 упускаемая возможность")

Заказник «Юнтоловский», водосбор Лахтинского разлива. Из: «Атлас ООПТ Санкт-Петербурга«, с.22
Резюме. Современный город имеет неожиданные потенции «заказника», в том смысле что сохранение там биоразнообразия региона, видов и/или сообществ (скажем, восстановленных из техногенных аналогов) может быть эффективнее, чем вовне. Некоторые из них (сохранение бабочек, других опылителей) в травянистых сообществах уже очевидны, и люди отстаивают их. Другие, следующие из возвратной урбанизации «диких» видов, пока неявственны; они обсуждаются в настоящей статье на орнитологическом материале.
Показывается:
1) лишь этот процесс «даёт шанс» сохранения редких и уязвимых видов в староосвоенных регионах Европы, северной Америки, Японии и других фрагментов «мирового города»;
2) при нынешней (экстремальной) степени трансформации «традиционного с/х ландшафта» региона развитием городских ареалов в составе агломераций «шанс» всё более актуален и для обычных/многочисленных видов, чьи популяции делаются неустойчивыми.
В статье обсуждается:
а) в чём состоит этот шанс, почему «диким» видов фауны (флора изучена менее, но можно предполагать сходные тенденции) урболандшафты создают преимущества, отсутствующие за их пределами — как на антропогенно-трансформированных территориях (скажем в сельхозугодьях), так и природных, относительно малонарушенных,
б) чем последние «выгодны» для осваивающих её видов (а в староосвоенных регионах – и спасительны), как они делают возможным урбанизацию всех видов региональной фауны,
б) как приспособления, связанные с освоением, выводят их «из-под» воздействия «островного эффекта»,
в) насколько уязвимые, консервативные и специализированные виды птиц «пользуются» этим,
г) что может – и должен! – сделать человек в плане экообстройства «техногенных аналогов» исходных местообитаний таких видов, чтобы «переселение» стало успешным. Также обсуждаются парадоксы городских экосистем, позволяющие в охране видов – но не сообществ — частично «уйти» от обычных ограничений территориальной охраны природы.
«Бабочки могут украшать город так же как птицы…»
Содержание
- 1 «Бабочки могут украшать город так же как птицы…»
- 2 Эволюция городских систем: агломерация, опорный каркас расселения, мегалополис
- 3 Метапопуляции в «островном ландшафте»: «прыжки» с «льдины на льдину»
- 4 Возвратная урбанизация редких видов — последний «шанс на спасение» в староосвоенных регионах
- 5 Формирование городской авифауны: баланс «пополнения» и «исчезновения»
- 6 Плюсы урбанизации для «диких» видов птиц
- 7 Парадоксы городских экосистем, реализующие «преимущества»
- 8 Литература
Современный город не только «теснит», «дробит» и «пережёвывает» природные ландшафты, почему «дикая природа», как виды, так и сообщества представляются полными антиподами, в т.ч. и в пространстве («центр vs периферия» урбанизирующегося региона). Однако это неверно: у глобально идущей урбанизации есть также «светлая сторона», которую природоохранникам грех не использовать, тем более успехи использования исключительно полезны всем городским обывателям (Фридман, 2012а). Чем дальше, тем больше города потенциально способны из «техногенных пустынь» стать местообитанием, привлекающим «дикие» виды растений и животных — особенно тех, чьи региональные биотопы уже сейчас нарушаются и разрушаются урбанизацией. Или даже «заказником», сохраняющим редкие и уязвимые виды лучше, чем «архипелаги» природных сообществ в урбанизированных регионах.
Некоторые возможности этого очевидны, и люди используют их уже сейчас. Так, пустыри и газоны, если их не косить (или специально формировать как разнотравные, противостоящие английским стриженым) представляют собой техногенные аналоги суходольных лугов. Они сохраняют бабочек, одиночных пчёл, других опылителей, многих хортобионтов лучше, чем однотипные ландшафты за городом, т. к. там при ведении с/х широко используются пестициды (Волкова, Бейко, 1998; Волкова, 2000).
То же верно и в отношении луговой флоры, большинство видов которой уязвимы, а то и угрожаемы при интенсивном ведении сельского хозяйства, с удобрением и мелиорацией; это плата за кратное увеличение урожайности кормовой травы (Разран, 2006). Для сохранения этих видов в Швеции, Германии, США, других развитых странах а) ревитализируют малые реки в черте города, с восстановлением «близкой к природной» мозаики пойменных сообществ[1] (Rasran, Vogt, 2018; Kiehl et al., 2010; Крамер и др., 2010; Ryan, Jensen, 2008; Rasran et al., 2006); б) возникло движение Sustainable Gardening, использующее в озеленении местные растения и имитирующее исходную растительность вместо обычных зелёных насаждений, линейных и однородных, с конечной целью наибольшего приближения к исходным ландшафтам до их урбанистической трансформации[2].
Другие потенции «города как заказника» лишь сейчас проявились во всей полноте, почему заметны существенно хуже и менее обсуждаются в обществе, почему многие не подозревают, что упускают их без борьбы (а с ними — и право на здоровую среду обитания, декларированное ст. 42 Конституции РФ). Они следуют из возвратной урбанизации «диких» видов птиц, млекопитающих, других групп биоты и обсуждаются в настоящей статье.
У первых двух она изучается давно и подробно, у других здесь сделаны только первые шаги (в т.ч. у сосудистых растений, где ряд реликтовых или угрожаемых видов успешно осваивает т. н. замещающие местообитания, как правило техногенные[3], Быченко, 1992; Чхобадзе, Филиппов, 2013; Фридман, 2017: 175-178).
Поэтому обсуждение природоохранных потенций урболандшафтов мы проведём на орнитологическом материале. Тем более именно птицы раз за разом оказываются отличной модельной группой для решения общебиологических проблем, особенно в эволюционной экологии и микроэволюции, исчерпывающих большинство проблем «взаимодействия» видовых популяций с урболандшафтами региона.
Эволюция городских систем: агломерация, опорный каркас расселения, мегалополис
«Россыпи» поселений первых связаны с характерным «кружевом» местообитаний — тем жёстче и специфичней, чем более специализирован вид, консервативней его жизненная стратегия. Вторые же закономерно 1) «растут» (Стольберг, 2000: 22-23; https://en.wikipedia.org/wiki/Urban_sprawl; Ewing, 2008; Ewing et al., 2008) и 2) «собираются в стаи», т. е. формируют системы взаимозависимых и соподчинённых друг другу городов — агломерации, где более периферийные города координированно растут сообразно росту «ядра». Дальше агломерации соединяются полимагистралями в образования более высокого уровня — опорный каркас расселения, начиная расти навстречу друг другу, а не в стороны и т. д. (Barry, 2008; Burgess, 2008).
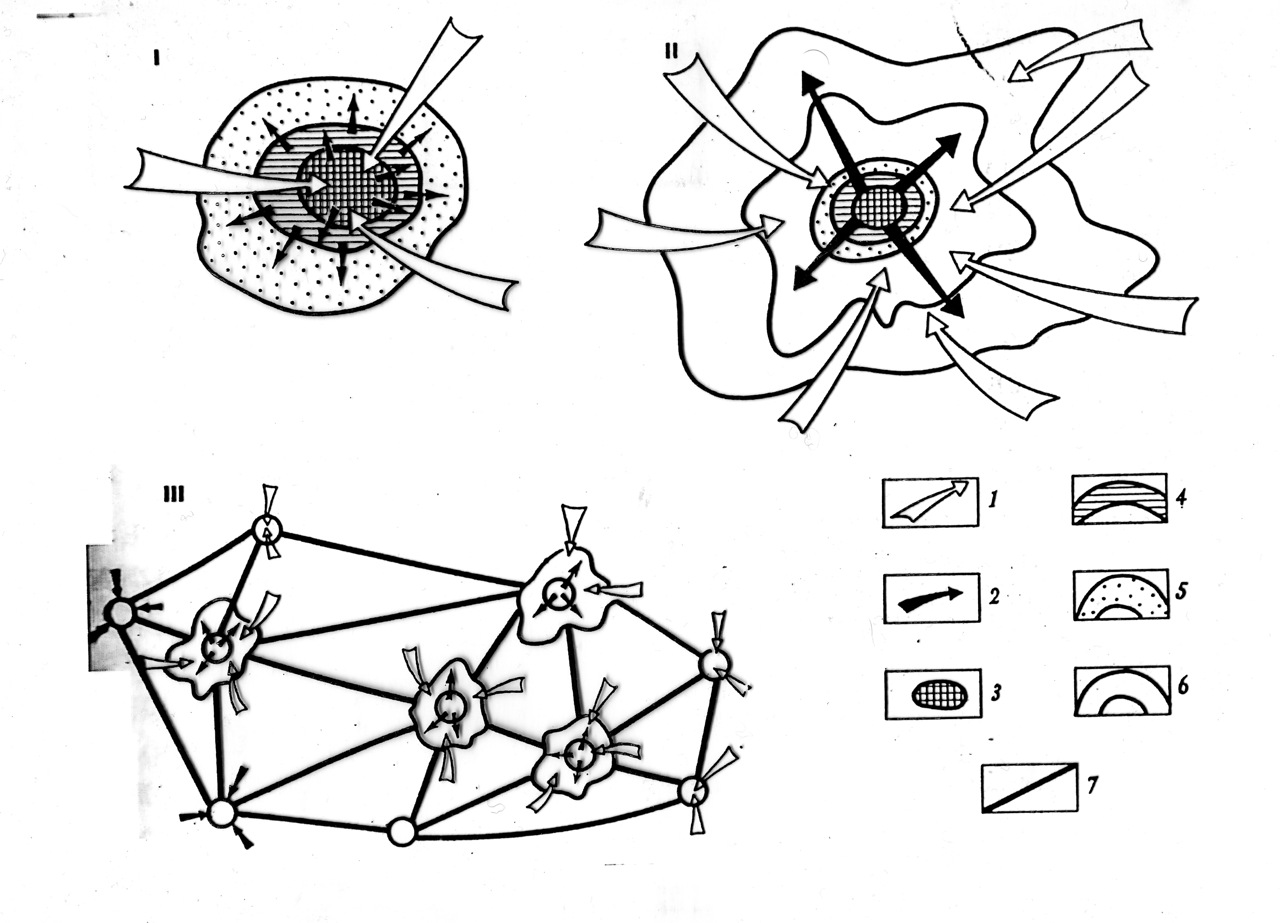
Природные ландшафты по ходу вышеописанного либо «фагоцитируются» растущими городами, заключаясь в «ядро» или один из подчинённых городских ареалов. Либо превращаются в «прослойки», «зажатые» между «кольцами» городов развивающейся агломерации, отчасти подобной Солнечной системе, с перспективой последующего «дробления» на всё более изолированные «острова» (Лаппо, 1997, цит.по: Фридман и др., 2016, также рис. 2 там же).
Развитие объединений на уровне «выше города» идёт так закономерно и единообразно на всех континентах, что может описываться общей для них эволюционной моделью урбанизации (Richardson, 1980; Klaassen, 1981; Лаппо, 1983; Кюммель, 1987; Полян, 1988; Geyer, 1993, цит.по: Фридман и др., 2016). Разнятся лишь давность начала индустриализации и модус последней; в плане нашей нами проблемы «города как заказника» также важны различия между урбанизацией в странах с плановой экономикой (СССР и другие «коммунистического блока») и их антиподах с рыночной (США и другие развитые страны).
В первом случае образуется т. н. поляризованный ландшафт, исключительно ценный для сохранения биоразнообразия. И, самое главное, он устойчиво сохраняется при увеличении людности, уровня промышленного и транспортного развития «ядер» и/или подчинённых городов в составе агломераций, если специально отслеживать и своевременно пресекать «покушения» на него в частных интересах — что масс рекреантов, что министерств и ведомств (Родоман, 2002: 18-48, 115-118, 202-210, 210-219; Фридман, 2017: 415-432).
Во втором (развитые страны с рыночной экономикой) — этот ландшафт закономерно разрушается уже в 1950-60-е гг., когда
1) во всём мире города из внемасштабных образований делаются массивами, отображаемыми в масштабе карты, сравниваются площадью с массивами лесов, лугов и болот региона, а потом и превосходят их.
2) также как в развитых странах с плановой экономикой (СССР, ГДР, ЧССР, ВНР) возникает положительная связь между людностью города и общим временем, проводимым жителями на природе (Фридман и др., 2016; Фридман, 2017).
Это стремление жителей более крупных городов больше времени проводить на природе не может не нарушать естественные экосистемы, фрагментировать крупные массивы и т.п.: пресс «отдыхающих и гуляющих» дополняется их расчленением сетью автодорог при одновременной инсуляризации изнутри дорожно-тропиночной сетью. Далее, всякое учащение посещений дикой природы увеличивает вероятность приобретения горожанами дачных участков, первое и второе стимулирует дальнейшее развитие дорожной сети, она доставляет «на природу» ещё больше отдыхающих и т.д. Круг замыкается[4].
3) хотя вроде бы люди с периферии концентрируются в крупных городах, во всех урбанизированных регионах планеты площадь городов растёт намного быстрей населения (Стольберг и др., 2000). Иными словами, за 200 лет городского строительства, в современных условиях индустриализации и научно-технического прогресса люди не смогли рационально использовать пространство. Всё большая часть функций современного города не может выполняться только на его территории и требует всё большей территории региона, включая понедельный и годовой отдых горожан[5].
Это значит, что рекреационные нарушения, составляющие важную (иногда критически) часть «давления» городских ареалов на коренные сообщества региона, «распространяются» в пространстве через дорожную сеть и «пятна» садово-дачной застройки, появляющиеся на концах «ветвей». Они «растут» пропорционально городскому развитию; «коэффициент пропорциональности» тем выше, чем развитей пригороды с коттеджной застройкой (suburbs), чем больше оттуда и из собственно города люди на уик-энд ездят частным автотранспортом, а не ж/д, и чем более автозависимы города (Никифоров, 2002; Вучик, 2011; Laurance et al., 2014). Вследствие всего вышеописанного города «расплываются» как масляное пятно на бумаге, вместо «фрактальной дифференциации» поляризованного ландшафта, сохраняющего связность природных территорий при урбанизации — правда, лишь при наилучшем развитии событий.
А географ Б.Б.Родоман, выделивший последний как идеальный объект, инвариантно проявляющийся в развитии разных регионов нашей страны, при всех местных особенностях природы или хозяйства, фактически первооткрыватель необходимости экосети (econet) для сохранения дикой природы в староосвоенных регионах. Последняя акцентирует важность связности и непрерывности «островов» природных территорий, находящихся в «окружении» урболандшафтов, с/х угодий, карьеров, гарей, вырубок и других «пятен» нарушения и трансформации» (Хански, 2010: 264-277; Arponen et al., 2014; Klenke et al., 2014). См. быстрый прогресс их мозаики в Подмосковье вследствие рыночных реформ (Blinnikov et al., 2006; Marzluff, Ewing, 2008; Карпачевский и др., 2009).
Возвратная урбанизация «диких» видов (не только обычных, но также редких и уязвимых, и даже находящихся под угрозой исчезновения) замечательна следующим. По мере её протекания вид чем дальше, тем больше «освобождается» от зависимости между жизнеспособностью популяций и связностью занятых им «островов» местообитаний. А это исключительно важно для его сохранения — ведь имеющаяся экосеть что в Европе, что в Подмосковье оказывается не вполне сформированной, не очень обеспечивает охрану видов и природных ландшафтов — тем хуже, чем жёстче последние привязаны к ним, чем более индикаторны для коренных сообществ и пр. См. пример с ключевыми видами бабочек на торфяных болотах Мещёры (Бутовский и др., 2001; Butovsky et al., 2003), или с ключевыми для лесных сообществ видами дятловых птиц, особенно специализированных, в лесных фрагментах на градиенте урбанизации в разных странах Европы (Mikusinski, 1998; Myczko et al., 2014).
Метапопуляции в «островном ландшафте»: «прыжки» с «льдины на льдину»
Последнее подрывает жизнеспособность видовых популяций в регионе, последовательно превращая их из «материка» в «архипелаг», всё более и более «неудобный» для воспроизводства популяционных «группировок» видов, «запертых» на «островах». Это т.н.метапопуляции: их изучение также стало отдельной дисциплиной — metapopulation ecology (см. https://www.helsinki.fi/en/researchgroups/metapopulation-research-centre; https://en.wikipedia.org/wiki/Metapopulation).
В чём их отличие от популяций без «мета-», свойственных нефрагментированным местообитаниям «материка»? Последние связаны — непосредственно краями мозаик в «лоскутное одеяло» природных сообществ, или соединены «коридорами» в виде пойм малых рек, опушек «лоскутов» и других направляющих линий исходного ландшафта. Благодаря им на «материке» видовые популяции реализуют свою пространственно-этологическую структуру «свободно», поток переселенцев из «подвижного резерва», связывающий местные группировки в системное целое, и интенсивен, и значителен во времени, обеспечивая возможность компенсации численности и прочей саморегуляции (Щипанов, 2003, 2016; Фридман, 2012б).
Конкретная конфигурация группировок здесь задана преимущественно сигнальным полем, структура которого в каждой местности как бы «оценивает» ландшафты «с т.з. вида», формируя «градиенты притяжения» и «градиенты страха», свободно сортирующие птиц между биотопами разного качества[6]. И наоборот, «островной ландшафт», созданный антропогенной фрагментацией местообитаний, «укладывает» поселения вида в «прокрустово ложе» наличных «островов», при затруднённости перемещений между ними в поисках территории и партнёра. Вероятность оказаться в среде, наиболее подходящей данной особи, с её индивидуальной стратегией и поведенческими потенциями, здесь существенно ниже, чем в первом случае. Грубо говоря, на «архипелаге» на наиболее подходящий «остров» птицы скорее «случайно наталкиваются» (с тем меньшей вероятностью, чем он меньше, изолированней от других, а пространство между ними — матрикс — хуже проницаемо для особей этого вида), также как присоединиться к наиболее привлекательным группировкам, тогда как на «материке» это закономерный процесс, устойчивый во времени и пространстве.
Поэтому природоохранники создают акустические и визуальные стимулы, «осаживающие» население разных видов видов на «островах», наиболее подходящих для размножения, если те изолированы внутри труднопроходимого для них ландшафта, (например, города) или способствующие концентрации населения исчезающих видов на «отдельных «островах» ООПТ. Скажем, обзор 24 современных исследований использования вторичных сигналов для привлечения конспецификов показал, что успех был достигнут в 20 случаях (Ahlering et al., 2010). Так, искусственное проигрывание песни успешно использовали для осаживания мигрантов исчезающей овсянки Ammodramus maritimus mirabilis во Флориде именно на охраняемых участках травянистых местообитаний в приморских маршах (Virzi et al., 2012). Аналогичный успех был достигнут для близкого вида — саванной овсянки A.savannarum (Andrews, 2013), как и для исчезающего черноголового виреона Vireo atricapilla. Там проигрыванием видоспецифических вокализаций на 7 экспериментальных участках с подходящими местообитаниями осадили 73 птицы в 2001 и 75 в 2002 г., многие из которых образовали пары и загнездились (Ward, Schlossberg, 2004). Тот же подход использовал С.П.Харитонов (2014) в восстановлении известной колонии озёрных чаек Larus ridibundus на оз.Киёво.
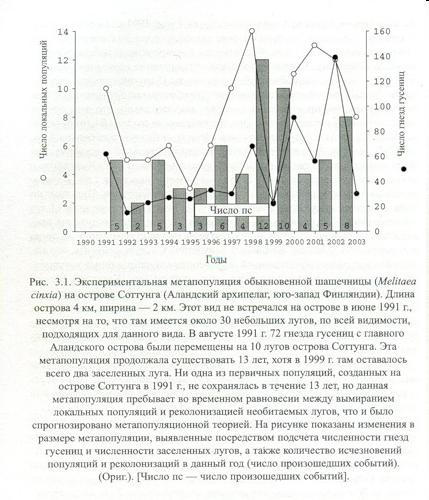
Отсюда жизнеспособность метапопуляций на «архипелагах» снижена обратно пропорционально размеру и прямо — степени изолированности «островов». Воспроизводство локальных поселений вида здесь в той же степени затруднено, т. к. оказывается неустойчивым. В отсутствие биотопических изменений поселения «материка» более-менее постоянны (по месту в сети поселений, хотя и колеблются численно), но на «архипелаге» они «вымирают» от случайных причин и вновь восстанавливаются реколонизацией. При обитании на «архипелаге» виды как бы «прыгают» с «льдины» на «льдину», и каждый «прыжок» создаёт риск вымирания. См. классический опыт И.Хански (2010: 120-126) с искусственно созданной метапопуляцией обыкновенной шашечницы Melitaea cinxia — угрожаемым видом Западной Европы — на 10 изолированных лугах о.Соттунга (Аландский архипелаг) из 30 имевшихся. Её численность в 1991-2003 гг. была нестабильна — сильно варьировала при примерно одном среднем значении. Ежегодно фиксировали от 2 до 12 случаев локального вымирания и пковторного заселения биотопов, и в 1999 г. она была на грани вымирания — вид обитал лишь на 2 лугах из 30 (рис. 3.1. там же).

Обыкновенная шашечница Melitaea сinxia
Согласно моделям «островной биогеографии», безусловно справедливым для метапопуляций, риск вымирания, следующий из их неустойчивости, тем больше, чем меньше по площади, сильней изолированы друг от друга «острова» и более случайным образом размещены в пространстве «архипелага». Неустойчивость же следует из «островного эффекта». Существование каждого из поселений вида на «острове», как и каждого вида в составе фауны (флоры) «архипелага» складывается как равновесие независимо идущих стохастических процессов вымирания и реколонизации (Хански, 2010; Акатов, 2012). В противоположность «материку», где то и другое «гарантировано» потоком переселенцев из «подвижного резерва», тем более мощным численно и устойчивым во времени, чем выше плотность населения вида и сохраннее «кружево» местообитаний (Фридман, 2012в; Фридман и др., 2016).
Но нет худа без добра: тот же самый процесс развития и соединения агломераций одновременно создаёт разного рода техногенные аналоги (ликвидируемых) природных местообитаний, которые виды пробуют осваивать, чтобы сохраниться в соответствующем регионе. Уничтожаемые луга «компенсируются» пустырями, обочинами дорог и другими лугоподобными участками в городе; болота – техногенными заболачиваниями и проточными подтоплениями в городе; старовозрастные леса – старыми парками и городскими лесами около них, водноболотные местообитания – прудами рыбхозов, отстойниками, полями орошения, затапливаемыми карьерами. Сюда же включаются элементы озеленения, от внутриквартальных фрагментов или куртин до заложенных парков, по мере старения деревьев, развития прочей растительности и пр. При минимальном экообустройстве «аналоги» превращаются в рефугиумы фауны и даже делаются «накопителями» редких видов (Авилова, 1997, 1998; Авилова, Ерёмкин, 2000, цит.по: Фридман, Ерёмкин, 2009; Фридман, 2017).
Поэтому важное направление охраны природы на староосвоенных территориях — «помощь» естественно идущим процессам урбанизации «диких» видов, их привлечение в техногенные аналоги природных местообитаний (от старых парков и разнотравных газонов до прудов водоподготовки), через экообустройство последних (Marzluff, Ewing, 2008). Страны с наименьшей устойчивостью популяций обычных видов – Великобритания, ФРГ, ГДР – в 1970-80-х гг. стали лидерами развития техники этой «помощи», экообустройства «аналогов» для привлечения туда птиц, амфибий, млекопитающих и пр., чтобы максимально приблизить к исходным сообществам. В 2000-е годы британские орнитологи так «вытащили» более половины видов птиц, находившихся на грани вымирания (Фридман, 2017: 106).
Сегодня необходим мониторинг обычных птиц и других групп биоты, пусть даже благополучных на данный момент. Преобразованность ландшафта в развитых странах такова, что неустойчивы даже обычные виды: они могут неожиданно увеличиться численно, занять новые районы или, наоборот, сократить численность и ареал, см тренды динамики в 1980-2006 гг.», http://www.ebcc.info/index.php?ID=358.

Виды Красной Книги г.Москвы, предпочитающие умеренное нарушение и исчезающие с его прекращением. 1. Пальчатокоренник балтийский Dactylorhiza báltica,

2. Подлесник европейский Sanicula europaea.

3. Истод большой Polygala major.

4. Тайник овальный Listéra ováta,

5. Дремлик болотный Epipactis palustris
Возможность вымирания видов, не имеющих хозяйственной ценности и процветавших при прежнем умеренном нарушении, показывает, что антропогенная трансформация природных ландшафтов подходит к критическому рубежу. За ним неизбежна потеря большей части прежнего биоразнообразия, и с нами останется лишь «серая биота». В развитых странах Европы и США прилагают отчаянные усилия, чтобы от него отойти, сохранив ключевые виды-регуляторы местных экосистем, от хищных птиц и млекопитающих (Mikusinski, Angelstam, 2004; Chapron et al., 2014) до дятлов с комплексом видов беспозвоночных и грибов, утилизующих мёртвую древесину в т. н. «посмертии дерева» (Mikusinski et al., 2001; Bobiek et al., 2005). И, конечно, оставшиеся «острова» природных ландшафтов «на всём протяжении» градиента урбанизации — как крупные, относительно ненарушенные на периферии региона, так и мелкие, часто нарушенные и преобразованные «с краёв» в его центре (Соболев, 1998; Соболев и др., 2006 (цит.по: Фридман и др., 2016); Shafer, 2008; Elbakidze et al., 2013)[7].
И именно урбанизация «диких» видов, происходящая спонтанно, закономерно, захватывающая все группы видов региональной (ави)фауны, в т.ч. почитавшиеся урбофобами, делает эти усилия успешными (местами — так даже очень) и одновременно сокращает затраты на их проведение, т. к. природа сама «делает тоже самое забесплатно (Balmford et al., 2002; Dearborn, Kark, 2010). Даже без «содействия» человека в форме техногенных аналогов прежних местообитаний виды региональной авифауны не только «отступают» на «материк», пока он остался на периферии (но быстро «тает», см. Фридман и др., 2016, рис.6).
Часть населения вида «движется» противоположно, с «архипелага» осваивает урболандшафты, сперва поселяясь на меньших по размеру и/или куда больше изменчивых во времени и пространстве «островах» природных территорий, «захваченных» городом внутрь или расчленённых и трансформированных распространением его влияния в ближних пригородах. Это и есть урбанизация (вида, в отличие от урбанизации населения, превращения сельчан в горожан и/или урбанизации территории, последовательного расширения городских ареалов и подчинения пригородных местностей динамике, свойственной урболандшафтам, вместо собственной, бывшей прежде, Фридман, Ерёмкин, 2009; Фридман и др., 2017).
Виды с лабильной стратегией делают это немедля. Более консервативные виды того же рода и/или гильдии[8] урбанизируются «с задержкой»: сперва «отступают» из «тающей», «рвущейся» сети местообитаний на «материк», т.ч. их численность в регионе падает, а ареал всё больше смещается кнаружи, на малонарушенную периферию. Но в какой-то момент (увы, чаще всего «пропускаемый» натуралистами) они поворачивают «на 1800», и вновь осваивают оставленный несколько лет или десятилетий назад «архипелаг» местообитаний, уже находящийся в структуре или под управляющим воздействием урболандшафта.
Это и есть возвратная урбанизация, сперва обнаруженная у обычных видов птиц: специализированных видов дендрофильной авифауны, «возвращающихся» в «острова» городских лесов, территориально консервативных видов хищных птиц, осваивающих лесные микрофрагменты в открытых (обычно сельскохозяйственных) ландшафтах и т. д. (Фридман, Ерёмкин, 2009; Галушин и др., 1998; Захарова, 2001, цит.по: Фридман и др., 2016). Самое замечательное, что такие же точно процессы, только с большей «задержкой» и длительным «отступлением» на периферию (30-60 лет, иногда более) фиксируются у всё большего числа редких, уязвимых и находящихся под угрозой исчезновения видов.

Средний дятел Dendrocopos medius — один из урбофобов, начавший возвратную урбанизацию в 1980-х гг. в Европе и в 2000-х в центре Европейской России
В т.ч. тех, кого раньше считали сугубыми урбофобами, жёстко связанными с крупными массивами малонарушенных широколиственных или бореальных лесов, торфяных болот, степей и других «остатков» природных ландшафтов, в староосвоенных регионах немногочисленных и далеко разнесённых друг от друга. Обзор и анализ таких случаев см. Фридман и др., 2007, 2016. Схемы прямой урбанизации лабильных видов и возвратной — более консервативных показаны на рис.1.
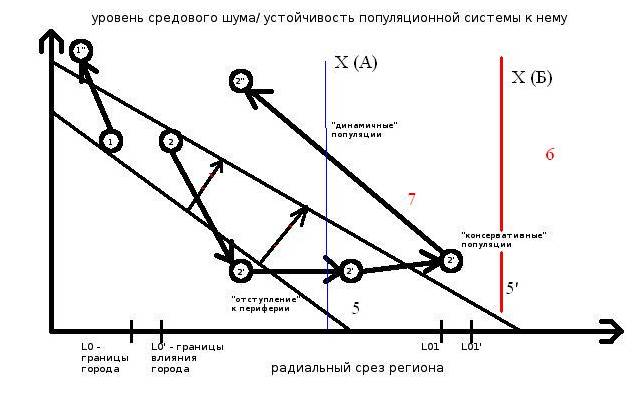
Рисунок 1. Прямая (1) и возвратная (2→2’→2“) урбанизации видов региональной авифауны (схема).
Обозначения. Ось Х – радиальный срез региона: L0/0’ – граница города в последовательные моменты урбанизации (5 и 5’, стрелки с красной полоской), L01/01’ – то же, его зоны влияния в регионе, 2’ – период «отступления» вида на периферию региона под «прессом» трансформации ландшафта, вызванной урбанизацией территории, 2“ – «поворот», вызванный лабилизацией жизненной стратегии вида (и типов отношений внутри популяции) после L лет «отступления».
4 — устойчивость и/или численность популяции редкого вида по завершению возвратной урбанизации. 5 и 5‘ — градиент региональной урбанизации в последовательные моменты времени, за которое её степень вырастет, а уязвимый вид – отступит к периферии. Этот «городской градиент» формирует закономерные изменения «архипелага» местообитаний и, соответственно, степени средового стресса (environmental stress) для особей на «островах» и неустойчивости — для метапопуляции вида.
6 — малонарушенная периферия региона, где существование вида устойчиво. 7. Момент превращения «консервативной» популяции в «динамичную», отличающуюся «проточной» структурой группировок и другими особенностями социальной и популяционной организации (Фридман, Еремкин, 2009, табл. 3). Х – оптимальный уровень средового стресса (ее проекция на ось Х – «точка старта» возвратной урбанизации) для обычных (А) и исчезающих (Б) видов.
Примечание. Жизнеспособность популяций 1’, 2’, 2” требует устойчивости их пространственно-этологической структуры большей, чем уровень средового стресса (оба параметра отложены по одной ос У).
Возвратная урбанизация редких видов — последний «шанс на спасение» в староосвоенных регионах
Возвратная урбанизация — это фактически хэппи энд «истории взаимоотношения» вида с урболандшафтом. Второй перестал подрывать его популяции, но напротив, поддерживает их жизнеспособность, часто лучше исходных местообитаний, вследствие ряда преимуществ, получаемых «дикими» видами при освоении городов. А первый «вышел из тупика», куда был «загнан» урбанизацией: ликвидировал зависимость своих поселений от местообитаний региона, под «давлением» города испытывающим неблагоприятную динамику, успешно «пересел» из деградирующих естественных местообитаний в их техногенные аналоги. «На счастье» дикой природы, последние порождаются развитием города непрерывно и примерно в том же количестве (по общей площади и размеру соответствующих «островов»), в каком деградируют первые, т.ч. общий баланс не меняется (Фридман, 2010, 2017).
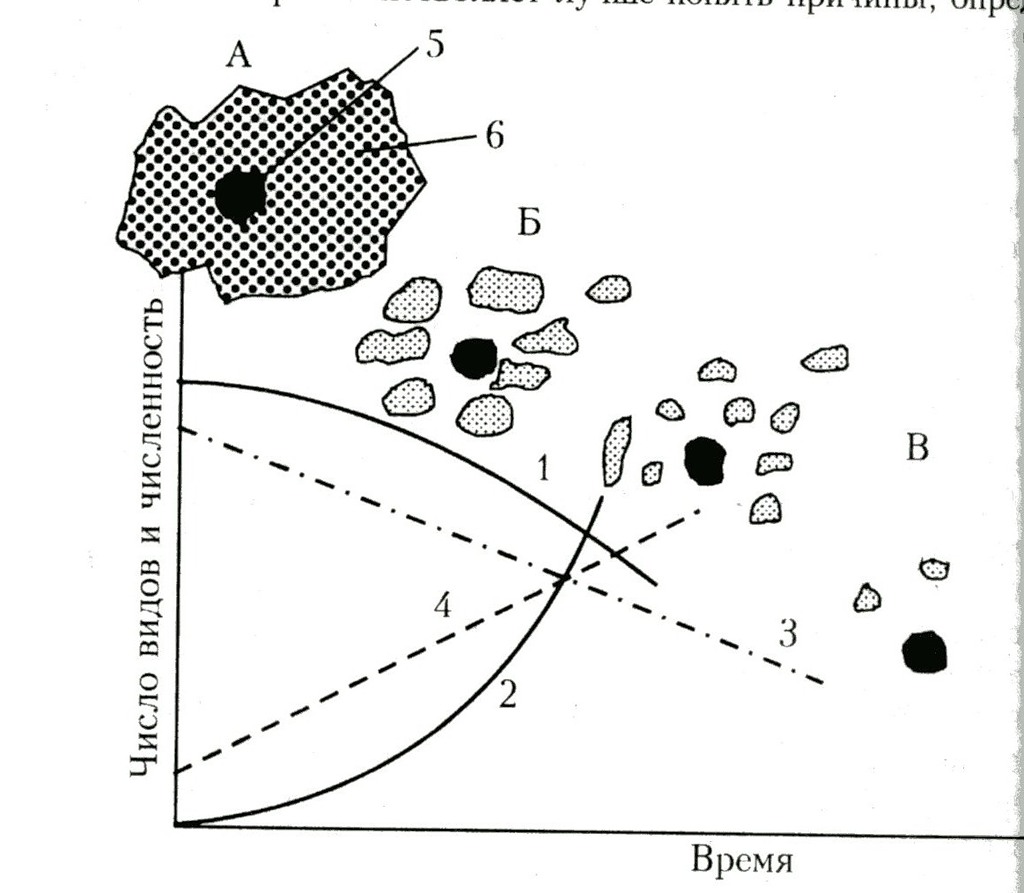
Пример неблагоприятной динамики «островного ландшафта» исходных местообитаний при урбанизации региона. Динамика биоразнообразия на ООПТ в связи с трансформацией природных ландшафтов вокруг: А – заповедник, окруженный коренными сообществами, Б – начало фрагментации окружающих территорий, В – усиление островного эффекта в связи с возрастающей фрагментацией окружающих заповедник территорий; 1 – видовое богатство аборигенной флоры, 2 – видовое богатство адвентивной и синантропной флоры, 3 – количество крупных хищников, 4 – межвидовая конкуренция крупных растительноядных, 5 – территория заповедника, 6 – окружающая природная территория. Из: См. Соколов В.Е., Филонов К.П., Нухимовская Ю.Д., Шадрина Г.Д, 1997. Экология заповедных территорий России. Ред. акад. РАН В.Е. Соколов, чл.-корр. РАН В.Н.Тихомиров. М.: Янус-К. 576 с. C.58.
Так, в Лондонской агломерации за 200 лет развития города было осушено 80 % водоёмов и болот, 40% площадей влажных лугов, и ровно настолько же приросла площадь техногенных болот и пустырей в составе разросшейся агломерации (Meritt, 1994).
Другой пример неблагоприятной динамики из: Хански, 2010
В Московском регионе площадь деградировавших околоводных местообитаний примерно соответствует площади вновь созданных рыбхозов и техногенных водоёмов. Другое дело, что вновь возникающие техногенные аналоги естественных местообитаний нужно надлежащим образом обустроить, чтобы дикие виды птиц (бабочек, тритонов, лягушек и т.п.) могли их заселить, тем самым облегчив «пересадку». Чтобы техногенные водоёмы, пустыри, рекреационные леса не оставались бэдлендами, нужно производить их экообустройство, способствующее максимальному а) заселению «дикими» видами фауны и флоры, б) «приближению» парцеллярной структуры сообщества к естественным экосистемам и в) содействующее урбанизации соответствующих видов (Авилова, 1997; Фридман, 2017).
Тем самым достигается двойная выгода (кроме очевидных природоохранных плюсов): формирование самоподдерживающейся экосистемы на месте техногенного водоёма, ещё вчера бывшего грязной лужей, отравляющей окрестности, позволяет бесплатно утилизировать отходы городского хозяйства, очищать стоки, которые иначе придётся очищать за деньги — плюс создаёт новый рекреационный ресурс.
Конечно, так будет лишь если вид за время «отступления» не вымрет вовсе. Обратите внимание: на рис.1 картина весьма динамична: кольцо II растет, III – «рвется» и сокращается наперегонки с отступлением вида к периферии региона. Чтобы возвратная урбанизация вообще началась, вид должен не вымереть за время L., поэтому крайне важно заблаговременное создание там ООПТ, позволяющих ему «продержаться» до начала процесса 3. возвратной урбанизации. Таким образом, смысл традиционной стратегии природоохраны в староосвоенных регионах – заранее подготовить «станции переживания» для максимального числа уязвимых видов, в достаточно крупных и малонарушенных массивах в кольцах II–III, и не отдельными пятнами, а соединенными в экосеть (Фридман и др., 2016).
Иными словами, территориальная охрана редких и уязвимых видов и «содействие» их урбанизации средствами биотехнии, планирования урболандшафта, специального восстановления природной мозаичности в рекреационных лесах, приближающей их к доагрикультурному «идеалу» и пр. (Коротков, 2017)
а) вовсе не исключают, но взаимно дополняют друг друга, «работая» на разных стадиях процесса рис.1, и лучше всего их использовать одновременно для разных задач, как щит и меч в рукопашном бою;
б) на «нижнем» полюсе градиента урбанизации (о нём см.: Фридман и др., 2016, рис.3, 4) первое будет ведущим, второе — ведомым, на «верхнем» наоборот, но оба они присутствуют, дополняя и усиливая друг друга, по всей «длине» градиента, нигде не обойтись чем-то одним, забыв про другое (Shafer, 2008; Marzluff, Ewing, 2008; Soulé, 2008; Yokohari, 2008).
Тем более в большинстве случаев заповедники, не говоря уж о прочих ООПТ, слишком малы и слишком преобразованы многовековым с/х использованием до заповедания, чтобы осуществлять все экологические функции на своей территории, эффективно саморегулироваться и пр. Увы, заповедание, как и использование, влекут за собой экологический риск в виде потери биоразнообразия, вспышек размножения филло- и ксилофагов, губящих древостой, других негативных процессов (Смирнова, Торопова, 2010; Торопова, Смирнова, 2014). Для пресечения их и снижения риска ООПТ на периферии требуют управления не меньше, чем «острова» дикой природы в городе, только немного иного.
В последние 30-40 лет возвратная урбанизация всё больше захватывает виды «с вершин» экологической пирамиды или высоко консервативные территориально, с жёсткими биотопическими предпочтениями (почему они угрожаемы и уязвимы в староосвоенных регионах). Их полагали неспособными неспособными обитать вне крупных массивов ненарушенных лесов, верховых болот и иных природных ландшафтов, и зря строили охрану именно и только на сохранении этих последних.
А это оказалось неверным, и именно «динамические популяции», изменившие прежние отношения со средой и «свернувшие» к возвратной урбанизации, растут численно, расширяют ареал, «выводя» вид из-под угрозы исчезновения. «Консервативные» популяции в традиционных местообитаниях, наоборот, не растут численно и неустойчивы даже при эффективной охране местообитаний. См. соответствующие «истории» для чёрного аиста Ciconia nigra (Грищенко, 1994), среднего Dendrocopos major и белоспинного D.leucotos дятлов, клинтуха Columba oenas (Бобенко и др., 2007; Гаврилюк, 2009; Белик и др., 2010), орла-могильника Aquila heliaca (Ветров, Милобог, 2008, 2010; Белик и др., 2012; Корепов, Стрюков, 2015; Павлов, 2015), лебедя-кликуна Cygnus cygnus (Boiko, Kampe-Persson, 2010), степной пустельги Falco naumanni (Ильюх, 2005), косматого поползня Sitta villosa (Назаренко, 2005), европейского тювика Accipiter brevipes (Белик и др., 2011; Федосов, 2015) и других видов данной категории (везде цит. по Фридман и др., 2016).
Обзор данных об использовании «города как заказника» глобально угрожаемыми видами птиц, млекопитающих, амфибий и рептилий показал, что 49 из них сохраняются в городских лесах, лесопарках и т.д. природных территориях городов существенно лучше чем вовне. В том числе в городах вне сходного ареала: какаду Cacatua sulphurea, исчезающий из-за торговли экзотическими птицами, сохраняется на островах в составе Гонконга. Сейчас там обитает 10% мировой популяции, причём «городское» население растёт, в природных же биотопах Индонезии и В. Тимора – падает. Эти благополучные популяции в городах могут быть и источником для реинтродукции и удовлетворять спрос на экзотов (Gibson, Li Yong, 2016).

Какаду Cacatua sulphurea, динамика «городского» и «природного» населения вида (следующий рисунок)
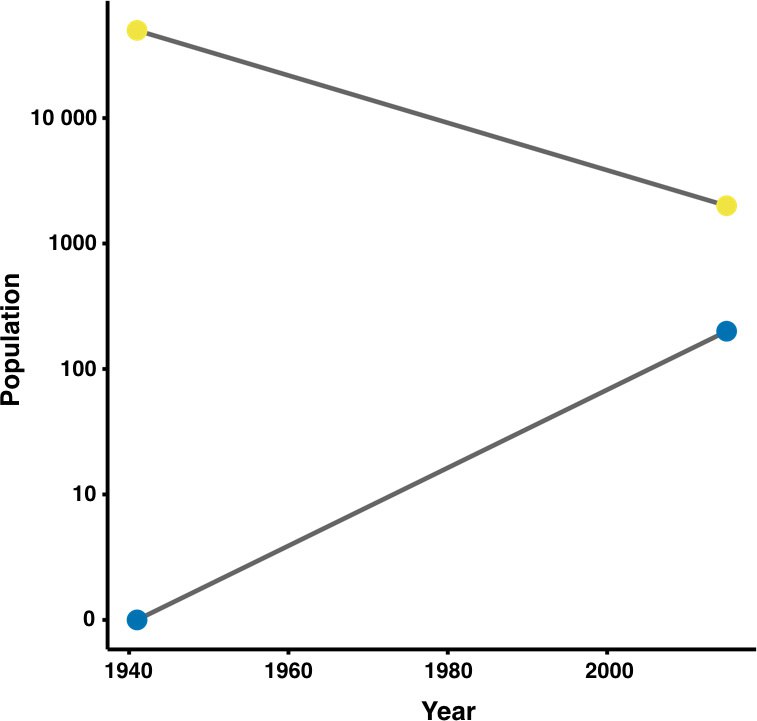
Понятно, что эта статья — попытка обратить нужду в добродетель, но важно не забывать о данной возможности. Другой момент: кроме позвоночных, среди «пользующихся» городом как «заказником» явно также растения и насекомые. Скажем, кизильник блестящий Cotoneaster lucidus из Красной книги РФ, активно используемый в озеленении городов.
Обобщение этих случаев и постоянное появление новых заставляет считать, что к возвратной урбанизации способны все виды региональной авифауны: урбофобия некоторых ситуативна или преходяща. Однако же «время задержки» (L рис.1) различно — чем консервативней жизненная стратегия вида в сравнении с родственными или экологически аналогичными, тем длительней «отступление» на периферию со спадом численности и сжатием ареала (Фридман, 2010, табл.1; Фридман и др., 2007, 2016). Дальше так или иначе следует «поворот» в противоположную сторону: «движение вверх» по градиенту урбанизации, переход к устойчивому существованию во всё более расчленённых, всё меньших по площади и непостоянных во времени местообитаниях, нежели на «материке» (рис.1).
При урбанизации, равно «прямой» и «возвратной», сначала осваиваются наиболее крупные «острова» природных территорий, «захваченные» по мере роста городов с соединением соседних в агломерации (и/или наиболее связанные с «материком» лесов, лугов и болот, оставшихся вне этих процессов региональной урбанизации), потом более мелкие, сильней изменяющиеся во времени, более изолированные застройкой/коммуникациями и т. д. В пределе эта тенденция завершается гнездованием в городской застройке и/или во вкраплённых в неё куртинах деревьев и кустов (Фридман, Ерёмкин, 2009; Фридман и др., 2016).
Т.е. «стартовые позиции» процесса представляют «архипелаг» рекреационных лесов, лугов, болот в зоне влияния городских ареалов (Dale et al., 2015, цит.по: Фридман и др., 2016). Последние же осваиваются от периферии к центру, по градиенту урбанизации «вверх» (рис.1), а не как-то иначе. Скажем, был бы возможен «альтернативный путь» от центральных районов, освоенных «наиболее городскими» птицами, выделившимися именно по этой причине из населения региона, к периферии, когда популяция разрастётся и пополнится «более лесными особями».

Однако пространственный вектор урбанизации соответствует первому варианту, а не второму. Так, он фиксировался на модельных видах – вяхире Columba palumbus, сороке Pica pica, чёрном дрозде Turdus merula, чибисе Vanellus vanellus, в Калининграде (Лыков, 2009), также как у других видов (сороке, ушастой сове Asio otus, среднем, белоспинном и малом пёстром D.minor дятлах и пр.) и в других городах: Берлине, Дрездене, Воронеже, Харькове и пр. (Фридман, Ерёмкин, 2009; Фридман и др., 2008, 2016). Что отражает рост устойчивости специализированной городской популяции к экстремально изменчивой во времени[9] и пространственно гетерогенной среде обитания, отличающей города от других типов «островных ландшафтов», созданных антропогенной фрагментацией — лесных масcивов, разделённых а/д сетью, лесными фрагментами среди с/хугодий, мозаикой частично осушенных торфяных болот и пр. (Pickett et al., 2008; Alberti et al., 2008; Фридман и др., 2016, рис.5).
Именно эти общие характеристики, а не «специфически городские» воздействия[10] ответственны за неспособность вида освоить урболандшафты данного региона. Действительно, в годы подъёма численности все виды региональной авифауны пробуют загнездиться в городе, обычно успешно — но не у всех возникшие поселения «удерживаются» надолго. Или, при многолетней сохранности отдельных группировок вид не может расширить городской ареал с первоначально занятых «островов» на ближайшие участки тех же местообитаний, остаётся зависимым от любых перестроек окрест (Фридман, Ерёмкин, 2009).
Иными словами, проблема не в негативных воздействиях города на живущих и размножающихся там особей, но в неустойчивости популяции на «архипелаге» местообитаний. Там она и так неустойчива вследствие «островного эффекта» (Хански, 2010; Акатов, 2012), плюс ещё в городе «острова», а равно разделяющий их матрикс максимально нестабильны во времени и гетерогенны пространственно, т. к. их размер, «кружево» размещения, взаимная изолированность и пр. постоянно меняются — для нас частью закономерно, для птиц полностью непредсказуемо.

Обретая устойчивость (т. е. адаптацию общего значения к новой экстремальной среде обитания), «городские» особи всех видов перестают быть метапопуляцией и складываются в новую популяцию — городскую. Одновременно их поселения теряют уязвимость от «островного эффекта»; видам больше не угрожает «отложенное вымирания» (extinction debt) даже на самых малых и изолированных «островах», бывшее главной угрозой доселе (Soulé, 2008; Акатов, 2012; Фридман и др., 2016).
Успешное завершение процесса (вид делается синурбанистом, хотя и необязательно синантропом, возникает специализированная городская популяция с динамикой и судьбой, независимой от «лесных») фиксируется при долговременно-устойчивом существовании популяционных группировок вида на «архипелаге» его городских местообитаний.
Формирование городской авифауны: баланс «пополнения» и «исчезновения»
Долговременный анализ «историй взаимодействия» разных видов птиц с растущим «ядром» Московской агломерации, организующим подчинённые города и города-спутники (Фридман, Ерёмкин, 2009, табл.2) показывает, что и жизнеспособные поселения могут сократить численность и исчезнуть вовсе вследствие постоянно идущих перестроек «архипелага» местообитаний в самом городе и в зоне его влияния.
На этом основывается прогноз динамики авифауны Большой Москвы, выполненный Г.С.Ерёмкиным в 2000 и 2010 гг., удовлетворительно оправдавшийся (Фридман, Ерёмкин, 2009, главы IV-V; Ерёмкин, 2010, 2014). За вычетом, увы, действий по «благоустройству», эффективно срывающих идущую урбанизацию разных видов. Так, благодаря «зелёным клиньям» советского периода Москва была «соловьиной столицей мира» (Марова и др., 2014). Проводимые СОПР учёты поющих самцов и исследования экологии восточного соловья Luscinia luscinia показывали успешную урбанизацию – если бы не «благоустройство» в последние годы[11] (Авилова, Кияткина, 2017).
Однако несмотря на неожиданные для нас и других обывателей покушения на местообитания «диких» видов в интересах стройкомплекса, последние в большинстве случаев остаются поблизости, в их трансформированных аналогах и пр. но не уходят из города. См. левый столбец табл.1; лишь меньшинство близких видов, в городе экологически идентичных оставшимся, исчезает, не пробуя закрепиться (правый).
Таблица 1.
Противоположное реагирование близких видов на локальное уничтожение гнездовых местообитаний (на примере авифауны г.Москвы).
| Вид, остающийся на гнездовании в городе,находящий замену утраченным местообитаниям | Вид, прекращающий гнездование в городе даже при наличии замены утраченным местообитаниям |
| Aythia fuligula | Aythia ferina |
| Podiceps cristatus | P.nigricollis |
| Sterna hirundo, Larus canus | L.ridibundus |
| Gallinula chlorophus | Fulica atra |
| Turdus pilaris | T.iliacus |
| T.merula | T.philomelos |
| Anas platyrhynchos | A.querquedula, A.clypeata |
| Charadrius dubius | Vanellus vanellus |
| Cyanosylvia svecica | Acanthis cannabina |
| Ficedula hypoleuca | F.parva |
| Motacilla citreola | M.flava |
| Dendrocopos minor | D.leucotos |
| Oenanthe oenanthe | Saxicola rubetra |
| Locustella fluviatilis | Locustella naevia |
| Erithacus rubecula | Troglodytes troglodytes |
| Sylvia atricapilla | Sylvia borin |
Источник. Фридман, Ерёмкин, 2009, табл.2.
Примечание. Данный перечень далек от завершения, а отдельные примеры требуют уточнения. Но не вызывает сомнения сам факт существования в городской авифауне «пар» экологически близких видов, противоположно реагирующих на антропогенную динамику местообитаний.
Фактически они остаются уязвимыми, даже заняв подходящие местообитания и успешно там размножаясь. Что происходит из-за неспособности гибко перераспределять поселения вида в городском ареале в ответ на постоянные трансформации «кружева» подходящих местообитаний, одни фрагменты которого ставятся под угрозу идущими перестройками города, другие появляются вновь, но тоже только на время и пр. Из-за «негибкой» реакции на направленные изменения урбосреды, сплавленные с флюктуациями, виды правой колонки табл.1 в противоположность экологически идентичным «левым» «заперты» на исходно освоенных «островах» и зависимы от судьбы этих локальных участков, рано или поздно оказывающихся под угрозой, т.к. постоянные перестройки охватывают весь городской ареал. Проблема здесь на уровне популяционной системы как целого, отдельные особи же успешно живут и размножаются, пока есть поселения — но они неустойчивы.
Какой вид ни возьми, в «кружеве» его городских биотопов одни «острова» исчезают, другие появляются, другие исчезают (для нас — в закономерной связи с эволюцией урболандшафта; для птиц — непредсказуемым образом), не говоря уж об изменениях размеров и связности их друг с другом. Успешно урбанизирующиеся виды не только устойчивы к подобным «прыжкам», но и используют их в «своих интересах». Анализ динамики фауны Калининграда (Лыков, 2009; Лыков, Гришанов, 2018), Москвы, Рязани и других городов показывает, что они направленно перемещают популяционные группировки от исчезающих «островов» к вновь появляющимся, и по мере успешности этих «прыжков» с «льдины» на «льдину» ещё больше урбанизируются, т.е. осваивают всё меньшие, нестабильные и более изолированные острова.
Данная положительная обратная связь доводит начавшуюся урбанизацию до максимума, возможного в данном городском ареале с его размером, мозаичностью и скоростью перестроек в разных масштабах пространства.
Отсюда устойчивость существования популяционных систем в местообитаниях города больше зависит не от их «качества», или локальных воздействий от дорог и застройки на соответствующие «острова» (Клауснитцер, 1990, рис.49), но от «конструктивных характеристик» самой системы, обеспечивающей её устойчивость «вопреки» экстремальному уровню нестабильности и гетерогенности урбосреды. Соответственно, судьба разных категорий видов в составе авифауны города зависит при прочих равных не столько от различий в морфоэкологической специализации или биотопических предпочтениях, сколько от степени «продвижения» в сторону урбанизации. См. табл.2 для авифауны г.Москвы.

Малый пёстрый дятел Dendrocopos minor. В Англии и большинстве стран ЕС неустойчив, в центре европейской России успешно урбанизируется, как и белоспинный дятел Dendrocopos leucotos (на следующей картинке). Фото В.В.Солодушкина.
Таблица 2
Устойчивость существования разных видов в Московской агломерации в зависимости от консерватизма-лабильности жизненной стратегии
| Видовая стратегия | Темпы урбанизации | Прогноз сохранения вида в урболандшафте | Число видов из авифауны Подмосковья |
| А. Реагирующие на урбанистические изменения пригородных ландшафтов | формируют специализированные городские популяции при успешном завершении урбанизации вида, после чего осваивают урболандшафты самостоятельно. | ||
| 1. Лабильная | Урбанизация немедленная, быстрая, необратимая | Прогноз благоприятный, образованные городские популяции устойчивы при спонтанной эволюции урболандшафта | 10 – синантропы 23 — синурбанисты |
| 2. Лабилизиру-ющаяся | Урбанизация возвратная – после оттеснения и 20…40 лет пребывания в оттеснённом состоянии | Прогноз умеренно благоприятный, существует риск сокращения и вытеснения вида при последующей эволюции урболандшафта, который снимается экообустройством конкретных «островов» местообитаний | 24 |
| 3. Консервативная | Урбанизация возвратная – после оттеснения и 40…80 лет пребывания в оттеснённом состоянии | Прогноз неблагоприятный, риск сокращения численности и вытеснения уже поселившихся видов высок, снимается созданием устойчивого экологического каркаса всего города | 7 – уязвимые синантропы 12 – виды остатков лесов и пашни. Или занявшие техногенные аналоги исходных биотопов, но не выходящие за их пределы (правый столбец табл.1). |
| Б. Не реагирующие на урбанистические изменения пригородных ландшафтов | |||
| `1. «Пассивные урбанисты» | - | Гнездятся пока существуют их местообитания. Но если в городе они разрушаются и/или пресечён «коридор», связывающий занятые участки с основными массивами местообитаний вовне, то снижают численность и исчезают пропорционально обоим процессам, не делая каких-либо «попыток остаться». | 32 |
Источники. Фридман и др., 2008, 2016; Фридман, Ерёмкин, 2009.
Поэтому авифауна города складывается как баланс «пополнения» устойчивыми видовыми популяциями, за счёт развивающейся урбанизации, и «исчезновения» видов, бывших и особенно ставших неустойчивыми в связи с преобразованием биотопов[12].
Второе случается независимо от степени синантропности вида =приспособленности особей к зданиям и сооружениям, толпам людей, дорогам, другим составляющим урболандшафта, давнее и успешное использование всего этого при поиске корма, ночёвках, размножении и пр. Всё это отнюдь не снижает риск подрыва устойчивости популяций таких видов на следующем «шаге» развития городских ареалов (Фридман и др., 2016).
 Действительно, многие традиционные синантропы — городские ласточки Delichon urbica, сипухи Tyto alba, обыкновенные пустельги Falco tinnunculus, даже домовые воробьи Passer domesticus с галками Corvus monedula в нынешних мегаполисах сокращают численность и исчезают. В противоположность синурбанистам, успешно освоившим природные территории в тех же ландшафтах, и не идущим в застройку. Следовательно, термин «синантроп» указывает на адаптированность особей к человеку и созданной им «второй природе» на уровне особей, тогда как «синурбанист» — на
Действительно, многие традиционные синантропы — городские ласточки Delichon urbica, сипухи Tyto alba, обыкновенные пустельги Falco tinnunculus, даже домовые воробьи Passer domesticus с галками Corvus monedula в нынешних мегаполисах сокращают численность и исчезают. В противоположность синурбанистам, успешно освоившим природные территории в тех же ландшафтах, и не идущим в застройку. Следовательно, термин «синантроп» указывает на адаптированность особей к человеку и созданной им «второй природе» на уровне особей, тогда как «синурбанист» — на
1) устойчивость видовых популяций,
2) достигаемой вопреки резкой пространственной гетерогенности занятого ландшафта и быстрым изменениям его мозаик во времени. Важно, что 1)-2) реализуется независимо от того, что именно занимается видом: «пятна» природных сообществ или участки застройки, и к каким изменениям устойчивы его группировки — направленной эволюции планировочной структуры всего города, или флюктуационных перестроек на уровне отдельных домов и кварталов в заселённых местностях и/или вокруг (Клауснитцер, 1990:.
Если развитие города реализует потенции «заказника»[13], виды «сами справляются» с освоением «островного ландшафта» вокруг и внутри городских ареалов, первое превалирует над вторым, равновесие сдвигается в сторону большего уровня биоразнобразия. Исходя из универсальности этих процессов, и протекания первого даже в городах с, мягко говоря, не лучшей экополитикой, мы предложили корректировку классических моделей «островной биогеографии» (Фридман, 2012в).
Фауна каждого из «островов», как и города в целом, больше предсказываемой ими равновесной (заданной балансом реколонизации и вымирания с учётом наличия или отсутствия «коридоров», соединяющих «остров» с «материком») на число видов, претерпевающих возвратную урбанизацию во всём регионе (Фридман и др., 2016). Поскольку консервативные и стенотопные виды чем дальше, тем больше вовлекаются в этот процесс, данное расхождение при прочих равных будет расти, при пропорциональном падении риска отложенного вымирания. И в идеале город «вместит» (обеспечит устойчивость существования) всей региональной авифауны – если сохранён и/или восстановлен экообустройством «техногенных аналогов» поляризованный ландшафт, обеспечивающий не меньшую связность природных участков, чем фиксируется у городских ареалов при агломерировании (Родоман, 2002).
Плюсы урбанизации для «диких» видов птиц
Факторы привлекательности рассматриваются для «урбанизированного ядра» региона вместе с зоной влияния города, где природные ландшафты сильно фрагментрованы извне и инсуляризированы изнутри, и долговременная динамика эти «осколков» естественных экосистем определяется развитием «ядра» и подчинённых ему городских центров. Граница современного города определяется изохронной часовой доступности, граница зоны влияния города – средним радиусом поездок горожан «на природу» на уик-энд (Лаппо, 1997, здесь и далее цит. по Фридман, 2010; Фридман, Ерёмкин, 2009). На аэрофотоснимках граница зоны влияния определяется по переходу между
а) чёткими и угловатыми границами «многоугольников» природных сообществ, разделённых дорогами, дачными участками и другими антропогенными участками/коммуникациями, к
б) плавным и извилистым границам между «пятнами» лесов, лугов, болот и т.п., когда последние примыкают естественным образом, а не разрезаны развитием человеческой инфраструктуры.
Всего просматриваются семь факторов привлекательности.
- Развитие городской экономики и территориальный рост городов формируют интенсивный подток в городской ареал органического вещества и энергии извне, с обширной территории «экологического следа» города (ecological footprint), размер которого намного превосходит тот регион, с которого городское развитие получает рабочую силу (Rees, Wackernagel, 2008). Эти дополнительные ресурсы вещества и энергии используются урбанизирующимися «дикими» видами птиц и снимают проблему конкурентного исключения «пришлых» видов, постоянно присутствующего в природных сообществах.
- Более мягкий климат городских центров (более морской по сравнению с континентальным климатом сельской периферии) обязан существованием «острову тепла» (Alcoforado, Andrade, 2008; Kuttler, 2008). Он ответственен за более тёплые-влажные зимы и ранние вёсны в городах сравнительно с округой, что позволяет раньше начать размножение и успешней его провести, в т.ч. вследствие сокращения вызванной холодом смертности взрослых зимой, кладок и птенцов – возвращением холодов после (Куранов, 2009; Фридман, Ерёмкин, 2009). Освещение негативно воздействует на «негородских» особей, в т.ч. в период миграций (McLaren et al., 2018). В городе же оно скорей позитивно: позволяет эффективно кормиться за пределами светлого времени суток, и размножаться – за границами обычного времени репродукции (Клауснитцер, 1990).
- Умеренное нарушение (и именно того типа, что производит рост «городского ядра» в «своём» регионе) увеличивает продуктивность природных ландшафтов. Фрагментация лесных массивов увеличивает запасы беспозвоночных в возникающих лесных фрагментах по сравнению с единицей площади исходного «материка», умеренное рекреационное нарушение лесных и луговых биотопов увеличивает их продуктивность в отношении зелёного и семенного корма для грызунов и так далее (Esa et al., 1994; Сарычев, 2001, Жигарев, 2002, 2004).
- Виды, освоившие урбосреду, освобождаются от лимитирования ресурсами (п. 1). С позитивным влиянием умеренных антропогенных нарушений в плане обеспеченности кормовой базой потенциальных вселенцев (особенно значимо здесь фрагментации природных ландшафтов извне и инсуляризации изнутри, п. 3) плюсуется увеличение продуктивности городских местообитаний из-за интенсивного подтока в город органического вещества и энергии с территории «экологического следа» города.
- Успешное освоение городской среды выводит вид из-под пресса хищников и/или более специализированных конкурентов. Последние, будучи крупнее и, как правило, обладая большим консерватизмом территориальных связей, в подавляющем большинстве случаев урбанизируются позднее.
- Успешная урбанизация вида позволяет ему увеличивать численность в «архипелаге» городских местообитаний, «не чувствуя» ограничений, связанных с островным эффектом и с недостаточной площадью подходящих «островов», Исчезает положительная связь площади «острова» и плотности/общей численности», достигаемой данным видом на нём. Эта же связь фиксируется для вероятности регулярного гнездования на данном «острове» разных видов из состава фауны «архипелага»; она тоже исчезает при урбанизации данных видов. Что верно для отдельной видовой популяции, верно и в отношении орнитокомплексов: последние на урбанизированных территориях не «ограничены сверху» по числу видов, способных к совместному обитанию на каждом отдельном «острове», если только соответствующие виды «умеют» поддерживать устойчивость популяций на «архипелаге» в целом, охватывающем всю территорию города. Соответственно, они всегда могут быть «пополнены» новыми видами из состава региональной фауны, как только их популяции станут устойчивы в урболандшафте.
- Из-за отсутствия лимитирования ресурсами и биоценотическими отношениями вроде хищничества и конкуренции, урбанизация выводит «дикие» виды из биоценотических ограничений. «Городской воздух делает свободными» урбанизированные популяции разных видов птиц и млекопитающих в том же смысле, в каком он делал свободным средневекового крестьянина. Соответственно, орнитокомплексы города, как на природных, так и на застроенных территориях всегда «открыты» для включения новых видов, если только те в достаточной степени урбанизированы и могут образовать популяцию, жизнеспособную и устойчиво воспроизводящуюся на данном сегменте городской территории. Напротив, фаунистические комплексы природных территорий вне города «закрыты» в том смысле, что включение новых видов возможно лишь по более богатым местообитаниям, например, поймам рек, или через разрушение коренных сообществ видами-интродуцентами, столь часто описываемое для островов (Шварц, 2004).
Например, в 6 типах местообитаний Валдайской возвышенности Е.А.Шварцем и др. (1992) обнаружена строго положительная корреляция между относительной биомассой разных видов землероек и беспозвоночных (P<0,008). Годовое потребление этого корма землеройками составляет от 32 до 151% летних запасов почвенных и напочвенных беспозвоночных.
Если же учесть суммарное изъятие этого корма другими группами потребителей (бесхвостые амфибии, птицы, ящерицы, грызуны, хищные беспозвоночные), то общее потребление беспозвоночных всеми группами консументов в каждом типе естественных местообитаний окажется у предела возможности нормального воспроизводства популяций жертв. Зимнее сокращение почвенной мезофауны вызывает сложности с поддержанием численности насекомоядных в весенне-летний период (Шварц и др., 1992).
Тот же результат получен относительно воздействия всего сообщества птиц на потребляемые группы беспозвоночных – листогрызущих, ксилофагов и пр. (Иноземцев, 1978). Норма для природных сообществ (включая антропогенно нарушенные) — отсутствие “лишних” ресурсов вещества, энергии и безопасности (плотность “убежищ”, их доступность в случае “штатной” опасности), “не освоенных” популяцией какого-либо вида из “постоянного состава” фауны (Алещенко, Букварёва, 1997). Что резко ограничивает включение новых видов в этот последний.
После «захвата» территории в структуру урболандшафта или хоть в зону влияния города этот ресурсный «предел роста» численности новых видов исчезает тем больше, чем полней первое. Действительно, осваивающие его виды птиц и млекопитающих, исходно экологически близкие, в отличие от природных сообществ отнюдь не делят ниши, но напротив, сближают их до неразличимости, см.далее. «Неизрасходованный» «постоянным составом» биоты избыток органики и энергии облегчает проникновение чужеродных видов. Часть из них на полностью изменённых терри- и акваториях оказывается позитивной: восстанавливает их экологические функции в рамках всего ландшафта (Фридман, 2017; Johnston, Munshi-South, 2017). Благодаря этому соответствующе группировки видов постепенно приближаются к сообществам (при невозможности полного приближения, т.к. все городские местообтания, в т.ч. максимально «прродоподобные», формируются больше антропогенными «возмущениями», нежели средопреобразованием своих видов).
Соответственно, орнитокомплексы города, как на природных, так и на застроенных территориях всегда «открыты» для включения новых видов, если только те в достаточной степени урбанизированы и могут образовать популяцию, жизнеспособную и устойчиво воспроизводящуюся на данном сегменте городской территории. Напротив, фаунистические комплексы природных территорий вне города «закрыты» в том смысле, что включение новых видов возможно лишь по более богатым местообитаниям, например, поймам рек, или через разрушение коренных сообществ видами-интродуцентами, столь часто описываемое для островов (Шварц, 2004).
Парадоксы городских экосистем, реализующие «преимущества»
С одной стороны (1), городские местообитания снимают ресурсные ограничения на число видов, способных одновременно жить на одной территории, городские местообитания содержат заведомый избыток ресурсов. Однако в городах увеличивается число случаев вытеснения одного вида другим, то есть конкуренция вроде бы обостряется.
Далее (2), в городских местообитаниях конкуренция ослабляется именно там и тогда, где вроде бы должна обостряться. В самых разных ситуациях мы видим увеличение перекрывания экологических ниш близких видов птиц именно при ухудшении кормовых и погодных условий, вплоть до полной экологической тождественности.
Так, в осенне-зимний период всякое снижение доступности корма в городских и рекреационных лесах увеличивает перекрывание топических и временных «ниш» у видов-членов синичьих стай (разные виды синиц, пищухи, поползни и пр.). «Ниши» очерчены на основании вероятности совместных встреч разных видов в определённых микростациях, регулярно посещаемых стаей при обследовании собственной территории (Berner, Grubb, 1985; Иноземцев, 1987). Расширение видовой ниши с большим перекрыванием ниш разных видов-членов синичьих стай в ответ на неожиданное снижение доступности корма и устойчивое ухудшение условий его добывания постоянно наблюдается
а) в межгодовых сравнениях,
б) при сравнении участков обитания разных стай, различающихся по данному показателю;
в) при сравнении разных периодов существования одной и той же стаи осенью и зимой (более и менее благоприятных), Rolando, Robotti, 1985.
Следовательно, эта зависимость устойчиво воспроизводится в широком диапазоне внешних условий и может считаться закономерностью организации многовидовых сообществ в городе. Упрощение ярусно-оконной структуры растительности в процессе урбанизации (которую обследует многовидовая стая в поисках корма) её усиливает и делает более выраженной (Rolando, Robotti, 1985, сравни с Иноземцев, 1987; Боголюбов, Преображенская, 1987).
Действительно, деление городских сизарей на три альтернативные стратегии становится гораздо менее выраженным при общем падении численности, вызванном интенсивным строительством современных зданий в центре города, распадом крупных колоний, появлением вместо них нестабильных мелких колоний (г.Пермь, Ангальт, 1988). Выраженность альтернативных стратегий в поведении индивидов в годы спада настолько уменьшилась, что само определение стратегий по данным о регулярностях поведения особей стало затруднительным.
Последнее доказывает, что выделение альтернативных стратегий и связанные с ним перестройки пространственно-этологической структуры городских поселений вида — следствие изменений на уровне популяции как системного целого, но не приспособления и отбора на уровне особей. Иными словами, в процессе освоения городской среды индукция приспособительных изменений на уровне особей (и иных уклонений от адаптивной нормы вида) происходит только в устойчивых и жизнеспособных популяциях, особенно экспоненциально растущих. А для этого популяционная система должна перестраиваться в сторону более «проточной» организации и лабильной стратегии особей (Фридман, Ерёмкин, 2009, табл.3).
Далее (3), согласно существующей экологической теории обострение конкуренции ведёт вроде бы к разделению ниш? но именно в городских биотопах ниши близких видов конвергируют до полного безразличия. То же самое наблюдается при выпадении снега, резком похолодании, затяжных дождях и других ухудшениях погодных условий, увеличивающих «напряжение жизни» у птиц (например, снижением доступности корма).
Эта конвергенция обнаружена у «городских» видов мышевидных грызунов даже в случае столь «мягкого» и постепенного воздействия, как рекреационная нагрузка на городские леса и парки.
На участках леса, не подверженных рекреации, экологические ниши (и характерные местообитания) близких видов в значительной степени разделены. Например, малые лесные мыши (Sylvaemus uralensis) приурочены к «неморальным» сообществам, рыжие полёвки – к «сухим» и «болотистым» растительным сообществам, имеющим «таёжный облик».
На участках рекреационного использования индивидуальные участки лесных мышей полностью (100%) пересекались с участками рыжих полёвок. Под воздействием рекреации также пропадает избирательность биотопических предпочтений разных видов: все устойчивые виды становятся эвритопными и биотопически пластичными. Например, у лесной мыши не обнаружено предпочтения каких-либо участков растительных ассоциаций и иных неоднородностей мозаичной среды (почвенного, эдафического характера), явно избегались лишь участки ксерофильного сосняка.
В годы депрессии численности виды-доминанты в естественных мозаичных биотопах — лесная мышь и рыжая полёвка — оптимально делят территорию, предпочитая разные микроусловия и разные типы растительности. На участках, нарушенных рекреацией, микробиотопические ниши обоих видов начинают полностью перекрываться при исчезновении сколько-нибудь видоспецифических предпочтений микростаций (Жигарев, Шаталова, 1985).
Сильная конвергенция местообитаний устойчивых видов также связана с концентрацией зверьков в общих «островках безопасности», на участках с наибольшей доступностью пищевых ресурсов (семян и зелёной массы). Это увеличивает степень перекрывания ниш разных видов. При умеренной рекреационной нагрузке исходно экологически различные виды оказывают в одних и тех же частично трансформированных местообитаниях с наилучшими кормовыми и защитными свойствами. Под воздействием рекреации увеличивается перекрывание даже оптимумов экологических ниш, но и их центров, вплоть до статистической идентичности последних (Жигарев, 1993).
Например, для малых лесных мышей «сухие» и «болотистые» сообщества с «таёжным» обликом – это в первую очередь «стации расселения», неблагоприятные для постоянного обитания. Но под воздействием умеренной рекреации позиции вида в первую очередь увеличиваются именно в этих сообществах, особенно сильно в сосняках-черничниках. Локальная плотность вида здесь максимальна (до 60 особей/га), средняя осенняя плотность в 3-20 раз превышает значения для участков леса, не подверженных рекреации, а интенсивность размножения наиболее высока. Сходным образом под воздействием рекреации для рыжей полёвки возрастает значение сосняка черничного. Тамошняя плотность населения – 101,8 особей/га – в «природных» местообитаниях фиксируется лишь в широколиственных или в островных лесах (Жигарев, 2002).
Далее, по мере нарушения растительности рекреационным воздействием у фоновых видов грызунов происходит рост эвритопности. Виды всё меньше дифференцируются в отношении предпочитаемых парцелл растительности, падает избирательность предпочтения каждым видом «своей» серии растительных сообществ (Жигарев, 2002). Процесс конвергенции ниш получает тем большее распространение, чем больше рост рекреационной нагрузки вызывает олуговение травостоя, а затем и замену лесных и луговых видов сорной и рудеральной растительностью. Тем более что сорная растительность в отличие от лесного и лугового разнотравья не зависит от градиентов гидротермического режима почвы, протягивающихся от положительных элементов рельефа к отрицательным (Коломыц и др., 2000).
Но увеличение эвритопности, расширение «городской» ниши происходит лишь у тех видов, популяции которых остаются устойчивы при данной интенсивности воздействия. У неустойчивых видов (тёмная полёвка Microtus agrestis) происходит обратный процесс — сокращение ниши с вытеснением популяций в наименее нарушенные, но пессимальные и нехарактерные местообитания. В своих естественных местообитаниях M.agrestis связана с влажными фитоассоциациями. В рекреационных лесах вид сохраняет стенотопность и не расширяет ниши, но занимает сухие участки нарушенного сосняка-беломошника и мезофильные – ольшанника-звездчаткового, противоположные по градиенту влажности (Жигарев, 2002).
M.agrestis также первой выпадает из фауны грызунов рекреационных лесов, причём вероятность вымирания вида на определённой территории резко возрастает после сдвига реализованной ниши в нетипичные местообитания. Важно подчеркнуть, что сдвиг ниши, увеличивающий уязвимость вида к рекреационным воздействиям, вызван именно неустойчивостью популяционной системы к средовому стрессу в условиях накопления нарушений растительного покрова при дополнительном воздействии фактора беспокойства, связанного с нарушением. Оно проявляется в первую очередь при пересечении тропинок, что связано со стрессом открытого поля, риском нападения хищника при переходе пространства, свободного от растительности.
По мере развития дорожно-тропиночной сети в рекреационных лесах неустойчивые виды грызунов всё больше блокируются внутри относительно ненарушенных «островов» растительности, что парализует нормальное кормление зверьков. Эта неустойчивость в условиях повышенного риска, — непосредственная причина вытеснения видов вроде тёмной полёвки, а не конкуренция и не рекреационное воздействие как таковое (Жигарев, 2002).
Следовательно, ни неблагоприятный сдвиг ниш стенотопных уязвимых видов, ни расширение ниш эвритопных устойчивых видов сами по себе не связаны с конкурентными отношениями между ними. Во всяком случае, первое под воздействием рекреации происходит существенно раньше, чем второе, конкуренция же требует продолжительного взаимодействия популяций. К тому же умеренное нарушение растительности под воздействием рекреации создаёт безусловный избыток зелёного и семенного корма для грызунов (как по качеству, так и по биомассе). Поэтому расширение топических ниш у одних и сужение у других видов обусловлено разным уровнем устойчивости соответствующих популяционных систем к разнообразным стрессорам, производным от всех трёх составляющих изменений — нарушения растительности, увеличения стохастической, непредсказуемой мозаичности среды, и процесса инсуляризации массива изнутри.
Тогда приспособительные возможности отдельных особей в городской популяции (выраженные «нетипичными» способами гнездования, сбора корма, защиты от хищников и пр.) есть функция устойчивости-неустойчивости популяционной системы вида в соответствующем городском ареале.
При устойчивости видовой популяции приспособительные возможности растут, увеличивается частота и разнообразие «уклонений» городских птиц от обычного видоспецифического поведения (что ошибочно трактуется натуралистами как «адаптация к условиям города). При потере устойчивости и падении численности популяции приспособительные возможности особей сокращаются до жёсткого следования видовым стереотипам. Это ведёт к ещё большей неустойчивости популяции в урбосреде. Например, в рекреационных лесах в паре малая лесная мышь – рыжая полёвка именно первый вид сильней меняет пространственное распределение при меньшей степени расширении ниши. Он же демонстрирует наибольшую колеблемость численности (Жигарев, 1997).
Если этот направленный сдвиг биотопических предпочтений был бы вызван «проигрышем» одного из видов в конкуренции, он сопровождался бы увеличением смертности (и других показателей увеличения скорости убыли популяции при сокращении темпов воспроизводства). Но в городской популяции интенсифицируются оба процесса: увеличивается гибель самих особей, их гнёзд и птенцов в разных «экологических ловушках» и интенсифицируется размножение в оптимальных местообитаниях. Подобная «двойственность» отмечена не только у млекопитающих, но и у птиц.
Парадоксы (1)-(3) позволяют уйти от ряда ограничений, затрудняющих охрану видов и сообществ в природных ландшафтах, в первую очередь зависимостей «виды-площадь»: более эффективная охрана биоразнообразия требует больших площадей ООПТ и большей связности. Дискуссии по проблеме SLOSS (some large or severall smаll — для размеров ООПТ, https://en.wikipedia.org/wiki/SLOSS_debate ) и негативные последствия для охраны природы там, где защитникам бизнеса или «экономического развития» удалось пробить второй вариант показывают, что вне города сохранение биоразнообразия строго пропорционально площади местообитаний, в т.ч. и особенно сохраняемых сетью ООПТ (Хански, 2010: 173-174).
При уменьшении последней «острова» местообитаний в ближних пригородах теряют виды ещё сильней, чем биотопически идентичные части «материка» (в чём и состоит, вообще говоря, «островной эффект», Акатов, 2012). Урбанизация видов ослабляет эти зависимости, как описано выше; кроме того, по ходу процесса у всех видов птиц фиксируется поведенческий «островной эффект» (ethological island effect) — ослабление территориальности, толерантность к повышенной плотности, усложнение социальных систем вроде пребывания части особей на чужих территориях в статусе неразмножающихся «помощников», как у чёрной вороны Corvus corone вместо самостоятельного поиска незанятых участков. Всё это ослабляет нужду в большей площади местообитаний проплорционально продвижению процесса урбанизации, хотя отнюдь не снимает её полностью.
Литература
Авилова Ксения, Кияткина Надежда. Птицы в городе// Наука и жизнь. 2017. №11. http://elementy.ru/nauchno-populyarnaya_biblioteka/433823/Ptitsy_v_gorode
Акатов В.В. 60 лет теории динамического равновесия островной биогеографии: проблемы тестирования, результаты полевых исследований, прикладное значение // Журн. общ. биол. 2012. Т.73. №3. С.163–182.
Быченко Т. М. Особенности биологии некоторых видов орхидных Южного Прибайкалья в связи с вопросами их охраны. Дис. … канд. биол. наук. М., 1992. 355 с.
Волкова Л.Б. Урботолерантные виды дневных бабочек и условия их сохранения в районах городской застройки// Животные в городе. Мат. I научн.-практ.конф. М.: РСХА, 200. С.71-74.
Волкова Л.Б., Бейко В.Б.. Проблемы сохранения разнообразия насекомых в Москве// Природа Москвы. М., Биоинформсервис, 1998. С. 239-250
Волкова Л.Б., Соболев Н.А. Разнотравный газон в современной концепции озеленения городов (на примере г.Москвы)// Лесной вестник. 2015. №5. С.145-152.
Вучик В.Р. Транспорт в городах, удобных доля жизни. М.: Территория будущего, 2011. 576 с.
Ерёмкин Г.С. О возможности прогноза состояния гнездовых популяций птиц города Москвы и ближнего Подмосковья// Охрана живой природы и природного комплекса Москвы. Мат-лы научн.-практ.совещ., посвящённого 100-летию со дня рождения К.Н. Благосклонова, 2010. С.38-40.
Карпачевский М.Л., Ярошенко А.Ю., Зенкевич Ю.Э. Природа Подмосковья: утраты последних двух десятилетий. М. : Изд-во ЦОДП, 2009. 92 с.
Коротков В.Н. Основные концепции и методы восстановления природных лесов Восточной Европы// Russian J. Ecosyst. Ecol. 2017. V.2. №1. http://rjee.ru/rjee-2-1-2017-1/
Крамер Д. А., Неруда М., Тихонова И.О.. Европейский опыт ревитализации малых рек// Биология. Экология. Естествознание. Науки о земле . 2012. №2. С.112-127.
Лыков Е.Л. Фауна, население и экология гнездящихся птиц городов Центральной Европы (на примере Калининграда). Автореф. и дисс.канд.биол.наук. М., 2009. 288 с.
Лыков Е.Л., Гришанов Г.В. Атлас гнездящихся птиц Калининграда// Калининград: Смартбукс, 2018. 320 с.
Марова И.В., Антипов В.А., Иваницкий В.В. Соловей в Москве: вокальные характеристики, современное состояние и перспективы выживания уникальной популяции//Редкие виды птиц Нечернозёмного Центра России. Мат. V. Совещания. М.: МПГУ, 2014. С.205-210.
Никифоров А.В. Рождение пригородной Aмерики: социальные последствия и общественное восприятие процессов субурбанизации в США (конец 40-х — 50-е гг. ХХ в.). М.: URSS, 2002. 356 с.
Разран Л. Восстановление видового разнообразия растений на сырых пойменных лугах в долине реки Айдер (Северная Германия) //Мастер-класс для Пантоподы. М.: Т-во научн.изд. KMK, 2007. С.195-197.
Родоман Б.Б. Поляризованная биосфера. Сборник статей. Смоленск: Ойкумена, 2002. 335 с.
Смирнова О.В., Торопова Н.А. Теоретические основы единой стратегии охраны природы и природопользования// Расширенный текст доклада на семинаре «Актуальные задачи Биологии охраны природы на Северо-Западе Российской Федерации», г. Вологда, 2 декабря 2010 г. http://www.socialcompas.com/2015/01/01/teoreticheskie-osnovy-edinoj-strategii-ohrany-prirody-i-prirodopol-zovaniya/
Соболев Н.А., Волкова Л.Б. Динамика состояния беспозвоночных, занесённых в Красную книгу города Москвы// Охрана живой природы и природного комплекса Москвы. Мат-лы научн.-практ.совещ., посвящённого 100-летию со дня рождения К.Н. Благосклонова, 2010. С.176-180.
Стольберг Ф.В. (ред.). Экология города. Учебник. Киев : Либра, 2000. 464 с.
Торопова Н.А., Смирнова О.В. Экологический риск заповедания и пути его преодоления: обзор современных представлений// Вестник ТГУ. 2014. Т.19. №5. С.1577-1580.
Фридман В.С., Ерёмкин Г.С., 2009. Урбанизация «диких» видов птиц в контексте эволюции урболандшафта. М.: УРСС. 235 с.
Фридман В.С. Город как рефугиум, и прогноз возвратной урбанизации «диких» видов птиц // Охрана живой природы и природного комплекса Москвы. Мат-лы научн.-практ.совещ., посвящённого 100-летию со дня рождения К.Н. Благосклонова, 2010. C.114-120.
Фридман В.С. Зачем нужна дикая природа в городе?// Скепсис. Ноябрь 2012а. http://scepsis.net/library/id_3354.html
Фридман В.С. Трудности биологической концепции вида и пути их преодоления (на примере птиц// Беркут. 2012б. Т.21. №1-2. С.127-182.
Фридман В.С. Урбанизация «диких» видов птиц и модели островной биогеографии // Биогеография: методология, региональный и методологический аспекты: Материалы конф., приуроченной к 80-летию со дня рождения В.Н. Тихомирова (1932–1997) (Москва, 30 января – 3 февраля 2012 г.), М., 2012в. С. 236-240
Фридман В.С., Королькова Е.О. Новое покушение на Воробьёвы горы// Инициативная группа студентов, аспирантов и сотрудников МГУ. 18.07.2013. http://igmsu.org/?itemid=875
Фридман В.С., Ерёмкин Г.С., Захарова Н.Ю., 2007. Становление популяционной устойчивости редких видов птиц в «островном ландшафте» // Природно-заповедный фонд – бесценное наследие Рязанщины. Мат-лы межд. конф. Рязань, 21-23 мая 2007. Рязань: изд-во РязГУ, 2007. С.107-111.
Фридман В.С., Ерёмкин Г.С., Кубарева Н.Ю. Урбанизация «диких» видов птиц: трансформация популяционных систем или адаптация особей // Журн.общ.биол. Т.59. №3. С. 207-219.
Фридман В.С., Ерёмкин Г.С., Захарова Н.Ю. Возвратная урбанизация — последний шанс на спасение уязвимых видов птиц Европы?// Russian J. of Ecosyst. Ecol. 2016. V.1. №4. http://rjee.ru/rjee-1-4-2016-3/
Фридман В.С. Глобальный экологический кризис. По материалам курса лекций «Охрана природы: Биологические основы, имитационные модели, социальные приложения». М.: URSS, 2017. 448 c.
Хански И. Ускользающий мир: экологические последствия утраты местообитаний. М.: КМК, 2010. 340 с.
Харитонов С.П. Эксперимент по восстановлению исчезнувшей колонии озёрных чаек// Редкие виды птиц Нечернозёмного Центра России. мат-лы V совещания. М.: МПГУ, 2014. С. 244-249.
Чхобадзе А.Б., Филиппов Д.А. Lycopodiella inundata и Selaginella selaginoides в Вологодской области //Бот.журн. 2013. Т.98. №4. С.515-532.
Щипанов Н.А. Популяция как единица существования вида. Мелкие млекопитающие // Зоол.журн. 2003. Т.82. №4. С. 450–469.
Щипанов Н.А. Функциональная структура популяции и видовое разнообразие. Мелкие млекопитающие // Сборник трудов Зоологического музея МГУ им. М.В. Ломоносова. 2016. Т.54. С.478—513.
Ahlering M.A., Arlt D., Matthew G. B. Et al. Research needs and recommendations for the use of conspecific-аttraction methods in the conservation of migratory songbirds //Condor. 2919. V.112. №2. Р.252–264
Alcoforado M.J., Andrade H. Global Warming and the Urban Heat Island// Urban Ecology. An International Perspective on the Interaction Between Humans and Nature. Eds. J.Marzluff et al. Springer, 2008. P.249-262.
Alberti M., Marzluff J.M., Shulenberger E. et al. Integrating Humans into Ecology: Opportunities and Challenges for Studying Urban Ecosystems// Urban Ecology. An International Perspective on the Interaction Between Humans and Nature. Eds. J.Marzluff et al. Springer, 2008. P.143-160.
Andrews J.R. Habitat selection in restored grasslands: the role of social cues in the settlement of grasshopper sparrows // Thes. for the ms degree in the graduate college of the University of illinois at urbana-champaign, 2013. 45 pp.
Arponen A., Heikkinen R.K., Paloniemi R. Et al. The importance of connectivity for agri-environment schemes// Scaling in Ecology and Biodiversity Conservation. Henle et al., K. (eds.). Sofia: Pensoft Publishers, 2014. P.161-166.
Berry B.J.L. Urbanization// Urban Ecology. An International Perspective on the Interaction Between Humans and Nature. Eds. J.Marzluff et al. Springer, 2008. P.25-49.
Balmford A., Bruner A. Cooper P. et al. Economic reasons for conserving wild nature// Science. 2002. V.297. №5583. P.550-553. Science. 2002. V.297. №5583. Р.950-953.
Bobiec A, Jaroszewicz B., Keczyński A., Szymura A., Zub K. The afterlife of a tree. Warszawa – Hajnówka, 2005. 252 pp.
Blinnikov M., Shanin A., Sobolev N., Volkova L. Gated communities of the Moscow green belt: newly segregated landscapes and the suburban Russian environment// GeoJournal. 2006. V.66. P.65–81.
Burgess E.W. The Growth of the City: An Introduction to a Research Project// Urban Ecology. An International Perspective on the Interaction Between Humans and Nature. Eds. J.Marzluff et al. Springer, 2008. P. 71-78.
Butovsky R. O., Reijnen R., Aleshenko G. M. Et al. Assessing the conservation potential of damaged peat bog networks in central and northern Meshera (central Russia) // J. Nat. Conserv. 2004. Vol. 12. P. 1–13.
Chapron G, Kaczensky P., Linnell J.D.C. et al. Recovery of large carnivores in Europe’s modern human-dominated landscapes // Science. 2014. V. 346. № 6216. P. 1517–1519.
Dearborn D.C., Kark S. Motivations for Conserving Urban Biodiversity// Conserv. Biol. 2010. V.24. №2. Р.432-440.
Elbakidze M., Angelstam P., Sobolev N. Protected Area as an Indicator of Ecological Sustainability? A Century of Development in Europe’s Boreal Forest// Ambio. 2013. V.42. P.201–214.
Ewing R.H. Characteristics, Causes, and Effects of Sprawl: A Literature Review // Urban Ecology. An International Perspective on the Interaction Between Humans and Nature. Eds. J.Marzluff et al. P. 519-532.
Ewing R.H., Schmid T., Killingsworth R., Zlot A., Raudenbush A. Relationship Between Urban Sprawl and Physical Activity, Obesity, and Morbidity // Urban Ecology. An International Perspective on the Interaction Between Humans and Nature. Eds. J.Marzluff et al. Springer, 2008. P.569-584.
Gibson L., Li Yong D., Saving two birds with one stone: solving the quandary of introduced, threatened species// Front. Ecol. Environ. 2016. doi:10.1002/fee.1449
Johnson M.T.J., Munshi-South J. Evolution of life in urban environments// Science. 2017. V.358. №607. http://dx.doi.org/10.1126/science.aam8327
Kiehl K., Kirmer A., Donath T.W., Rasran L., Hölzel N. Species introduction in restoration projects–Evaluation of different techniques for the establishment of semi-natural grasslands in Central and Northwestern Europe// Bas. & Appl. Ecol. V.11. №4. Р.285-299.
Klenke R., Mertzanis Y., Papanikolaou A., Arponen A., Mazaris A. Stay in contact: Practical assessment, maintenance, and re-establishment of regional connectivity// Scaling in Ecology and Biodiversity Conservation. Henle et al., K. (eds.). Sofia: Pensoft Publishers, 2014. P.167-172.
Kuttler W. The Urban Climate – Basic and Applied Aspects // Urban Ecology. An International Perspective on the Interaction Between Humans and Nature. Eds. J.Marzluff et al. Springer, 2008. P.233-248.
Laurance W.F., Gopalasamy R.C., O’Connell Ch.S. et al. A global strategy for road building //Nature. 2014. V.513. P.228-232. Перевод см. http://www.socialcompas.com/2015/11/30/globalnaya-strategiya-dorozhnogo-stroitelstva/
Marzluff J.M., Ewing K. Restoration of Fragmented Landscapes for the Conservation of Birds: A General Framework and Specific Recommendations for Urbanizing Landscapes// Urban Ecology. An International Perspective on the Interaction Between Humans and Nature. Eds. J.Marzluff et al. Springer, 2008. P.739-756.
McLaren J.D., Buler J.J, Schreckengost T.et al.Artificial light at night confounds broad‐scale habitat use by migrating birds// Ecology Lett. 2018. V.21.№3. Р.356-364.
Mikusiński G., Angelstam P. Economic Geography, Forest Distribution, and Woodpecker Diversity in Central Europe // Conserv. Biol. 1998. V.12. №1. Р. 200–208.
Mikusiński G. Angelstam P. Occurrence of mammals and birds with different ecological characteristics in relation to forest cover in Europe – do macroecological data make sense? // Ecol. Bull. V.51. P.265–275.
Mikusinski G., Gromadzki M., Chylarecki P. Woodpeckers as Indicators of Forest Bird Diversity// Conserv.Biol. 2001. V.15. №1. Р. 208-217.
Myczko Ł, Rosin Z.M., Skorka P., Tryjanowski P. Urbanization Level and Woodland Size Are Major Drivers of Woodpecker Species Richness and Abundance // PLoS ONE. 2014. V.9. №4. e94218. doi:10.1371/journal.pone.0094218
Rasran L., Vogt K. Ditches as species‐rich secondary habitats and refuge for meadow species in agricultural marsh grasslands//Applied Vegetation Science. 2018. V.21. №1. Р.21-32.
Pickett S.T.A., Cadenasso M.L., Grove J.M., et al. Urban Ecological Systems: Linking Terrestrial Ecological, Physical, and Socioeconomic Components of Metropolitan Areas // Urban Ecology. An International Perspective on the Interaction Between Humans and Nature. Eds. J.Marzluff et al. Springer, 2008. P. 99-122.
Rasran L., Vogt K., Jensen K. Seed content and conservation evaluation of hay material of fen grasslands // J. for Nat. Conserv. 2006. V.14. №1. Р.34-45.
Rees W., Wackernagel M. Urban Ecological Footprints: Why Cities Cannot be Sustainable—and Why They are a Key to Sustainability // Urban Ecology. An International Perspective on the Interaction Between Humans and Nature. Eds. J.Marzluff et al. Springer, 2008. P.537-566.
Ryan C.M., Jensen S.M. Scientific, Institutional, and Individual Constraints on Restoring Puget Sound Rivers// Urban Ecology. An International Perspective on the Interaction Between Humans and Nature. Eds. J.Marzluff et al. Springer, 2008. P.647-660.
Shafer C.L. Terrestrial Nature Reserve Design at the Urban/Rural Interface// Urban Ecology. An International Perspective on the Interaction Between Humans and Nature. Eds. J.Marzluff et al. Springer, 2008. P.715-735.
Soulé M.E. Land Use Planning and Wildlife Maintenance: Guidelines for Conserving Wildlife in an Urban Landscape// Urban Ecology. An International Perspective on the Interaction Between Humans and Nature. Eds. J.Marzluff et al. Springer, 2008. P.699-710
Yokohari M., Takeuchi K., Watanabe T, Yokota Sh. Beyond Greenbelts and Zoning: A New Planning Concept for the Environment of Asian Mea-Cities// Urban Ecology. An International Perspective on the Interaction Between Humans and Nature. Eds. J.Marzluff et al. Springer, 2008. P.783-796
Virzi T., Boulton R.L., Davis M.J. et al. Effectiveness of Artificial Song Playback on Influencing the Settlement Decisions of an Endangered Resident Grassland Passerine//Condor. 2012. V.114. №4. Р.846-855.
Ward M.P., Schlossberg S. Conspecific attraction and the conservation of territorial songbirds// Cons. Biol. 2004. V.18. №4. Р. 519-525.
Примечания
[1]См.также Los Angeles River Revitalization Master Plan, Los Angeles, CA, https://www.asla.org/2009awards/064.html
[2]https://en.wikipedia.org/wiki/Sustainable_gardening; наиболее общее наименование этих усилий — «реставрационная экология», см. https://en.wikipedia.org/wiki/Restoration_ecology. Наиболее известны проекты по восстановлению прерий, уничтоженных кукурузными полями, https://www.luriegarden.org/about/sustainability/ Внутри экологии выделилась специальная дисциплина — восстановительная экология, «с собственной общественной организацией — Международное общество восстановительной экологии (The Society for Ecological Restoration International — SER), собственными журналами (Restoration Ecology, Ecological Restoration) и учебниками (Хански, 2010).
[3]«…переходят в техногенные и искусственные местообитания самые разные виды. Например, из лесных вдоль железных дорог стали расти чина лесная, из лесостепных реликтов береговых осыпей и абсолютных суходолов на обочины дорог переселился лядвенец рогатый Lótus corniculátus, из болотно-береговых видов полностью перешёл в карьеры хвощ пестрый Equisétum variegatum, амфиатлантический реликт плауночек топяной Lycopodiella inundata расселился на просеках ЛЭП и вдоль трасс газо(нефте)проводов и т.д. При этом их спутники по исходным местообитаниям, тоже редкие, не смогли внедриться в нарушенные местообитания. Или не успели? Очень плохо переходят на замещающие местообитания стенотопные водно-околоводные растения и «верные» болотные виды. Короче, тут у нас в России по этому вопросу пахать и пахать, но, учитывая с какой скоростью исчезают исходные местообитания, мы можем оказаться в ситуации, когда возможность увидеть момент перехода антропофобов в антропотолерантов/филов будет упущена» (А.Б.Чхобадзе, личн.сообщ.).
[4]Эта рекреационная эксплуатация природных сообществ региона крайне полезна для горожан. Отдых на природе в течение 20 дней снижает число дней на больничном по нетрудоспособности от ОРВИ-простуд в среднем на 3-7 дней, в зависимости от конституции человека, есть и другие оздоровительные эффекты, см. «UK green spaces worth at least £30bn a year in health and welfare, report finds»// Hardian. 2 Juni 2011. https://www.theguardian.com/environment/2011/jun/02/uk-green-spaces-value. Для ландшафта же он разрушителен, в т.ч. вследствие избирательности нагрузки. Некоторые более привлекательные ландшафты притягивают большинство отдыхающих, т.ч. их биотопы разрушаются первыми. Таковы водно-болотные местообитания, берега рек и озёр, поймы и долины рек, особенно в ближнем Подмосковье, а также опушки лесных массивов, особенно светлых сосновых боров (Фридман, 2017).
[5]Первая попытка ограничения роста городов — т. н. Кампания в защиту сельской Англии 1926 г. https://en.wikipedia.org/wiki/Campaign_to_Protect_Rural_England, существенно повлиявшая на становление Сети участков выдающихся природных ландшафтов в Великобритании (Area of Outstanding Natural Beauty, 1949 г.) и появление “зелёных колец” городов в 1955 г. В СССР они появляются в 1930-х, начиная с первого Генплана Москвы 1935 г., и завершаются в 1940-е, Фридман, 2017.
[6]Сегодня понятно, что все экологические отношения («хищник-жертва», внутри- и межвидовой конкуренции и пр.) особей, популяций и видов в сообществах предваряются информационными, проявлением чего оказываются как «градиенты» (оценивающие местообитания «с точки зрения вида» по параметрам привлекательности vs риска), так и «сортировка» особей «вдоль» них. Обзор соответствующих работ см. http://www.socialcompas.com/2014/10/28/nomogeneticheskoe/
[7]См. также «Уроки волков», http://www.socialcompas.com/2014/08/18/urokei-volkov/. Другое дело, что «восстановленные» крупные хищники не могут существовать вне последних «островов» дикой природы, которых в Европе осталось совсем немного (см. классификацию местообитания для природоохранных целей в ЕС, Хански, 2010: 28-30), и пока без поддержки со стороны человека, требующей постоянного контроля и значительных затрат. Однако возвратная урбанизация даже здесь позволяет ослабить первое и уменьшить второе.
[8]Оппозиции «консерватизм–лабильность» жизненных стратегий видов и/или особей определены по В. М. Галушину и А. В. Кузнецову. Это противоположные типы приспособительных реакций популяционной системы на разнотипные «возмущения» в ареале: трансформацию местообитаний, периодическое обеднение и/или флюктуации доступности ресурсов. Консервативные виды отличает стабильная численность, низкий потенциал размножения, способность существовать на минимуме ресурсов. У них высоки требования к устойчивости ресурсной базы и минимален динамизм населения при максимальном разнообразии биотопов (и ресурсов), эксплуатируемых на занятой территории.
Лабильные виды отличает высокий потенциал размножения, резкие колебания численности, высокая изменчивость пространственных и биотопических связей. Их население быстро (без запоздания) концентрируется в местах появления массового корма, будь то размножение грызунов, вспышки численности ксило- и филлофагов, изобилие семенного корма. Они предпочитают менее структурированные и более нарушенные местообитания, с флюктуирующими ресурсами и парцеллярной структурой, сильно варьирующейся во времени и пространстве.
Почти однотипная классификация создана Н.А.Щипановым (2003, 2016) для мелких млекопитающих по соотношению контролирующей и восстанавливающей подсистем в популяции как целом. Только там оппозиции «консерватизма–лабильности» Щипанова выстраиваются на базе восстановительной реакции на истребление зверьков локальными катастрофами или, напротив, немедленного заселения и интенсивного размножения «подвижного резерва» популяции в недавно возникших, богатых кормом, но временных биотопах, там. В. М. Галушин и А. В. Кузнецов же концентрируются на механизмах динамики благополучного населения во флюктуирующей среде, экологической и социальной. В первом случае речь идет о жизнеспособности популяции во внештатной ситуации («латание дыр»), во втором – о поддержании устойчивости в «штатной ситуации».
Сообразно вышесказанному, у «диких» видов птиц нами выделены пять градаций изменения жизненной стратегии:
А. «Виды-выскочки» (термин В.В. Леоновича): лабильность максимальна сравнительно с прочими видами рода (гильдии) – и в целом, и для отдельных типов связей в пространственно-этологической структуре популяции (социальных, к местообитанию, к территории). Фрагментация местообитаний не только не снижает численности сравнительно с «материком», но даже стимулирует рост, отчего динамизм их популяций высок (Белик, 1997, цит.по: Фридман, Ерёмкин, 2009).
Б. Эвритопные и «пластичные» виды: лабильность повышена в большинстве перечисленных отношений.
В. Полуконсервативные виды – консервативные и лабильные отношения уравновешивают друг друга. Скажем, при территориальном консерватизме социальные или биотопические связи вида лабильны, и наоборот.
Г. Консервативные виды – отношения в популяционной системе консервативны, динамизм низок или отсутствует. Однако отдельные черты лабильности обязательно присутствуют в биологии; за счет них вид приспосабливается к средовой и/или демографической стохастичности.
Д. Сверхконсервативные виды – консерватизм всех типов отношений максимален, виды жестко привязаны к определенным биотопам или элементам растительности. Редкие и уязвимые виды относятся к «Г» и «Д»; большинство из них – ключевые виды коренных сообществ в соответствующих природных зонах: верховых болотах, бореальных или широколиственных лесах и т.д. (Фридман и др., 2016).
[9]Здесь суммируются 1) направленные изменения, идущие одновременно, но с разной скоростью на уровне всего города («каркас»), мозаики элементов в отдельных районах и кварталах («ткань») и форм использования (застройка, дорога, зелёные насаждения, природная территория и пр.) на разных «позициях» внутрни этих последних («ткань»), 2) флюктуационные изменения на каждом из этих уровней, причём более локальные трансформации протекают быстрей и как бы «вложены» в более генеральные. См. Мерлен, 1977; Лаппо, 1997, Sukopp, Wittig, 1998; Towards an Urban atlas…, 2002; цит.по: Фридман и др., 2016.
[10]Среди них обычно называются освещение, шум, риск гибели взрослых от проводов, стеклянных покрытий; риск гибели гнёзд и птенцов в городских кварталах и на природных территориях от регулярно случающихся перестроек/блгоустройства; повышенный риск хищничества, в первую очередь по краям «островов», их разделённость застройкой и пр., реже – гибель от перегрева при гнездовании в зданиях и сооружениях и пр. В силу всего перечисленного города при всей их привлекательности часто становятся «экологической ловушкой» для обитающих там пассивных урбанистов и проникающих новых видов из региона. Однако чем дальше зашла урбанизация, тем менее «городские» особи «подставляют себя» под действие перечисленных факторов риска (Клауснитцер, 1990; Sukopp, Wittig, 1998; Куранов, 2008; Фридман, Ерёмкин, 2009; Фридман и др., 2016).
[11] См. также «Москвичи «за» соловьёв и «против» благоустройства», http://www.socialcompas.com/2018/03/20/moskvichi-za-solovyov-i-protiv-blagoustrojstva/
[12]Есть и «банк пополнения» для такого «баланса»: т. н. пассивные урбанисты (табл.2). Таковы многие виды дендрофильной авифауны, лугов и кустарников.
[13]или хотя бы целенаправленно не пресекает их ради обслуживания интересов стройкомплекса, как в разных городах б.СССР — Москве, Харькове, Киеве, Минске, Бишкеке, а то и в Европе. См. Соболев, Волкова, 2010; Фридман, Королькова, 2013; Фридман, 2017: 406-409.
Рекомендуем прочесть
 «Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве
«Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве Основные концепции и методы восстановления природных лесов Восточной Европы
Основные концепции и методы восстановления природных лесов Восточной Европы Предисловие книги о глобальном экологическом кризисе
Предисловие книги о глобальном экологическом кризисе Модели урбанизации «диких» видов птиц
Модели урбанизации «диких» видов птиц Москвичи «за» соловьёв и против «благоустройства»
Москвичи «за» соловьёв и против «благоустройства» Картографируя города: разделение кварталов на бедные и богатые хорошо видно
Картографируя города: разделение кварталов на бедные и богатые хорошо видно Города-сада больше не будет
Города-сада больше не будет Города, география с экологией
Города, география с экологией
