом. Его левая часть — константные признаки — практически одинакова у всех особей, характеризуя таксон высокого порядка — род, семейство, отряд. Правая часть — неустойчивые признаки — распадется по степени варьирования на группы, соответствующие низшим таксонам (от подвида до жорданона — монотипической популяции)1 (Вавилов, 1931, 1935, 1967). Уникальное сочетание константных и варьирующих признаков «физиономически» характеризует вид, причем, в отличие от современных концепций вида, константные и варьирующие признаки одинаково важны, а репродуктивная изоляция, если она есть, является лишь одним из признаков в радикале (Вавилов, 1931; Васильева, 2002).
Формирование радикала вида в эволюции Н. И. Вавилов (1931) объяснял через взаимодействие генетической составляющей дарвиновской неопределенной изменчивости2 предковой популяции с разнообразием контрастных физико-географических факторов внешней среды в пределах небольшого ареала.
Таким условиям удовлетворяли центры происхождения культурных растений (Вавилов, 1965), приуроченные к гористым районам с пересеченной местностью и разнообразными, часто контрастными условиями. Здесь вся изменчивость небольших популяций-полуизолятов имела наилучший шанс проявиться и быть закрепленной отбором3 — из первоначального полиморфизма выделялась левая, константная часть радикала. Тогда его правая часть — либо незавершенный эволюционный процесс, либо признаки, чей полиморфизм выгоден виду (системе популяций — линнеону) в целом (Вавилов, 1931, 1965). Позже М. М. Камшилов (1935) на дрозофиле смоделировал формирование константной части радикала в контрастных (Камшилов, 1935) и меняющихся (Камшилов, 1939, 1940 а, б) условиях среды в популяции с исходно изменчивым значением признака.
Одновременно с Н.И.Вавиловым Д. Н. Соболев (1924) построил циклы изменчивости (филоциклы) упрощения–усложнения сутуры отрядов аммонитов. Гомологические ряды в палеонтологии оказались удобны для прогноза. Так, пустые клетки в таблице археоциат А.Ю.Розанова (1973) продолжают заполняться4. Идеи Н. И. Вавилова нашли горячий отклик у Л. С. Берга (1977). Изучая сходные ряды изменчивости у строго изолированных групп (отдаленные таксоны, например рыбы и млекопитающие, либо географические изоляты — популяции озерных и речных рыб), он предположил, что сохранение генетической общности у них маловероятно, поэтому отверг творческую роль отбора, предложив собственные динамические законы эволюции, зашифрованные в конформации белков. Ограниченный набор конформаций задает пространство возможностей для эволюции, обеспечивая повторяемость гомологических рядов.
В 70-е годы XX в. академик Г.А.Заварзин (1973) ввел понятие «пространство логических возможностей» для взаимоисключающих признаков в систематике бактерий. Мы воспользуемся для ограниченного перебора комбинаций признаков термином «пространство возможностей».
Генные сети
Закон гомологических рядов вызвал большой энтузиазм, но концепция вида, объясняющая их эволюционное становление, современниками Н. И. Вавилова не была воспринята. После классической работы Дж. Холдейна (Haldane, 1927) по окраске млекопитающих гомологические ряды стали связывать только с гомологичными генами, что хорошо совпало с концепцией «один ген — один фермент» (Beadle, Tatum, 1941). Но строить, опираясь на столь простой подход, гомологические ряды не удавалось и они практически выпали из синтетической теории эволюции, хотя широко (и часто неосознанно) применялись в медицине (модели патологий на животных) и селекции.
В отличие от биохимических моногенных признаков, с которыми работал Дж. Холдейн, гомологические ряды Н. И. Вавилова построены на материале более сложных морфофизиологических признаков. Формирование таких признаков, как показал М. М. Камшилов (1935, 1939, 1940а, 1940б, 1974), возможно лишь на базе сложных коэкспрессирующихся комплексов множества генов, связанных регуляторными взаимодействиями.
Для таких комплексов С. Кауфман ввел термин «генная сеть» (ГС) (Kauffman, 1977). Любая ГС имеет:
1) группу генов («ядро»), обеспечивающую выполнение ее функций;
2) центральные регуляторы — транскрипционные факторы (ТФ), координирующие гены «ядра», взаимодействуя с сайтами связывания (ССТФ) в их регуляторных областях;
3) рецепторы, запускающие работу ГС в ответ на внешние сигналы;
4) пути передачи сигналов с рецепторов ГС на ее центральные регуляторы. Часть продуктов генов «ядра» ГС играют роль сигналов, взаимодействуя с рецепторами собственной ГС и/или других генных сетей, формируя регуляторные контуры с положительными и отрицательными обратными связями (Колчанов и др., 2005).
Консерватизм генных сетей
Таким образом, вопрос параллельной изменчивости упирается в консерватизм не отдельных генов, а генных сетей. Его можно оценить по микрочиповым профилям коэкспрессии, часто дополненным попарным анализом гомологии последовательностей коэкспрессирующихся генов (Bergmann et al., 2004) и анализом сетей белок-белковых взаимодействий (Xia et al., 2006). Так, сравнение 3182 экспериментов по изучению экспрессии ортологов Homo sapiens (6591 ген), Drosophila melanogaster (5802 гена), Caenorhabditis elegans (5180 генов) и Saccharomyces cerevisiae (2434) выявило 22 163 консервативных случая коэкспрессии. Они распределились по 11 консервативным паттернам, состав генов в которых коррелировал с хорошо изученными генными сетями (Stuart et al., 2003), как древними (сети «домашнего хозяйства» и базовых метаболических процессов — клеточный цикл, трансляция, транскрипция, протеосомная деградация белков и др.), так и молодыми (например, сети нейрогенеза, сигнальных путей и т. д.).
Паттерны коэкспрессии можно разделить на три типа (Bergmann et al., 2004; Stuart et al., 2003).
Генным сетям базовых процессов «домашнего хозяйства» соответствуют паттерны с высококонсервативным составом и коэкспрессией ортологов, а также высокосвязным графом (тип 1).
Паттерны, где состав ортологов варьировал при переходе от таксона к таксону (ГС регуляции транскрипции, межклеточной коммуникации), связаны с появлением новых функций (тип 2);
паттерны, где консервативность состава ортологов не соответствует консервативности коэкспрессии (ГС нейрогенеза) — с дивергенцией функций в эволюции (тип 3).
Три типа паттернов коэкспрессии заставляют вспомнить вавиловский радикал: тип 1 соответствуют левой части, тип 3 — правой части с варьирующим фенотипическим проявлением. Тип 2 связан с признаками, которые невозможно включить в радикал на данном таксономическом уровне.
Для консервативных модулей коэкспрессии предложен термин «регулог» («regulog») (Babu et al., 2004). У прокариот взаиморасположение генов регулога часто консервативно, что связано с оперонной структурой генома (Захаров-Гезехус, 2008).
У эукариот в ряде случаев отмечена связь регулогов и определенных районов хромосом (Taddei et al., 2001; Pauli et al., 2006). В регулогах можно выделить «ядро» из гомологичных генов (ортологов и паралогов) и «шлейф» негомологичных видоспецифичных генов. «Ядра» могут сильно варьировать как по составу генов, так и по степени коэкспрессии (Bergmann et al., 2004). Так, «ядро» ГС циркадного ритма у Metazoa консервативно (Badiu, 2003): мутации в гене казеинкиназы эпсилон I укорачивают циркадный ритм у дрозофилы, грызунов и человека, снижая уровень фосфорилирования белка PER (Vanselow et al., 2006). Мутации по другим казеинкиназам сохраняют гомологичность, хотя направленность нарушений (удлинение или укорочение циркадного ритма) в разных видах может не совпадать (Xu et al., 2005).
Замечательно, что ассоциация с регулогами позволяет предсказывать функцию генов (Stuart et al., 2003, Huynen et al., 2004), что взаимодополнительно к закону гомологических рядов. Высокосвязный граф регулогов типа 1 объясним действием стабилизирующего отбора (Шмальгаузен, 1968) — чем больше процессов, в которые вовлечен ген, тем менее вероятна его эволюция из-за плейотропного эффекта мутаций. О двух других типах этого сказать нельзя. Каков же механизм их эволюционной устойчивости?
Пространство возможностей для эволюции генных сетей
Простейший механизм сохранения регулогов следует из вавиловской концепции вида. Восстановление ГС, чьи элементы повреждены разными мутациями, возможно путем комплементации при контактах между полуизолятами. Так, цветы, характерные для цветной капусты и брокколи, возникают у кочанной капусты при объединении нонсенс-мутаций в экзонах 4 и 5 генов ТФ BoAP1-B и BoCAL, а у арабидопсиса — в их ортологах AP1 и СAL. Эти мутации существуют как полиморфизмы в популяциях у крестоцветных (Lowman, Purugganan, 1999). Таким образом, ареал вида5, формируя сложную популяционную структуру вида6 (Вавилов, 1931), задает внешнее пространство возможностей для эволюции, препятствуя потере потенциального разнообразия в ходе специализации или дивергенции.
Внутреннее пространство возможностей для эволюции формируется модулями генных сетей. Простейший модуль — пара «ген ТФ + ген с ССТФ». Консерватизм таких пар можно оценить попарным внутри- и межвидовым анализом корегулируемых генов (Snel et al., 2004) Критерий корегуляции — наличие общих ССТФ в генах и высокий (r>0,6) коэффициент коэкспрессии. У S. cerevisiae выявлено 975 таких пар. В выборке генов C. elegance для ортологов таких пар также характерен высокий коэффициент коэкспрессии. Наглядно выявляет модули трансгенез. Трансгенез геном Pax-6 (центральный регулятор ГС развития глаза) мыши, асцидии и кальмара индуцировал у дрозофилы развитие фасеточных глаз.
Обратные эксперименты не столь четки (вызвано развитие ряда глазных тканей), что, видимо, связано со сложным строением камерального глаза (Gehring, Ikeo, 1999). Фасеточные глаза появились не позже кембрия, камеральные — в ордовике (Михайлова, Бондаренко 1997), параллельно развившись из светочувствительных клеток плательминтов и книдарий (Gehring, 2005).
Тем не менее достаточно появления нужного центрального регулятора, чтобы воссоздать «ядро» ГС развития глаза! Морфогенез крыльев насекомых и парных конечностей хордовых (с учетом инверсии дорзовентральной оси) также базируется на общих модулях (Шаталкин, 2003).
Таким образом, раз возникнув на заре эволюции, модули используются независимо, многократно и параллельно в разных, порой очень отдаленных таксонах для формирования как гомологичных, так и негомологичных структур. Обрастая «шлейфом» видоспецифичных генов, модули формируют «ядра» регулогов, которые, взаимодействуя друг с другом в глобальной ГС организма, формируют пространство возможностей для дальнейшей эволюции. Так, ложные ноги гусениц по набору корегулируемых генов оказались гомологичны ногам имаго, а ложные ноги личинок пилильщиков — мандибулам (Шаталкин, 2003).
Ясно, что в обоих случаях речь идет не об истинной гомологии, а о развитии параллелизмов на базе различных регулогов. Благодаря экспансии, вероятность потери регуляторного модуля в эволюции мала7. Это объясняет устойчивые миллионы лет гомологические ряды в репродуктивно изолированных таксонах, а также низкий уровень молекулярной дивергенции в целом по геномам. Мутационная реактивация «молчащего» регулога может объяснить филоциклы. Причем принцип Долло (Грант, 1991) соблюдается: генные сети морфологически сходных органов различаются по «шлейфу» видоспецифичных генов.
Пространство возможностей для эволюции в генных сетях
Экспансию модулей обеспечивают дупликации. В паре «ген-ТФ + ген с ССТФ» возможно лишь три варианта: дупликация гена ТФ, дупликация гена с ССТФ и последовательно оба события. У E. сoli по первому пути возникло 272 регуляторных связи, по второму — 128, по третьему — 74. У дрожжей больше роль дупликаций гена ТФ (Teichmann, Babu, 2004). Обеспечивая экспансию модулей, дупликации одновременно снижают интеграцию генных сетей и плейотропность мутаций (Duret, Mouchiroud, 2000), открывая путь движущему отбору.
Другой путь экспансии — конвергентное возникновение ССТФ, чему способствуют их небольшие размеры и ограничения, накладываемые отношением типа «лиганд-рецептор». Так, анализ ССТФ фактора SF-1 в регуляторных районах большой выборки генов выявил сходные особенности встречаемости динуклеотидов и корреляции между частотами динуклеотидов (Levitsky et al., 2007). Параллелизм во взаиморасположении уже не нуклеотидов, а самих ССТФ, образующих консервативные мотивы, характерен для коэкспрессируемых генов. Так, промоторы интерферон-индуцируемых генов обогащены комбинацией сайтов связывания факторов ISGF3, STAT1, IRF1 и NF-kappaB (Ananko et al., 2008).
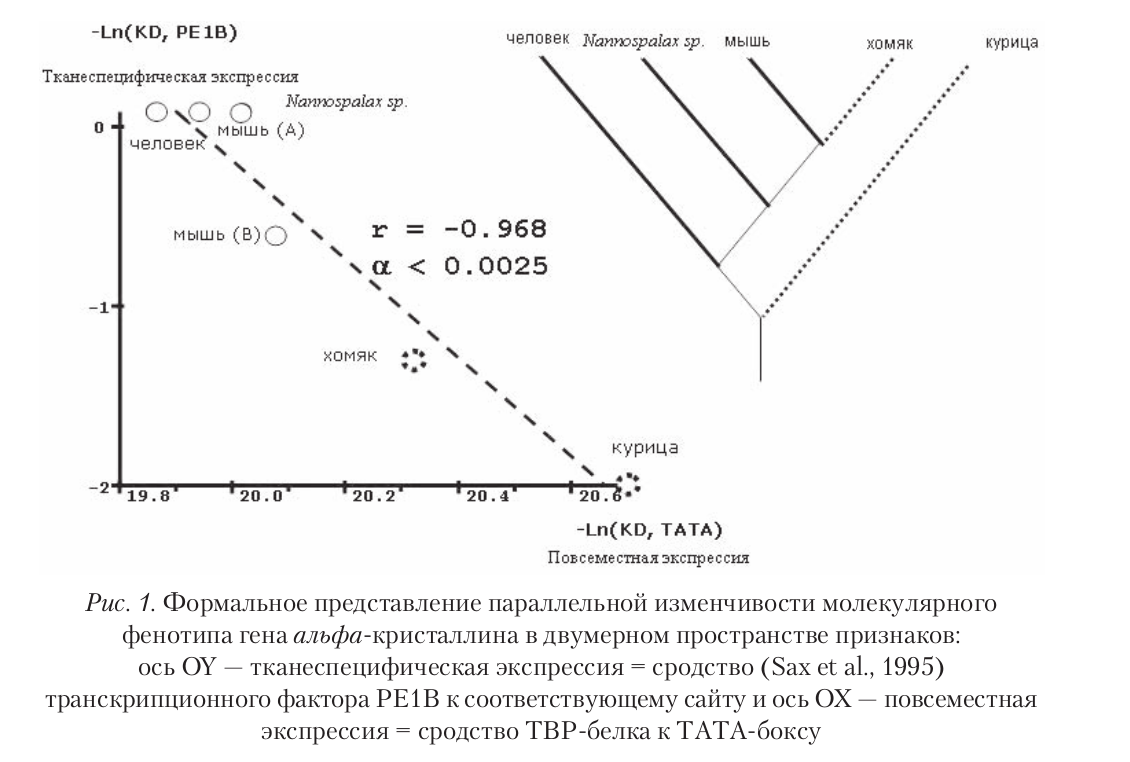
Комбинации нуклеотидов и/или ССТФ формируют свое собственное пространство возможностей для эволюции регуляции отдельных генов, тем более ограниченное, чем меньше регуляторный район. Так, в промоторе гена альфа-кристаллина ССТФ PE1B, отвечающий за тканеспецифичную экспрессию и ТАТА-бокс сближены, образуя композиционный элемент (Sax et al., 1995). Эволюция в сторону эффективности тканеспецифичной экспрессии ведет к усилению PE1B-сродства у человека, мыши и Nannospalax sp. Тренд усиления сродства ТВР-белка к ТАТА-боксу выявлен нами у хомяка и курицы. Другие тренды эволюции здесь просто невозможны (рис. 1).
 Регуляторные модули формируются и на базе пар «миРНК — сайт-мишень в мРНК». Так, миРНК miR319 регулирует экспрессию генов TCP у томата и арабидопсиса. Интенсивное воздействие миРНК на мРНК-мишень дает лист с изрезанным краем (норма томата, мутация у арабидопсиса), снижение интенсивности — цельнокрайний лист (мутация у томата, норма арабидопсиса). Важно, что сходство фенотипов получено разными молекулярными механизмами: у томата снижение воздействия получено за счет мутации, меняющей сродство к миРНК, а у арабидопсиса интенсивность воздействия повышена путем усиления экспрессии miR319 (мутация jaw) (Palatnik et al., 2003).
Регуляторные модули формируются и на базе пар «миРНК — сайт-мишень в мРНК». Так, миРНК miR319 регулирует экспрессию генов TCP у томата и арабидопсиса. Интенсивное воздействие миРНК на мРНК-мишень дает лист с изрезанным краем (норма томата, мутация у арабидопсиса), снижение интенсивности — цельнокрайний лист (мутация у томата, норма арабидопсиса). Важно, что сходство фенотипов получено разными молекулярными механизмами: у томата снижение воздействия получено за счет мутации, меняющей сродство к миРНК, а у арабидопсиса интенсивность воздействия повышена путем усиления экспрессии miR319 (мутация jaw) (Palatnik et al., 2003).
Аналогичные соображения о пространстве возможностей эволюции будут справедливы для любых биополимеров (белки, различные РНК), рабочие структуры которых формируются за счет взаимодействия ограниченного множества мономеров. Так, в молекуле родопсина за длину волны преимущественно поглощаемого света λmax, отвечает участок аминокислот 76–94. Среднее значение изоэлектрической точки и момент гидрофобности аминокислот здесь хорошо коррелируют с λmax. Поэтому замены аминокислот у всех видов, использующих родопсин, повторяются в эволюции (Chang et al., 2002, Briscoe, 2002).
Один из механизмов ускользания вируса гриппа А от иммунного ответа — образование сайтов связывания ионов в антигенной детерминанте. Ион мешает преципитации антител. Небольшие размеры белка ограничивают количество вероятных сайтов связывания ионов. В итоге, у близкородственных штаммов вируса сайты связывания ионов занимают различное положение — перебор пространства возможностей только начат, а у дальнородственных — сходное: перебор пространства возможностей завершен и завершен цикл (филоцикл) изменчивости8 (Иванисенко и др., 2008). Таким образом, для молекулярного фенотипа белка, как ни странно, подходит концепция Л. С. Берга с поправкой на роль отбора.
Консерватизм молекулярного фенотипа активных сайтов белков позволяет делать прогноз. Так, база данных PDBSite (Ivanisenko et al., 2005) содержит библиотеку шаблонов — консервативных трехмерных структур активных сайтов хорошо изученных белков. Программа PDBSiteScan позиционирует эти шаблоны in silico на неизученные белки с целью предсказания функций. Эти исследования показали, что в нативном белке активный сайт может быть окружен набором аминокислот, оптимизированных так, что для появления нового сайта достаточно одной-двух мутаций (Иванисенко и др., 2008). Можно предположить, что кроме действующего пространства возможностей эволюции молекулярного фенотипа существует также потенциальное пространство возможностей, куда белок может быть переброшен определенными мутациями в районе активного сайта.
Матрица взаимодействия между мономерами существенно корректирует пространство возможностей для эволюции, делая его вырожденным по отношению к составу мономеров за счет коадаптивных замен. Эффект коадаптивных замен выявлен как для нуклеиновых кислот (Алёшин и др., 1999), так и для белков (Афонников, Колчанов 2001). Так, в протаминах повышена доля основных аминокислот, что необходимо для компактизации ДНК в сперматозоиде. Мутация, элиминирующая основную аминокислоту, будет повышать вероятность фиксации компенсирующей мутации. В итоге, характерный для протаминов высокий уровень dn/ds (Wyckoff et al., 2000), на деле не связан с изменением их функции9, хотя ведет к превалированию несинонимичных замен и изменению последовательности ДНК (Rooney et al., 2000).
Коадаптивные замены обеспечивают консерватизм структуры рРНК, оставляя при этом как возможность таксон-специфически в квазинейтральном режиме варьировать состав нуклеотидов, так и фиксировать конвергентные замены в неродственных таксонах. (Алёшин и др., 1999).
Разорвать круг филоциклов можно, видимо, двумя способами:
1) переходом в потенциальное пространство возможностей, что стохастически меняет один филоцикл на другой, или
2) прорывом в линейную эволюцию, ведущую к приобретению новых функций.
Последнее возможно лишь после разрушения пространств возможностей. Для белков, по-видимому, это возможно в результате длительного периода эволюции, близкой к нейтральной. Так, эволюция ряда генов, связанных с антропогенезом — FOXP2, BRCA1 (Arbiza et al., 2006), MYH16 (Stedman et al., 2004), — лучше объясняется не движущим отбором, а длительным его ослаблением, с последующей краткой вспышкой адаптивной эволюции.
Заключение: эволюция в пространствах возможностей
Итак, путь от гена до признака можно представить как иерархию различных пространств возможностей:
1) на уровне биополимеров,
2) модулей в локальных генных сетях и
3) регулогов в глобальной ГС организма. Кроме того,
4) сформировавшийся признак ограничен сверху экоценотическим пространством возможностей ареала вида.
Канализируя эволюцию, эти пространства возможностей в то же время значительно снижают мутационный груз, переводя ряд несинонимичных мутаций в режим отбора, близкий к нейтральному. Таким образом, хотя пространство возможностей и ограничивает изменчивость, в нем никогда не будет достигнута полная унификация: высококонсервативную часть (коровый район ССТФ, пространственная структура белка или РНК, «ядро» регулога) всегда будет сопровождать низкоконсервативный «шлейф».
Рассмотрим простейший контур с отрицательной обратной связью (ООС), регулирующей концентрацию белка. Параметр «отклонение концентрации от нормы» отслеживается регуляторным звеном ООС, меняющим скорость синтеза белка. Контуру безразличны причины отклонения от нормы. Таким образом, ООС сужает норму реакции, «обнейтраливая», выводя из-под отбора, любое изменение на нижележащих уровнях иерархии. (Колчанов, Шиндялов 1991). Аналогичным свойством обладают лимитирующие звенья — наиболее быстротекущие реакции (Демиденко и др., 2004).
Но такая унификация на высшем уровне означает неизбежное размывание гомологии на нижележащих уровнях иерархии за счет квазинейтральной эволюции. Эти данные хорошо согласуются с фактом неравномерности темпов морфофизиологической и молекулярной эволюции (Glazko et al., 2005; CSAC, 2005; Arbiza et al., 2006).
Палеонтологические и современные данные показывает, что гомологические ряды наиболее характерны для раннего периода эволюции филумов — периода «архаического разнообразия», когда четкий архетип не сформирован (Мамкаев, 1991), и периода анагенеза, когда архетип устойчив, но филум еще не впал в стазис (Рожнов, 2006). В первом случае радикал имеет небольшую левую часть и длинную правую часть, варьирующую от таксона к таксону. Во втором случае — радикал с выраженной левой частью и четким иерархическим соподчинением признаков (Рожнов, 2006). Чем сильнее ООС, тем уже норма реакции, следовательно, стабилизирующий отбор благоприятствует таксонам с ООС.
При дизруптивном или движущем отборе ситуация противоположна: преимущество получают таксоны без ООС, слом которых должен вести к выявлению всех ранее обнейтраленных мутаций (гиперманифестация изменчивости). Такой дестабилизирующий отбор экспериментально продемонстрирован школой Д. К. Беляева при доместикации лисиц (Беляев, 1983) и куньих (Трапезов, 2008). Таким образом, во-первых, любая мутация будет нейтральной лишь до тех пор, пока она не способна повлиять на регуляцию в генной сети10.
Во-вторых, при сломе регуляторного контура все накопленные мутации будут тестироваться отбором «оптом», случайно подобранными ансамблями. Такой режим отбора мы предлагаем назвать «адаптивной оптимизацией» в противовес классической схеме движущего отбора, когда мутации тестируются сразу после их появления. При «архаическом разнообразии» мы видим процесс такой оптимизации, а момент, когда в палеонтологической летописи формируется устойчивый архетип, указывает на формирование системы ООС.
Литература
Алёшин В. В., Владыченская Н. С., Кедрова О. С., Милютина И. А., Петров Н. Б. Вторичная структура шпильки 17 низшего многоклеточного животного Rhopalura ophiocomae (Mesozoa: Orthonectida) как пример «прерываемого равновесия» в эволюции 18S рибосомной РНК // Молекулярная биология. 1999. Т. 33. No 2. С. 319–329.
Анистратенко В.В. Гомеоморфия: суть явления и его значение для систематики и филогенетики (на примере брюхоногих моллюсков) // Вестник зоологии. 1998. Т. 32. No 1/2. С. 98–107.
Афонников Д. А., Колчанов Н. А. Консервативные особенности ДНК-связывающих доменов класса «гомеодомен», обусловленные коадаптивными заменами аминокислотных остатков // Доклады АН. 2001. Т. 380. No 5. С. 691–695.
Беляев Д. К. Дестабилизирующий отбор // Развитие эволюционной теории в СССР (1917– 1970 годы). Л. : Наука, 1983. C. 266–277.
Берг Л. С. Номогенез или эволюция на основе закономерностей // Труды по теории эволюции. 1922–1930. Л. : Наука, 1977. С. 95–311.
Вавилов Н. И. Закон гомологических рядов в наследственной изменчивости. Саратов : Губполиграфотдел, 1920. 16 с.
Вавилов Н. И. Линнеевский вид как система // Труды по прикладной ботанике, генетике и селекции. 1931. Т. 26. Вып. 3. С. 109–134.
Вавилов Н. И. Закон гомологических рядов в наследственной изменчивости. 2-е изд., перераб. и расшир. М. ; Л. : Сельхозгиз, 1935. 56 с.
Вавилов Н. И. Мировые центры сортовых богатств (генов) культурных растений // Избранные труды в 5 томах. Т. 5. М. : Наука, 1965. С. 108–119.
Вавилов Н. И. Научные основы селекции пшеницы // Избранные произведения в двух томах. Т. 2. Л. : Наука, 1967. С. 7–259.
Васильева Л. Н. Кризис проблемы вида: причины и следствия // Эволюционная биология: материалы II Международной конференции «Проблема вида и видообразование». Томск: Изд-во Томского гос. ун-та, 2002. Т. 2. С. 31–50.
Горлов И. П., Горлова О. Ю. Движущий отбор в ходе эволюции человека // Информационный Вестник ВОГиС. 2007. Т. 11. No 2. С. 363–372.
Грант В. Эволюционный процесс. М. : Мир, 1991. 488 c.
Дарвин Ч. Происхождение видов путем естественного отбора. СПб. : Наука, 1991. 539 c.
Демиденко Г. В., Колчанов Н. А., Лихошвай В. А., Матушкин Ю. Г., Фадеев С. И. Математическое моделирование регуляторных контуров генных сетей // Журнал вычислительной математики и математической физики. 2004. Т. 44. No 10. С. 1921–1940.
Заварзин А. А. Параллелизм структур как основной принцип морфологии // Изв. Биологич. науч.-исслед. ин-та при Пермском ун-те. 1923. Т. 2. Вып. 4. С. 135–140.
Заварзин А. А. Сравнительная гистология. СПб. : Изд-во СПбГУ, 2000. 518 с.
Заварзин Г. А. Несовместимые признаки в системе родов бактерий // Журнaл общей биологии. 1973. Т. 34. No 4. С. 530–538.
Захаров-Гезехус И. А. Проблема гомологии в эволюционной биологии. М. : [б. и.], 2008. 127 с.
Иванисенко В. А., Деменков П. С., Фомин Э. С., Крестьянова М. А., Ощурков И. С., Иванисенко Т. В., Иванисенко Н. В., Пинтус С. С., Яркова Е. Э., Степаненко И. Л., Сурнина Н. Ю., Колчанов Н. А., Гончаров С. С. Распознавание функциональных сайтов в пространственных структурах белков // Системная компьютерная биология. Новосибирск: Изд-во СО РАН, 2008. С. 231–245.
Камшилов М.М. Отбор в различных условиях проявления признака // Биологический журнал. 1935. Т. 4. No 6. С. 1005–1013.
Камшилов М. М. 1939. Отбор как фактор, меняющий зависимость признака от изменений внешних условий // Доклады АН СССР. 1939. T. 23. No 4. С. 361–364.
Камшилов М.М. Проявление признака и изменчивость. Эффект порога проявления // Доклады АН СССР. 1940а. Т. 26. No 6. С. 605–608.
Камшилов М.М. Изменчивость и проявление. Проблема нормального фенотипа // Доклады АН СССР. 1940б. Т. 29. No 3. С. 239–243.
Камшилов М.М. Эволюция биосферы. 2-е изд. М. : Наука, 1974. 256 с.
Колмогоров А.Н. Уклонение от формул Харди при частичной изоляции // Доклады АН СССР. 1935. Т. 3. No 7. С. 129–132.
Колчанов Н. А., Подколодная О. А., Игнатьева Е. В., Хлебодарова Т. М., Проскура А. Л., Суслов В. В., Воронич Е. С., Дубовенко Е. А. Интеграция генных сетей, контролирующих физиологические функции организма // Информационный Вестник ВОГиС. 2005. Т. 9. No 2. С. 179–198.
Колчанов Н. А., Шиндялов И. Н. Теоретическое исследование эволюции регуляторных контуров при различных типах отбора // Проблемы генетики и теории эволюции. Новосибирск : Наука, 1991. С. 268–279.
Мамкаев Ю. В. Методы и закономерности эволюционной морфологии // Современная эволюционная морфология. Киев : Наукова думка, 1991. С. 88–103.
Мейен С. В. О соотношении номогенетического и тихогенетического аспектов эволюции // Журнал общей биологии. 1974. Т. 35. No 3. С. 353–364.
Мейен С. В. Олигомеризация и полимеризация в эволюции древнейших высших растений // Значение процессов полимеризации и олигомеризации. Л. : Зоологический ин-т АН СССР, 1977а. С. 75–77.
Мейен С. В. Таксономия и мерономия // Вопросы методологии в геологических науках. Киев: Наукова думка, 1977б. С. 25–33.
Мейен С. В., Шрейдер Ю. А. Методологические аспекты теории классификации // Вопросы философии. 1976. No 12. С. 67–79.
Михайлова И. А., Бондаренко О. Б. Палеонтология. Т. 1. М. : Изд-во МГУ, 1997. 446 c.
Политов Д. В., Крутовский К. В. Клинальная изменчивость и интрогрессивная гибридизация в популяциях европейской и сибирской елей // Жизнь популяций в гетерогенной среде. Йошкар-Ола : Периодика Марий Эл, 1998. P. 78–89.
Рожнов С. В. Закон гомологических рядов Н. И. Вавилова и архаическое многообразие по данным палеонтологии // Эволюция биосферы и биоразнообразия. М.: Т-во научных изданий КМК, 2006. С. 134–147.
Розанов А. Ю. Закономерности морфологической эволюции археоциат и вопросы ярусного расчленения нижнего кембрия. М. : Наука, 1973. 164 с.
Соболев Д. Н. Начала исторической биогенетики. Симферополь : Государственное издательство Украины, 1924. 202 с.
Трапезов О. В. Гомологические ряды изменчивости окраски меха у американской норки (Mustela vison Schreber, 1777) в условиях доместикации // Информационный Вестник ВОГиС. 2008. Т. 11. No 3/4. С. 547–559.
Филипченко Ю. А. О параллелизме в живой природе // Успехи экспериментальной биологии. 1925. Т. 3. Вып. 3/4. С. 242–258.
Шаталкин А. И. Регуляторные гены в развитии и проблема морфотипа в систематике насекомых // Чтения памяти Н. А. Холодковского. Вып. 56(2). СПб. : Наука, 2003. 109 с.
Шмальгаузен И. И. Факторы эволюции. М. : Наука, 1968. 452 с.
Ananko E. A., Kondrakhin Y. V., Merkulova T. I., Kolchanov N. A. Recognition of interferon-inducible sites, promoters, and enhancers // BMC Bioinformatics. 2007. Vol. 8. P. 56.
Arbiza L., Dopazo J., Dopazo H. Positive selection, relaxation, and acceleration in the evolution of the human and chimp genome // PLoS Computational Biology. 2006. Vol 2. No 4. e38. Epub 2006. Apr 28.
Aspöck G., Kagoshima H., Niklaus G., Bürglin T.R. Caenorhabditis elegans has scores of hedgehog-related genes: sequence and expression analysis // Genome Research. 1999. Vol. 9. No 10. P. 909–923.
Babu M. M., Luscombe N. M., Aravind L., Gerstein M., Teichmann S. A. Structure and evolution of transcriptional regulatory networks // Current Opinion in Structural Biology. 2004. Vol. 14. No 3. P. 283–291.
Badiu C. Genetic clock of biologic rhythms // Journal of Cellullar and Molecular Medicine. 2003. Vol. 7. No 4. P. 408–416.
Beadle G. W., Tatum E. L. Genetic control of biochemical reactions in Neurospora // The Proccedings of the National Academy of Sciences of the USA. 1941. Vol. 27. No 11. P. 499–506.
Bergmann S., Ihmels J., Barkai N. Similarities and differences in genome-wide expression data of six organisms // PLoS Biology. 2004. Vol. 2. No 1. E9. Epub 2003. Dec 15, E9.
Briscoe A. D. Homology modeling suggests a functional role for parallel amino acid substitutions between bee and butterfly red- and green-sensitive opsins // Molecular Biology and Evolution. 2002. Vol. 19. No 6. P. 983–986.
CSAC — Chimpanzee Sequencing and Analysis Consortium. Initial sequence of the chimpanzee genome and comparison with the human genome // Nature. 2005. Vol. 437. No 7055. P. 69–87.
Chang B. S., Jönsson K., Kazmi M. A., Donoghue M. J., Sakmar T. P. Recreating a functional ancestral archosaur visual pigment // Molecular Biolology and Evolution. 2002. Vol. 19. No 9. P. 1483–1489.
Cook C. E., Yue Q., Akam M. Mitochondrial genomes suggest that hexapods and crustaceans are mutually paraphyletic //Proceedings of the Royal Society of London. Series B, Biological Sciences. 2005. Vol. 272. No 1569. P. 1295–1304.
Duret L., Mouchiroud D. Determinants of substitution rates in mammalian genes: expression pattern affects selection intensity but not mutation rate // Molecular Biology and Evolution. 2000. Vol. 17. No 1. P. 68–74.
Fayngerts S. A., Najakshin A. M., Taranin A. V. Species-specific evolution of the FcR family in endothermic vertebrates // Immunogenetics. 2007. Vol. 59. No 6. P. 493–506.
Garcia-Fernàndez J. Hox, ParaHox, ProtoHox: facts and guesses // Heredity. 2005. Vol. 94. No 2. P. 145–152.
Gehring W. J. New perspectives on eye development and the evolution of eyes and photoreceptors // Journal of Heredity. 2005. Vol. 96. No 3. P. 171–184.
Gehring W. J., Ikeo K. Pax 6: mastering eye morphogenesis and eye evolution // Trends in Genetics. 1999. Vol. 15. No 9. P. 371–377.
Glazko G., Veeramachaneni V., Nei M., Makałowski W. Eighty percent of proteins are different between humans and chimpanzees // Gene. 2005. Vol. 346. P. 215–219.
Guselnikov S. V., Ramanayake T., Erilova A. Y., Mechetina L. V., Najakshin A. M., Robert J., Taranin A. V. The Xenopus FcR family demonstrates continually high diversification of paired receptors in vertebrate evolution // BMC Evolutionary Biology. 2008. Vol. 8. P. 148.
Haldane D. B. The comparative genetics of colour in rodents and carnivore // Biological Review. 1927. Vol. 11. P. 199.
Huynen M. A., Snel B., Noort V. van Comparative genomics for reliable protein-function prediction from genomic data // Trends in Genetics. 2004. Vol. 20. No 8. P. 340–344.
Ivanisenko V. A., Pintus S. S., Grigorovich D. A., Kolchanov N. A. PDBSite: a database of the 3D structure of protein functional sites // Nucleic Acids Research. 2005. Vol. 33. Database issue. Р. D183–D187.
Kauffman S. A. Gene regulation networks: A theory for their global structure and behaviors // Current Topics in Developmental Biology. Vol. 6. N. Y. : Academic Press, 1977. P. 145–182.
Kuwabara P. E., Lee M. H., Schedl T., Jefferis G. S. A C. elegans patched gene, ptc-1, functions in germ-line cytokinesis // Genes and Development 2000. Vol. 14. No 15. P. 1933–1944.
Levitsky V. G., Ignatieva E. V., Ananko E. A., Turnaev I. I., Merkulova T. I., Kolchanov N. A., Hodgman T. C. Effective transcription factor binding site prediction using a combination of optimization, a genetic algorithm and discriminant analysis to capture distant interactions // BMC Bioinformatics. 2007. Vol. 8. P. 481.
Lowman A. C., Purugganan M. D. Duplication of the Brassica oleracea APETALA1 floral homeotic gene and the evolution of domesticated cauliflower // Journal of Heredity. 1999. Vol. 90. No 5. P. 514–520.
Malko D. B., Ermakova E. O. Evolution of splicing in insects // Proceedings of the MCCMB 2007. Moscow: MSU Press, 2007. P. 193–194.
Meyen S. V. Plant morphology in its nomothetical aspects // The Botanical Review. 1973. Vol. 39. No 3. P. 205–260.
Meyen S. V. Parallelism and its significance for the systematics of fossil plants // Geophytology. 1971. Vol. 1. No 1. P. 34–47.
Ogishima S., Tanaka H. Missing link in the evolution of Hox clusters // Gene. 2007. Vol. 387. No 1–2. P. 21–30.
Palatnik J. F., Allen E., Wu X., Schommer C., Schwab R., Carrington J. C., Weigel D. Control of leaf morphogenesis by microRNAs // Nature. 2003. Vol. 425. No 6955. P. 257–263.
Pauli F., Liu Y., Kim Y. A., Chen P. J., Kim S. K. Chromosomal clustering and GATA transcriptional regulation of intestine-expressed genes in C. elegans // Development. 2006. Vol. 133. No 2. P. 287–295.
Politov D. V. Patterns of reticulate evolution in plants and animals of the boreal zone // II International Conference “Biosphere Origin and Evolution”. Abstracts book. Novosibirsk : Boreskov Institute of Catalysis SB RAS. 2007. P. 69.
Regier J. C., Shultz J. W., Kambic R. E. Pancrustacean phylogeny: hexapods are terrestrial crustaceans and maxillopods are not monophyletic // Proceedings of the Royal Society of London. Series B, Biological Sciences. 2005. Vol. 272. No 1561. P. 395–401.
Rooney A. P., Zhang J., Nei M. An unusual form of purifying selection in a sperm protein // Molecular Biology and Evolution. 2000. Vol. 17. No 2. P. 278–283.
Ryan J. F., Mazza M. E., Pang K., Matus D. Q., Baxevanis A. D., Martindale M. Q., Finnerty J. R. Pre-bilaterian origins of the Hox cluster and the Hox code: evidence from the sea anemone, Nematostella vectensis // PLoS ONE. 2007. Vol. 2. No 1. P. e153.
Sax C. M., Cvekl A., Kantorow M., Gopal-Srivastava R., Ilagan J. G., Ambulos N. P., Piatigorsky J. Lens-specific activity of the mouse alpha A-crystallin promoter in the absence of a TATA box: functional and protein binding analysis of the mouse alpha A-crystallin PE1 region // Nucleic Acids Researsh. 1995. Vol. 23. No 3. P. 442–451.
Sheldon P. R. Parallel gradualistic evolution of Ordovician trilobites // Nature. 1987. Vol. 330. No 6418. P. 561–563.
Snel B., Noort V. van, Huynen M. A. Gene co-regulation is highly conserved in the evolution of eukaryotes and prokaryotes // Nucleic Acids Research. 2004. Vol. 32. No 16. P. 4725–4731.
Stedman H. H., Kozyak B. W., Nelson A., Thesier D. M., Su L. T., Low D. W., Bridges C. R., Shrager Minugh-Purvis N., Mitchell M. A. Myosin gene mutation correlates with anatomical changes in the human lineage // Nature. 2004. Vol. 428. No 6981. P. 415–418.
Stuart J. M., Segal E., Koller D., Kim S. K. A gene-coexpression network for global discovery of conserved genetic modules // Science. 2003. Vol. 302. No 5643. P. 249–255.
Swanson W. J., Vacquier V. D. The rapid evolution of reproductive proteins // Nature Reviews Genetics. 2002. Vol. 3. No 2. P. 137–144.
Taddei I., Morishima M., Huynh T., Lindsay E. A. Genetic factors are major determinants of phenotypic variability in a mouse model of the DiGeorge/del22q11 syndromes // The Proccedings of the National Academy of Sciences of the USA. Vol. 98. No 20. P. 11 428–11 431.
Teichmann S. A., Babu M. M. Gene regulatory network growth by duplication // Nature Genetics. 2004. Vol. 36. No 5. P. 492–496.
Vanselow K., Vanselow J. T., Westermark P. O., Reischl S., Maier B., Korte T., Herrmann A., Herzel H., Schlosser A., Kramer A. Differential effects of PER2 phosphorylation: molecular basis for the human familial advanced sleep phase syndrome (FASPS) // Genes and Development. 2006. Vol. 20. No 19. P. 2660–2672.
Wyckoff G. J., Wang W., Wu C. I. Rapid evolution of male reproductive genes in the descent of man // Nature. 2000. Vol. 403. No 6767. P. 261–263.
Xia K., Xue H., Dong D., Zhu S., Wang J., Zhang Q., Hou L., Chen H., Tao R., Huang Z., Fu Z., Chen Y. G., Han J. D. Identification of the proliferation/differentiation switch in the cellular network of multicellular organisms // PLoS Computational Biology. 2006. Vol. 2. No 11. P. 145.
Xu Y., Padiath Q. S., Shapiro R. E., Jones C. R., Wu S. C., Saigoh N., Saigoh K., Ptácek L. J., Fu Y. H. Functional consequences of a CKIdelta mutation causing familial advanced sleep phase syndrome // Nature. 2005. Vol. 434. No 7033. P. 640–644.
Zugasti O., Rajan J., Kuwabara P. E. The function and expansion of the Patched- and Hedgehog-related homologs in C. elegans // Genome Research. 2005. Vol. 15. No 10. P. 1402–1410.
«Чарльз Дарвин и современная биология«. Труды Международной научной конференции 21–23 сентября 2009 г. СПБ: Нестор-история, 2010. С.61-75.
Примечания
1 Н. И. Вавилов (1967) разрабатывал, но не закончил классификацию признаков злаков. В настоящее время гомологические ряды наиболее широко используют палеонтологи. Данные по растениям (Мейен, 1974; 1977 а, б; Meyen, 1971, 1973) и бентосным морским организмам (Анистратенко, 1998; Рожнов, 2006; Sheldon, 1987) говорят, что наиболее четко ряды выявляются на уровне рода, а не вида.
2 Ч. Дарвин (1991) отличал неопределенную изменчивость от «спорта» (садоводческий термин) — внезапного, случайного и константного в дальнейшем ряду поколений изменения признака (то, что позднее Г. де Фриз назовет «мутацией»). Неопределенная изменчивость — признаки с неустойчивым, варьирующим в ряду поколений и от особи к особи проявлением. Отбор фиксирует ее, кумулятивно ограничивая степень варьирования, тогда как константные «спорты» отбор либо выбраковывает, либо комбинирует их сочетания.
3 Аналитически строго это показал А. Н. Колмогоров (1935).
4 Первоначально таблица содержала 140 родов, в настоящее время — более 200 (Рожнов, 2006).
5 Или группы близких видов. Молекулярно-филогенетические исследования близких, длительно викарирующих видов свидетельствуют о неполной репродуктивной изоляции, что ведет к эпизодическому обмену генами (вследствие кроссинговера за счет значительных участков гомологии и обратного скрещивания гибридов и родителей). См.: Политов, Крутовский, 1998; Politov, 2007.
6 В современной литературе для внутренней структуры таксона (число субтаксонов) принят термин «экстенсионал», тогда как отличительные признаки таксона (его архетип) образуют его интенсионал (Мейен, Шрейдер 1976; Васильева, 2002).
7Даже если ГС не используется, полной потери не происходит. Так, в Hh-каскаде нематод утрачен морфоген Hh, один из его рецепторов — Smo, но сохранен другой рецептор — Ptс. То есть, от ГС Hh-каскада остался только один из двух модулей передачи сигнала через цитоплазму на ядро. Какой сигнал принимает Ptс пока неясно, хотя у нематод выявлены Hog- белки, кодируемые семейством генов, гомологичных Hh (Aspöck et al., 1999, Kuwabara et al., 2000, Zugasti et al., 2005).
8 Возможен также перебор пространства возможностей на уровне субъединиц белка. Например, таким образом в разных таксонах независимо сформировались FcR-рецепторы иммунной системы (Fayngerts et al., 2007; Guselnikov et al., 2008).
9 Так как протамины играют важную роль и в оплодотворении, движущий отбор, фиксируемый кимурианским критерием, видимо, играет роль в двух противоположных трендах эволюции: изменяя белок, формирует репродуктивный барьер (Swanson, Vacquier, 2002) и в то же время путем коадаптивных замен «следит», чтобы общие параметры белка не слишком флуктуировали (Rooney et al., 2000).
10 Вывод подтвержден выявлением ненулевой относительной вредности синонимичных замен у человека (вредность — недопредставленность в геноме мутаций данного класса по сравнению с теоретически возможным). Авторы (Горлов, Горлова, 2007) объясняют это влиянием синонимичных замен на сайты сплайсинга, играющие важную регуляторную роль, что согласуется с данными (Malko, Ermakova, 2007) по дрозофиле.
Рекомендуем прочесть
 В защиту изучения коми языка
В защиту изучения коми языка Когнитивный диссонанс: теория с социальными приложениями
Когнитивный диссонанс: теория с социальными приложениями Положение женщины и патриархальная реакция в Центральной Азии
Положение женщины и патриархальная реакция в Центральной Азии О борьбе в науке
О борьбе в науке Высокие цены на продовольствие не зависят от урожая
Высокие цены на продовольствие не зависят от урожая Анализируя пальцевый индекс, мы «говорим со стенкой»
Анализируя пальцевый индекс, мы «говорим со стенкой» За пределами роста-3
За пределами роста-3 Нервная булимия — болезнь открытого общества
Нервная булимия — болезнь открытого общества
