

А.М.Гиляров
Долгое время в экологии господствовало представление, что виды, живущие в одном сообществе, должны обязательно занимать разные экологические ниши. Более того, сосуществование таких видов признавалось возможным лишь постольку, поскольку они различаются своим отношением к окружающей среде. Данное положение, ставшее по сути основной догмой экологии сообществ, ведет свое начало от математических моделей, предложенных еще в 1920 е годы В.Вольтеррой и А.Лоткой, и лабораторных экспериментов Г.Ф.Гаузе, проведенных в 1930е годы [см. пример опытов, прямо показывающих роль конкуренции в поддержании высокого разнообразия видов, правда, на сообществах однолетников на серпентиновых почвах, не в тропических лесах, о которых ниже; но и для ни есть соответствующие данные, деревья соседствуют друг с другом неслучайно, но так что у соседей с большей вероятностью разная жизненная стратегия. Прим.публикатора]. Согласно «принципу конкурентного исключения» (закону Гаузе) в современной его трактовке, количество видов одного трофического уровня, живущих вместе, не может превышать числа факторов, ограничивающих рост их популяций «сверху». Упрощая ситуацию, можно сказать, что каждый вид должен быть ограничен своей пищей или своим хищником.
Однако накопление новых эмпирических данных и попытки их теоретического осмысления привели к выводу, прямо противоположному основной догме: разные виды могут сосуществовать в силу сходства своих экологических характеристик. Идея на самом деле не нова. В той или иной форме она уже не раз высказывалась: например, по отношению к растительным сообществам еще в 1920 е годы нашим соотечественником Л.Г.Раменским и независимо — американским исследователем Г.Глисоном. Однако обычно подобные взгляды рассматривались как еретические и не заслуживающие серьезного внимания1.
Сейчас положение изменилось во многом благодаря работам американского эколога С.Хаббела из Университета штата Джорджия (США). Принцип, положенный им в основу новой гипотезы, гласит: виды могут сосуществовать, если в расчете на одну особь имеют примерно равные вероятности рождения, гибели, миграции и видообразования [1]. Такая экологическая эквивалентность особей разных видов и была названа нейтральностью. По собственному признанию Хаббела, формированию этих представлений немало способствовали знакомство с теорией островной биогеографии (предложенной в 1960е годы Р.Макартуром и Э.Уилсоном) и его огромный личный опыт изучения тропических лесов, сначала — сухих, листопадных в Коста-Рике, а позднее — влажных на о.Барро Колорадо (посреди оз.Гатун в зоне Панамского канала).
Мы постараемся объяснить, почему весьма абстрактная модель устройства островного сообщества и сугубо конкретные эмпирические данные о чрезвычайно высоком видовом разнообразии тропических лесов заставляют усомниться в основной догме экологии сообществ, согласно которой виды сосуществуют, поскольку занимают разные ниши.
Сомнение, однако, не есть еще отрицание, поэтому мы затронем здесь и недавно предпринятую попытку примирить разные подходы к сообществу в рамках одной модели.
Антинишевая модель Макартура—Уилсона
Содержание
Один из вопросов, который задавали себе Макартур и Уилсон, можно сформулировать так: почему на острове, как правило, видов меньше, чем на такой же площади территории материка [2]? Видимо, рассуждали авторы, на острове скорость вымирания видов выше, чем на материке, а скорость заселения новыми не столь высока, чтобы быстро компенсировать вымирание. Число видов, входящих одномоментно в островное сообщество, есть результат сбалансированности двух процессов, а именно — вселения (иммиграции) новых видов на остров и вымирания уже присутствующих. Оба процесса можно выразить как функции числа видов, имеющихся на острове (рис.1).
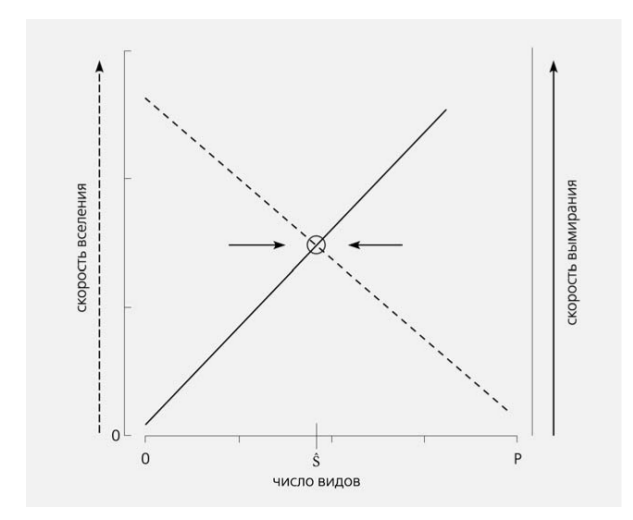
Рис.1. Модель поддержания равновесного числа видов S на острове в результате взаимодействия процессов вселения новых видов и вымирания старых. Интенсивность обоих процессов выражена как функция числа видов, имеющихся на острове. P — общий пул видов для заселения. Пересечению кривых скорости вселения и скорости вымирания соответствует некое равновесное число видов [2]. 3 0
Скорость вселения снижается, поскольку все меньше видов из исходного пула (живущих на близлежащем материке или крупном острове) останется за пределами острова. Скорость же вымирания возрастает — ведь будет кому вымирать, а кроме того, сокращение среднего размера одной популяции автоматически равносильно повышению вероятности ее случайного вымирания. Равновесное сообщество в теории островной биогеографии не есть некий сложившийся набор определенных видов, долго (в теории — неограниченно долго) сохраняющийся на острове.
Модель предполагает только равенство скоростей двух процессов — заселения и вымирания. Видовой же состав такого сообщества со временем может меняться. Интересно, что сами авторы модели на это обстоятельство особого внимания не обратили. Тогда господствовало убеждение, что сосуществование видов возможно только при разделении экологических ниш. Более того, один из авторов модели, Макартур, исследовал механизм разделения экологических ниш американских славок — мелких воробьиных птиц рода Dendroica.
Оказалось, что обитающие вместе виды этого рода собирают корм в разных частях крон хвойных деревьев: один ближе к стволу, другой — к периферии кроны, третий — к вершине, четвертый — к комлю. Позднее Макартур потратил немало усилий на теоретическое изучение максимально допустимого, т.е. разрешающего сосуществование, перекрывания экологических ниш видов.
Однако в модели островной биогеографии Макартура—Уилсона удивительным образом не фигурирует представление о необходимости расхождения видов по разным нишам (факт, справедливо отмеченный Хаббелом). Сообщество в этой модели формируется расселением организмов, а не конкуренцией между уже присутствующими, поэтому такие группировки называют сформированными расселением, в отличие от сформированных расхождением по нишам. Предполагается, что последние складываются только из тех видов, которые, вселившись, смогли занять свободные экологические ниши. Очевидно, что между вселяющимися видами должна быть конкуренция, которая и расставит все по своим местам: одни виды разойдутся по свободным нишам, а другие останутся «не у дел» и не смогут дать начало размножающимся популяциям.
На самом деле в любом сообществе всегда есть некоторое расхождение видов по разным нишам. Так, в лесу мы видим, что кроны разных видов деревьев занимают разные ярусы — одни образуют полог леса, другие — подлесок, третьи выступают над основным пологом. Корни разных видов также могут находиться на разной глубине. Однако многие виды живут бок о бок и не проявляют (их не наблюдают) сколь либо существенных различий в своих требованиях к среде. Утверждать, что все виды одного сообщества обязательно занимают разные ниши, вряд ли возможно. Особенно если речь идет о сообществах с высоким видовым разнообразием, к коим безусловно относятся влажные тропические леса.
Необъяснимое разнообразие тропического леса
Жителю умеренных широт очень непросто представить себе то разнообразие видов древесных растений, с которым он столкнется во влажном тропическом лесу. Ведь он привык, что все найденные виды деревьев в лесу можно пересчитать по пальцам. С такими крайне бедными по видовому составу лесами имели дело и лидеры основных школ в области геоботаники и фитоценологии — науки о растительных сообществах.
Отсюда и их представления о лесах, в которых преобладают один-два вида. В частности, в нашей стране такой подход развивал В.Н.Сукачев, оказавший огромное влияние на становление отечественной фитоценологии, но, справедливости ради, надо заметить, вовсе не способствовавший развитию критического взгляда на доминирующие воззрения.
Сосуществование разных видов деревьев в лесах умеренной зоны более или менее удовлетворительно объясняется в рамках традиционных представлений о нишевой организации сообществ. Кроме того, во многих случаях совместное произрастание разных видов справедливо трактуется как некое преходящее состояние, как результат еще не закончившейся смены одних растений другими в ходе обычной сукцессии. Так, в европейской части России легко наблюдать, как быстро зарастают вырубки породами пионерами (например, березой и осиной), а потом их постепенно сменяют основные лесообразующие породы — ель, сосна (на песчаных почвах) или в более южных районах — дуб.
Но позволит ли подход, оправдавший себя при изучении лесов умеренной зоны, объяснить сосуществование 100 разных видов деревьев на 1 га? А подобное разнообразие во влажных тропических лесах вовсе не редкость. Так, на о.Барро-Колорадо на участке леса в 50 га произрастает 316 видов деревьев, в Таиланде — 269 видов, на Малазийском п-ове — 823 вида, а на о.Борнео — 1186 видов (возможно, рекордное число на сегодняшний день). Причем во всех случаях к деревьям относили растения, имевшие диаметр ствола не менее 1 см на высоте груди, и не относили лианы.
Основной вопрос, на который нужно ответить, столкнувшись со столь высоким разнообразием, сводится к поиску механизма, обеспечивающего выживание множества редких видов. Иначе его можно сформулировать так: почему редкие виды не вытесняются из сообщества более многочисленными и, скорее всего, более успешными конкурентами? Ведь все они используют одни и те же ресурсы, все конкурируют прежде всего за свет, за место под солнцем в буквальном смысле этого слова. Подобный вопрос, конечно, не раз возникал, хотя чаще он только подразумевался из общего контекста. И тем не менее попытки объяснить, какие преимущества могла бы давать видам их малочисленность, время от времени предпринимались.
Одно из таких объяснений — гипотеза Джанзена—Коннела: ее в несколько разных терминах независимо сформулировали в 1970 г. два американских исследователя — Д.Джанзен и Ж.Коннел. Суть гипотезы сводится к тому, что в тропических лесах с очень высоким разнообразием деревьев семена и проростки имеют гораздо больше шансов выжить, если они находятся вдали от родительских растений. Джанзен полагал, что значительную часть семян, упавших рядом с «родителем», поедают специализированные «хищники» (на самом деле растительноядные животные), которым гораздо легче разыскать свои излюбленные кормовые объекты, если они располагаются скоплениями. Коннел считал, что усиленное истребление более массовых видов деревьев происходит чуть позже — на стадии проростков, и решающую роль играют листогрызущие насекомые, например, гусеницы тех или иных видов бабочек, кормящиеся определенными видами.
Заметим, что и Джанзени Коннел исходили из предположения о строгой пищевой специализации животных тропических лесов. Считалось, что у каждого растения есть свои потребители, которые в свою очередь контролируются специфическими хищниками и паразитами. Сразу заметим, что каких либо серьезных эмпирических данных, подкрепляющих эти предположения, тогда не было. Позднее же чешский энтомолог В.Новотный, работавший с большой группой помощников в тропическом лесу Новой Гвинеи, на основе огромного материала убедительно доказал, что подавляющее большинство листогрызущих насекомых вовсе не узкие специалисты,а питаются по крайней мере несколькими видами растений [3, 4]. Более того — средний уровень специализации насекомых в тропиках оказался не выше, чем у насекомых, обитающих в широколиственных лесах Центральной Европы.
Надо сказать, что строгая проверка гипотезы Джанзена— Коннела в полевых условиях дело очень непростое. Результаты получались порой противоречивыми, а кроме того, не прекращались споры о том, каковы должны быть размеры площадок для учета численности растений разного возраста. Однако проведенное недавно по единой схеме крупномасштабное изучение семи участков тропических лесов (два в Центральной Америке и пять в Юго Восточной Азии) позволило сделать важный вывод: по мере роста и развития деревьев (от семян до зрелых растений) их разнообразие увеличивается, а позднее, на стадии старения и отмирания, снижается [5]. Разнообразие оценивалось как число видов с определенным числом особей, причем анализ проводился отдельно для четырех возрастных групп деревьев и на пробных площадках разной площади. Интервал между двумя обследованиями составил 5 лет, а для двух участков — 10 лет (с промежуточным обследованием через 5 лет).
Как заметили авторы этой работы, обнаруженную ими лучшую выживаемость редких видов в сравнении с массовыми можно объяснить не только гипотезой Джанзена—Коннела. Не исключено, что эффект связан с некоторым разделением экологических ниш, в результате чего интенсивность внутривидовой конкуренции становится ниже межвидовой. Еще одна возможность — положительное влияние соседних растений, если они оказываются другого вида (это так называемая гипотеза облегчения) [и наоборот. Прим.публикатора]. Но каковы бы ни были конкретные процессы, происходящие в сообществе, очевидно, что у редких видов вероятность выживания на ранних стадиях выше, чем у массовых.
Относительное обилие видов
Среди экологов никогда не прекращался спор о том, что есть сообщество — высоко интегрированная система, развивающаяся и реагирующая на изменения среды как нечто целое (порой напоминающее организм), или же это достаточно произвольно выделяемая исследователями совокупность особей разных видов, волею судеб оказавшихся соседями, у которых особенности физиологии, строения и жизненного цикла позволяют существовать на определенной территории. Тот же вопрос можно сформулировать несколько иначе: что важнее для поддержания структуры и функционирования сообщества — система взаимосвязей между видами, жестко ограничивающая индивидуумы разных видов, или же свойства отдельных видов, позволяющие им выживать в ряду поколений вне зависимости от того, какие виды соседствуют с ними.
Подчеркнем еще раз, что речь идет только о сообществах, образованных организмами одного трофического уровня (их иногда называют гильдиями). Мы не рассматриваем те, в которых продукты выделения одной группы организмов служат пищевыми ресурсами для другой (как это часто бывает в бактериальных системах, в частности в разных биопленках). Фактически это уже экосистемы, и принципы их организации совсем другие.
В своих попытках охарактеризовать природу сообществ исследователи всегда стремились распознать их структуру, вычленить общие черты организации для самых разных группировок. В частности, задолго до моды на слово «биоразнообразие», означающее все, что угодно, и потому особого смысла не имеющее, экологи изучали видовое разнообразие сообществ и пытались выяснить, как зависит число видов от размера изучаемой выборки (числа особей в пробе) или как соотносятся между собой численности (или биомассы) разных видов, входящих в одно сообщество.
Интуитивно ясно, что чем больше проба, тем больше видов можно в ней обнаружить, но каков характер этой зависимости и насколько она универсальна? Можно ли найти некий показатель разнообразия, не просто оценивающий число видов, но и учитывающий соотношение численностей разных видов, меняющийся в зависимости от того, сколь сильно выражено доминирование немногих видов?
Среди работ, пытающихся ответить на подобные вопросы, особого внимания заслуживает статья трех британских ученых: крупнейшего специалиста по статистике Р.Фишера и двух энтомологов — А.Корбета и К.Уильямса [6]. В основе их исследования — коллекция дневных бабочек, собранных Корбетом в Малайзии (поскольку отлавливались все встреченные экземпляры, коллекция давала представление об относительной численности разных видов), а самое главное, — результаты лова ночных бабочек специальными световыми ловушками, проведенного Уильямсом в Англии2.
Пойманных бабочек определяли до вида и подсчитывали (мелких молей не учитывали, ограничиваясь только более крупными формами — Macrolepidoptera). Когда выборка стала достаточно большой (работа велась несколько лет, и счет пошел на сотни, а потом и на тысячи экземпляров), виды разбили на классы обилия — по количеству особей, которыми они были представлены.
![Рис.2. Распределение числа видов по классам обилия для выборок ночных бабочек, пойманных на Ротамстедской станции. Внизу — данные за четыре года; в середине — за один год в среднем; вверху — за 1/8 года в среднем. Пунктирная линия — аппроксимация логарифмическими рядами. Больше всего малочисленных видов. Числа со стрелками указывают на число особей самого многочисленного вида в выборке. Классы обилия не логарифмические [13].](http://www.socialcompas.com/wp-content/uploads/2019/02/Snimok-ekrana-ot-2019-02-07-14-43-41.png)
Рис.2. Распределение числа видов по классам обилия для выборок ночных бабочек, пойманных на Ротамстедской станции. Внизу — данные за четыре года; в середине — за один год в среднем; вверху — за 1/8 года в среднем. Пунктирная линия — аппроксимация логарифмическими рядами. Больше всего малочисленных видов. Числа со стрелками указывают на число особей самого многочисленного вида в выборке. Классы обилия не логарифмические [13].
Больше всего оказалось видов, в суммарной выборке представленных одним единственным или несколькими экземплярами, более многочисленных видов было меньше, а самых массовых — еще меньше (рис.2). Полученный эмпирически ряд убывания числа видов по мере возрастания их численности Фишер предложил описывать логарифмическими рядами. На основе такого статистического распределения разработали индекс разнообразия, количественнооценивающий скорость уменьшения числа видов в выборке по мере увеличения числа особей.
Однако эта работа, получившая широкую известность, вскоре подверглась критике со стороны американского исследователя Ф.Престона, который заметил, что при увеличении размера выборки характер статистического распределения, описывающего соотношение частот видов с разной численностью, меняется: из логарифмического становится логнормальным [7]. Для разбивки видов по классам обилия Престон использовал неравномерную шкалу — так называемые октавы. В первую октаву входили виды, представленные одной особью, во вторую — двумя, в третью — четырьмя, в четвертую — восемью, в пятую — 16, затем — 32, 64, 128 особями и т.д. (в обсуждаемой работе они соответствовали верхним границам каждого класса).
Выяснилось, что кривая, показывающая распределение числа видов, объединенных в октавы обилия, имеет вид колокола, напоминающего классическую гауссову кривую нормального распределения (рис.3). Больше всего оказалось видов, представленных не одной особью, а несколько более много численных. Правда, левая часть графика была как бы отрезана.
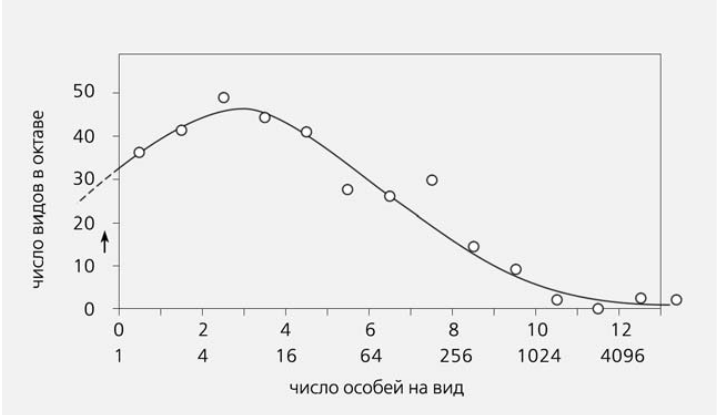
Рис.3. Распределение числа видов бабочек по логарифмическим классам обилия (октавам). За четыре года в местечке Ороно в США (штат Мэн) поймана 56 131 особь [7].
Создавалось впечатление, что общая выборка еще недостаточно велика, чтобы в нее попали самые редкие виды. По мере же ее увеличения должен добавляться «хвост редких видов» и таким образом достраиваться недостающая левая часть графика. В отношении бабочек, пойманных на Ротамстедской станции, примерно так оно и происходило. Их ловили еще несколько лет, полученные данные объединялись в одну большую выборку, в ней появлялись новые редкие виды, а виды, ранее представленные одной особью, становились более многочисленными и переходили в следующий класс обилия (рис.4). Правда, левый хвост кривой, соответствующей самым редким видам, все равно был отрезан.
![Рис.4. Число видов по логарифмическим классам обилия (октавам). С увеличением выборки (от 1/8 года до 8 лет) распределение больше становится похожим на логнормальное. Хвост редких видов еще не выявлен полностью, и левая часть гистограммы срезана. По данным Уильмса и др. [1].](http://www.socialcompas.com/wp-content/uploads/2019/02/Snimok-ekrana-ot-2019-02-07-14-44-54.png)
Рис.4. Число видов по логарифмическим классам обилия (октавам). С увеличением выборки (от 1/8 года до 8 лет) распределение больше становится похожим на логнормальное. Хвост редких видов еще не выявлен полностью, и левая часть гистограммы срезана. По данным Уильмса и др. [1].
Хотя работа Престона была хорошо известна специалистам, идея использовать логнормальное распределение для описания соотношения численностей разных видов широкого распространения не получила. Отчасти, видимо, потому, что данная зависимость проявлялась только в очень больших выборках из множества особей, относящихся к разным видам. Такие выборки были скорее не сообществами, а «таксоценами» — совокупностями организмов определенной таксономической группы (например, всех бабочек, или всех птиц), обитающих на какой то территории, нередко довольно обширной, включающей разные биотопы. Конечно, любая таксономическая группа организмов в самых общих чертах есть и некое экологическое единство.
Кроме того, некоторые авторы (в частности, Р.Мэй), подчеркивали, что логнормальное распределение если и представляет какой-то интерес для изучения структуры сообщества, то только как некая точка отсчета. Само же по себе оно не свидетельствует о наличии какой-то специфической структуры сообщества, а получается как результат независимого воздействия многих случайных факторов на величину, меняющуюся путем умножения. В том, что любая популяция растет именно так, легко убедиться, построив график роста популяции бактерий, берущей начало от одной клетки, которая делится, к примеру, раз в час. Если следовать этой логике (а почему бы ей не следовать), то соответствие распределения частот встречаемости видов разной численности логнормальному распределению можно трактовать как свидетельство отсутствия какой-то специфической структуры сообщества и прекратить дискуссию. Но этого не произошло.
Доводы, базирующиеся только на математической статистике, не очень-то убеждали биологов. Скорее всего у них оставалась некая внутренняя убежденность, что все не так просто и взаимодействия видов порой становятся важной силой, влияющей на формирование структуры сообщества. Спустя более чем четверть века после публикации работы Мэя [8] логнормальное распределение снова оказалось в центре внимания, на этот раз в связи с появлением нейтральной модели.
Противостояние и примирение
Выдвинув принцип нейтральности как основу сосуществования видов, Хаббел разработал модель, предсказывающую относительное обилие видов в сообществе. Помимо собственно нейтральности (т.е. одинаковой для разных видов удельной рождаемости, смертности и скорости колонизации) в этой модели важны были еще два условия. Во-первых, объект изучения, конкретное локальное сообщество, рассматривали как часть метасообщества, содержащего весь пул видов и служащего источником для восполнения тех из них, которые по каким-то причинам вымерли в локальных сообществах. Иначе говоря, метасообщество — это нечто вроде материка или большого архипелага островов по отношению к небольшому изучаемому острову.
Во-вторых, принимали, что в сообществе существует некоторое конечное число мест, которые могут быть заняты особями. Если известно, к примеру, что численность какого-то вида в сообществе возрастает, это автоматически означает снижение численности других видов. На примере деревьев тропического леса на Барро Колорадо Хаббел показал, что его модель подтверждается реальной картиной распределения частот встречаемости видов с разной численностью. При этом использовалась весьма солидная выборка — 21 457 деревьев, относящихся к 225 видам. Для сравнения та же совокупность эмпирических данных была аппроксимирована логнормальным распределением. Оба статистических распределения оказались весьма похожи (рис.5), но логнормальное распределение все же недооценивало число самых редких видов, представленных в выборке одним или несколькими экземплярами.
По поводу модели Хаббела и ее применения для анализа относительного обилия видов разгорелась жаркая дискуссия на страницах журнала «Nature». В частности, Б.Макгилл утверждал, что логнормальное распределение, не требующее каких-либо гипотез о нейтральности видов и случайном дрейфе, нужно рассматривать как основное, соответствующее правилу Оккама — не умножать сущности сверх надобности [9]. Однако предлагаемые Макгиллом численные методы проверки подразумевали довольно большую статистическую погрешность и были весьма уязвимы для критики. На помощь Хаббелу при шли специалисты, владеющие более изощренным математическим аппаратом. Так, И.Волков (физический факультет Университета штата Пенсильвания) совместно с коллегами предложил аналитическое решение задачи в модели Хаббела, показав, что она лучше описывает число самых редких видов [10].
По мнению Волкова и его соавторов, модели Хаббела следует отдать предпочтение, поскольку она исходит из биологически обоснованных предположений и реально подразумевает определенный механизм формирования сообщества. Пока спор этот продолжается, и точка в нем еще не поставлена. На наш взгляд, вне зависимости от его исхода попытка примирить в одной модели представления о двух принципиально разных механизмах сосуществования видов безусловно заслуживает внимания.
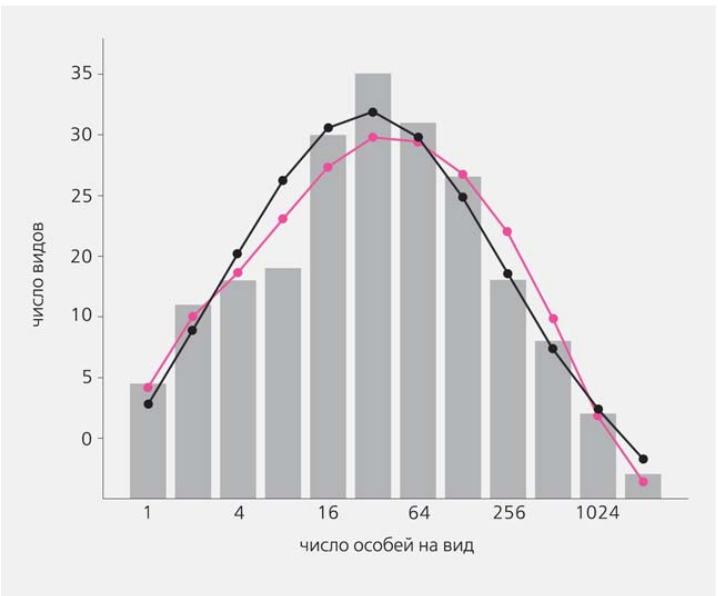
Рис.5. Распределение числа видов деревьев в тропическом лесу на Барро Колорадо по логарифмическим классам обилия (октавам). Черной линией показана аппроксимация логнормальным распределением, а цветной — многомерным распределением Хаббела. Оба распределения достаточно хорошо описывают эмпирические данные, но распределение Хаббела лучше для редких видов (левая часть графика) [10].
П.Чессон обозначил их как выравнивание и стабилизация [11]. Нейтральная модель Хаббела основана на сближении видов по своим демографическим характеристикам, прежде всего по удельной (в расчете на особь) рождаемости, смертности и скорости колонизации, т.е. на выравнивании. Если, например, уменьшается разность рождаемости и смертности видов, шанс их сосуществования увеличивается. Стабилизация предполагает разделение ниш, но о его эффективности судят не по способу, которым оно достигается (к примеру, расположением ли корней на разной глубине, или разной чувствительностью к нехватке тех или иных элементов питания), а по достигнутому результату. Результатом же считается усиление внутривидовой конкуренции по сравнению с межвидовой по мере того, как растет численное доминирование вида в сообществе. Так, если какой-то редкий вид становится все более массовым (а при наличии постоянного числа «мест» в сообществе это означает и увеличение его относительного обилия), скорость его популяционного роста должна снижаться.
Суть стабилизации легко понять из графической модели зависимости удельной скорости популяционного роста какого то вида от его относительной численности в сообществе, или иначе — частоты встречаемости (рис.6). В отсутствие стабилизации скорость популяционного роста будет оставаться неизменной, не зависящей от того, редкий или массовый данный вид3.
При стабилизации (она может трактоваться как косвенное свидетельство разделения ниш) удельная скорость популяционного роста будет снижаться по мере увеличения относительной численности данного вида. Чем больше наклон соответствующей линии, тем сильнее выражена стабилизация.
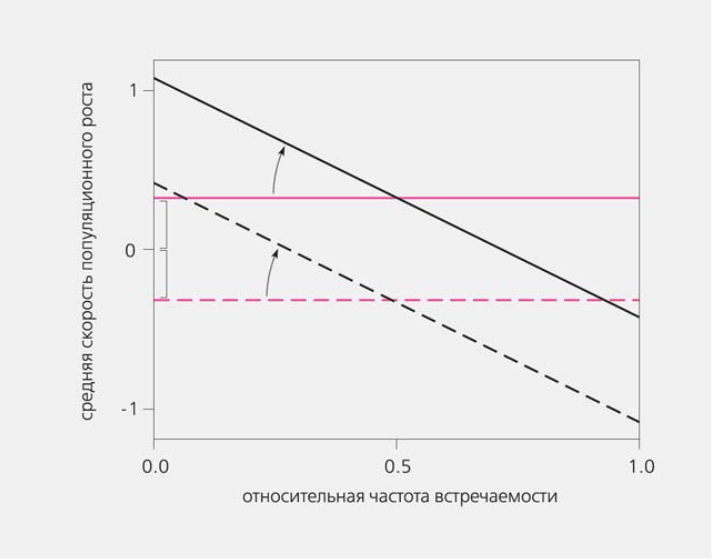
Рис.6. Предполагаемая взаимосвязь между средней удельной скоростью роста популяции и относительной частотой встречаемости того же вида в сообществе. Два гипотетических вида показаны разными линиями (черной и цветной). В левой части графика вид редок, в правой — доминирует, вплоть до исключения всех остальных (1.0). Если по мере возрастания частоты встречаемости вида скорость его роста не меняется (прямые идут параллельно оси Х), механизма, специально сдерживающего рост доминанта, нет, и вероятность сосуществования двух видов тем выше, чем ближе их удельные скорости (фигурные скобки) и чем ближе друг к другу линии разных видов. Этот механизм признается основным сторонниками гипотезы нейтральности. Если же с увеличением частоты встречаемости вида скорость его роста падает (наклонные прямые), значит, существует механизм стабилизации, не допускающий вытеснения данным видом других. Большей степени стабилизации соответствует больший отрицательный наклон (показано стрелками). Наличие механизма стабилизации может трактоваться как свидетельство разделения ниш [12].
П.Эдлер с соавторами, развивая представления Чессона, предложил графическую модель сосуществования видов, объединяющую механизмы выравнивания и стабилизации [12]. Для этого на одном поле графика (рис.7) были нанесены: по оси X — сила стабилизации (т.е. степень наклона прямой на рис.6), а по оси Y — степень сходства видов по скорости их популяционного роста (значение 1.0 соответствует полной идентичности видов по данному демографическому параметру). Область стабильного сосуществования видов — это верхняя правая часть графика, а область конкурентного исключения — нижняя левая. Обозначенные буквами точки соответствуют различной комбинации выравнивания и стабилизации — двух способов достижения сосуществования конкурирующих видов. Чем более схожи виды по своим демографическим характеристикам (чем больше значение Y), тем больше шансов на их сосуществование: точка A — исключительно за счет сходства демографических характеристик конкурирующих видов; точка E — преимущественно за счет стабилизирующего механизма; точка B предполагает устойчивое сосуществование, поскольку к сходству значений скорости популяционного роста добавляется стабилизирующий механизм; в точке C один вид будет вытесняться другим, но достаточно медленно, поскольку их демографические характеристики схожи; в точке D такое вытеснение произойдет скорее всего очень быстро; в точке F стабилизирующий механизм хорошо развит, но различие в демографических характеристиках слишком большое для устойчивого сосуществования.
Описанная «объединительная» модель Чессона—Эдлера опубликована совсем недавно, и пока трудно судить, насколько она поможет сближению разных точек зрения на механизм формирования сообществ.
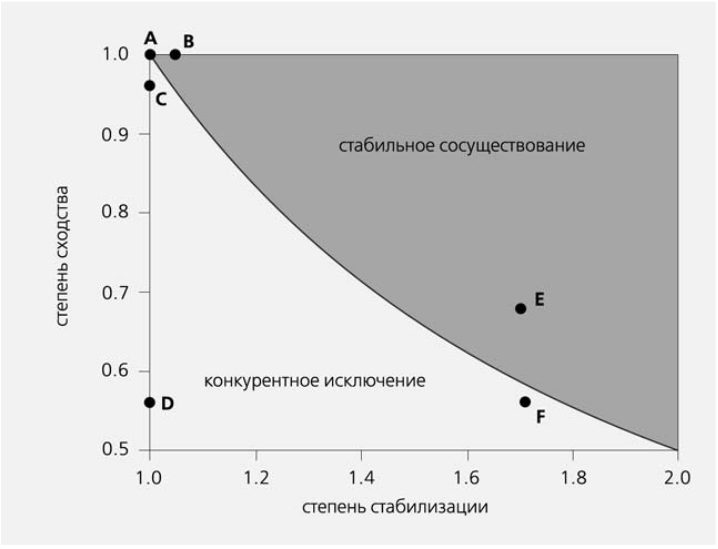
Рис.7. Схема, объединяющая механизмы выравнивания (степень сходства удельной скорости популяционного роста двух конкурирующих видов) и стабилизации (насколько быстро снижается скорость популяционного роста вида по мере возрастания его частоты встречаемости). Чем больше выравнивание (степень сходства), тем меньше потребность в стабилизирующем механизме для сосуществования видов. Если же их скорости популяционного роста сильно различаются (малые значения степени сходства), то сосуществование возможно только за счет разделения ниш (большей стабилизации). Область сосуществования — правая верхняя часть поля графика, а область конкурентного исключения — нижняя левая [12].
Прогресс в вопросах
Прогресс безусловно есть, и заключается он прежде всего в изменении характера задаваемых вопросов. Экологи наконец подошли к тому, чтобы задавать вопросы, на которые можно получить недвусмысленные ответы. На первый взгляд, эти вопросы могут показаться слишком простыми и частными. Например, существует ли отрицательная зависимость удельной скорости популяционного роста вида от относительной частоты его встречаемости? Отличаются ли локально сосуществующие виды большим сходством демографических характеристик в сравнении с видами случайной выборки из метасообщества? Меняется ли видовое разнообразие сообщества, если мы оценим его отдельно для выборок особей разного возраста?
Но получить ответы на эти вопросы очень непросто. Для этого, как правило, требуются весьма трудоемкие полевые исследования, нередко и постановка опытов, а также вспомогательные математические модели. И тем не менее затраченные усилия оправданы, поскольку полученные ответы имеют общее значение: позволяют принять одну гипотезу и, что еще важнее, — отказаться от других. Мне кажется, что наблюдаемое сейчас оживление интереса к изучению сообществ есть не просто преходящая мода, а достижение действительно некого нового важного этапа.
Эпоха продолжавшегося с начала 1920-х годов плюрализма, параллельного существования разных школ, представители которых сохраняли свою независимость и никоим образом не вторгались в чужую епархию, кажется, близится к концу, и это нельзя не приветствовать. Сознаю, что, осуждая затянувшийся плюрализм, неизбежно навлеку на себя гнев многих экологов, особенно тех, кто считает необходимым «сохранять верность традициям», под которыми подразумевается обычно не что иное, как запрет критики «мэтров» и боязнь взглянуть на проблему с совершенно другой стороны. Не буду называть никаких школ, не хочу невольно обидеть ни их основателей, ни адептов.
Ограничусь только всеобщим призывом к освоению сделанного в последние годы и к постановке новых вопросов, последовательно продвигающих нас к пониманию действительно общих механизмов формирования и поддержания биотических сообществ.
Литература
Hubbell S.P. The Unified Neutral Theory of Biodiversity and Biogeography. Princeton; Oxford, 2001.
MacArthur R.H., Wilson E.O. The theory of island biogeography. Princeton; N.J., 1967.
MacArthur R.H. // Ecology. 1958. V.39. P.686—706.
Novotny V., Drozd P., Miller S.E. et al. // Science. 2006. V.313. P.1115—1118.
Wills C., Harms K.E., Condit R. // Science. 2006. V.311. P.527—531.
Fisher R.A., Corbet A.S., Williams C.B. // J. Anim. Ecol. 1943. V.12. P.42—58.
Preston F.W. // Ecology. 1948. V.29. P.254—283.
May R.M. Patterns of species abundanсe and diversity // Ecology and Evolution of Communities / Ed. M.L.Cody,
J.M.Diamond. Cambridge, 1975. P.81—120.
McGill B.J. // Nature. 2003. V.422. P.881—885.
Volkov I. et al. // Nature. 2003. V.424. P.1035—1037.
Chesson P. // Annual Review of Ecology and Systematics. 2000. V.31. P.343—366.
Adler P.B., HilleRisLambers J., Levine J.M. // Ecology Letters. 2007. V.10. P.95—104.
Williams C.B. Patterns in the balance of nature. L., 1964.
Источник Природа. 2007. №11. С.29-37.
1Автор этих строк в начале 1980 х годов опубликовал две статьи, в которых показал, что два вида дафний сосуществу ют в одном водоеме именно благодаря тому, что сходно реагируют изменениями рождаемости на смену трофических условий в водоеме (Гиляров А.М. // Доклады АН СССР. 1981. Т.257. С.251—254; Ghilarov A.M. // Oikos. 1984. V.43. P.46—52).
На эти статьи почти не было реакции, а эколог и специалист по теории эволюции М.Линч, прочитав статью, заметил, что трудно представить себе ситуацию, когда отбор у двух видов, направленный в одну сторону, приводит к конвергенции, а не дивергенции сосуществующих видов.
2Ловы проводили на расположенной неподалеку от Лондона Ротамстедской экспериментальной станции. В историю науки эта станция вошла прежде всего как место длительных экспериментов по применению удобрений, где Фишер для обработки полученных данных предложил методы факторного анализа.
3Подчеркнем, что речь идет об удельной скорости популяционного роста («на душу населения»): абсолютная скорость прироста, т.е. на сколько особей увеличилась популяция, конечно, будет становиться больше по мере роста численности.
Рекомендуем прочесть
 «Ловушки», эволюционные и экологические
«Ловушки», эволюционные и экологические Методология изучения рекреационного воздействия на городские леса и подходы к проектированию рекреационных объектов
Методология изучения рекреационного воздействия на городские леса и подходы к проектированию рекреационных объектов Экологические аспекты гидростроительства и идеи Виттфогеля
Экологические аспекты гидростроительства и идеи Виттфогеля Международная торговля угрожает биоразнообразию развивающихся стран
Международная торговля угрожает биоразнообразию развивающихся стран Зачем нам в России «английский» газон?
Зачем нам в России «английский» газон? Птицы и муравьи не мешают друг другу есть гусениц
Птицы и муравьи не мешают друг другу есть гусениц Паразитические манипуляции фенотипом хозяина: эффекты во внутренней и внешней среде
Паразитические манипуляции фенотипом хозяина: эффекты во внутренней и внешней среде Перестройка в экологии: от описания видимого к пониманию скрытого
Перестройка в экологии: от описания видимого к пониманию скрытого
