Она может быть интересна и коллегам, т.к. суммирует аргументы, почему даже симметричная модель полового отбора работает ну не очень, ухаживание и образование пар неотделимо от других форм социальной жизни с её приматом системности (воспроизводства видовой структуры отношений) над адаптивностью (выигрышем в приспособленности индивидов) — или проигрышем. А они важны для теории эволюции вообще.
1. При всякой попытке посчитать реальные затраты самцов и самок на заботу о потомстве (т.н. parental investment) в единицах времени и/или энергии неизменно оказывается, что «вложения» в конкуренцию за партнёра (прекопуляторные затраты) и вложения в собственно репродукцию неразделимы, т.к.одни и те же формы активности участвуют в том и другом одновременно. Например,
б) местообитания и кормовые запасы, охраняемые самцом в рамках территориальной активности – лишь отчасти способ выделиться среди самцов, став таким образом наиболее привлекательным для самок. В случае полифункциональной территории «типа А» (Панов, 1983, с.***) это и форма заботы о потомстве: самец расходует время и энергию,, монополизируя «для потомства» подходящие кормовые местообитания. Действительно, у многих видов птиц самки выбирают именно территорию, а отнюдь не самца, и достаточно тщательно, посещая разные участки для сравнения. Во многих исследованиях кормодобывания также показано, что размер территории весной определяется видами на прокормление «среднестатистического выводка» летом, а не удобством привлечения самок и необходимостью уединения с ними на обширном пространстве для спаривания вот прямо сейчас (чтобы токующие соседи не беспокоили вторжениями, они это делают и процесс срывается).
Это т.н. супертерритории: они занимаются весной так, «чтобы» летом, при другой погоде и с другими кормами, обеспечивали последним пару и 5-6 птенцов, почему в смысле запасов корма заведомо избыточны (как и затрат усилий по их охране). Скажем, таковы они у тиранна Empidonax trailiii и много ещё у кого.

Амос Захави, великий израильский эволюционист, орнитолог, создатель местного общества охраны природы
Конечно, через качество территории как местообитания, кормового и гнездового, самки также выбирают самца.
Правда косвенно, в соответствии с принципом гандикапа Амоса Захави — самая разумная модель отбора на увеличение брачных украшений и, шире, механизмов оценки качества и выбора парнёров в половом отборе вообще. Ещё это, наверно, единственная социобиологическая концепция, которая «имеет тенденцию подтверждаться», а не опровергаться новыми данными, у всех остальных с этим очень и очень плохо.
2. Следовательно, что половой отбор неотделим от естественного. Привлекательность партнёра, «оцениваемая» первым, обычно зависит от разных факторов, оцениваемых вторым — выживания, успеха в занятии территорий, социального ранга и пр. событий внегнездового времени. См. пример с «галстучком» домовых воробьёв и подобными «метками иерархии» у других видов. В самом деле, согласно Захави носители затратных и рискованных приспособлений, исполнители вычурных, неестественных и не менее рискованных поз предпочитаются самками из контринтуитивных оценок их качества («коль может так тратиться и рисковать, значит он сверхэффективен в добыче корма, или в обеспечении безопасности вне тока»), т. е. «оценивание» половым отбором зависит от «работы» отбора естественного. А она произошла раньше, во внегнездовое время, или происходит одновременно, но в «неполовой» сфере, скажем, в «продвижении» по лестнице иерархии в группе.
3. Верно и обратное: тип брачных отношений, число партнёров, формы их привлечения, выбирающими или выбираемыми будут самцы (или самки) — все эти важные обстоятельства строения mating system, все те, что правило Бейтмана пробует объяснять, задаются не столько полом, сколько внешними обстоятельствами, социальными и экологическими, в том числе вообще не относящимися к брачной активности, а часто и вовсе случившиеся в нерепродуктивный период.
При определении того, какой пол у данного вида окажется конкурентным, а какой — заботливым–выбирающим, в какой пропорции оба партнёра «вкладываются» в заботу о потомстве и пр. , критически важны три фактора, никак ею учитываемые и даже отрицаемые, ибо представляют собой не всеобщие правила, но локальные зависимости, следующие из типологических экстраполяций в пределах конкретных филогенетических ветвей.
Это, во-первых, тип пространственно–этологической структуры (формы демонстраций и формы отношений данного вида), причём часто в аспектах, не связанных с собственно ухаживанием, скажем у пёстрых дятлов — с охраной объединённого участка пары (т. н. «натравливание» доминирующим в паре партнёром подчинённого на соседей-потенциальных захватчиков у пёстрых дятлов). Во-вторых, важны, хоть и несколько менее, сложившиеся тренды её преобразования в той филогенетической ветви, куда относится вид.
Так, считавшиеся социобиологами «экзотическими» системы размножения – полиандрия, полигинандрия, обычно с реверсией половых ролей, сдвоенное гнездование и пр. – у птиц, напротив, первичны для всего класса, будучи перепредставлены в примитивных группах, где выступают предшественником моногамии: у бескилевых, сорных кур, трёхпёрсток, примитивных ржанкообразных: якан, цветных бекасов, белых ржанок, рачьих ржанок и пр.
С другой стороны, у ряда видов из молодых и продвинутых семейств того же отряда, вроде хрустана Eudromias morinellus, пятнистого перевозчика Achtitis macularia, некоторых песочников, зуйков и чибисов, тот же тип отношений вторичен, развивается из моногамии через хорошо различимый этап «сдвоенного гнездования» (double nesting, последовательной полиандрии, где одна кладка насиживается самкой, другая — вторым самцом, см. про него у больших пёстрых дятлов).

Источник табл.1-2 — Томкович, 1984. За прошедшие 30 лет полиандрия была надёжно показана у
С третьей, по тому какой пол будет «конкурирующим», какой «выбирающим», экологически и таксономически близкие виды птиц, рыб, амфибий и млекопитающих часто различны «на все 100″, см. про близкие виды полёвок. Причём, как именно будет в каждом отдельном случае, определяется особенностями «конструкции» социальной системы каждого из них, т. е. частными обстоятельствами, а не общим правилом для всех таких случаев (и уж точно не асимметричной версии полового отбора претендовать на роль этого правила, по причинам см.что выше, что далее).
4. Не подтверждается самое очевидное следствие этой теории, связанное с «выигрышем» и «проигрышем» в «войне полов»: согласно ей «более конкурентный» пол использует репродукцию «выбирающего» пола как средство для «тиражирования» в нисходящем колене своих генов в большей степени,чем наоборот. И чем сильней конкуренция и эффективней отбор «лучших» среди первых, тем диспропорциональней роли тех и других в репродукции. Первые, чем дальше, тем больше вкладываются в «брачные украшения» и другие специализированные (поэтому «дорогие») средства конкурентной борьбы, вторые, напротив, в выращивание общего потомства.
И пока не созрел соцзаказ на обоснование пошатнувшегося неравенства, асимметричную модель и «правило Бейтмана» вспоминали в основном для того, чтобы попинать. Тема полового отбора занимала прочное, но скромное место, и интересовала только орнитологов и других специалистов по группам, где актуально понимание эволюции «брачных украшений», сложного ухаживания и заботы о потомстве. Причём половой отбор рассматривался в основном по симметричной модели Дж.Хаксли 1930-х гг.
Согласно ей, самцы и самки совместно «вкладываются» в обмен брачными демонстрациями, реализующий видовой инстинкт, и в прочие формы участия в репродукции, асимметрия которой, если есть, следует из биологии данного вида и его родственников, а не из каких-либо общих правил, противопоставляющих самцов самкам. Характерный пример — чомга и другие поганки, например, красношейная.
Ангус Джон Бейтман, генетик растений по специальности, выполнил эту — почти единственную зоологическую — работу именно для оппонирования идеям Хаксли, казавшихся ему непригодными для растений. И он совсем не имел в виду объяснять «женско-мужские различия» у человека, диморфизм заботы о потомстве у животных, и прочее, к чему пристёгивают эти опыты сегодня.

Ангус Дж. Бейтман
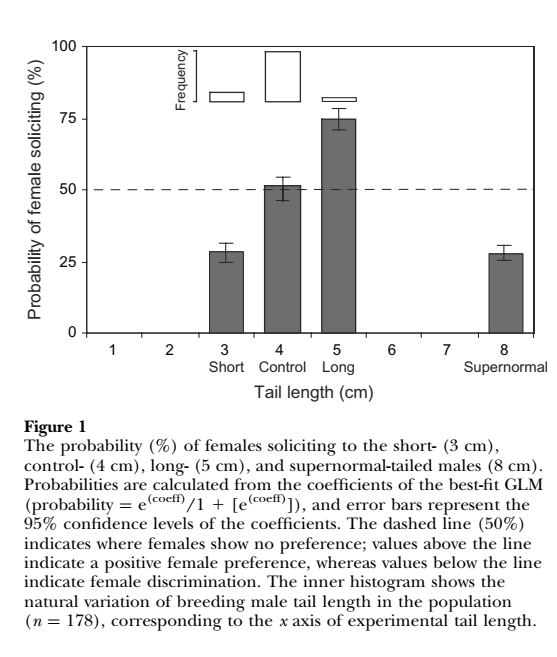
5. Не подтверждаются [предсказанные асимметричной моделью] ни сплав большей конкурентности с большей промискуитетностью у самцов, ни сопряжённый с ним сплав избирательности с верностью у самок, для видов без реверсии половых ролей (притом что само понятие роли дезавуируется критически важной зависимостью вложений в разные аспекты репродукции от территориальности сейчас до иерархии в зимних стаях). На деле всё прямо наоборот – «более сексапильные самцы», первыми образующие пары и самые предпочитаемые в сравнении с прочими, оказываются и самыми верными супругами (данные по большим синицам Parus majot), а игнорируемые поначалу, напротив, оказываются самыми промискуитетными, также как клептогамные самцы–неудачники на токах. Т.о., спаривания на последних вполне симметричны: полигинии наиболее предпочитаемых самками центральных самцов соответствует полиандрия наиболее активных (легче привлекаемых током) самок, см. данные по турухтану Phylomachus pugnax, почему система спаривания в целом характеризуется как полигинандрия. 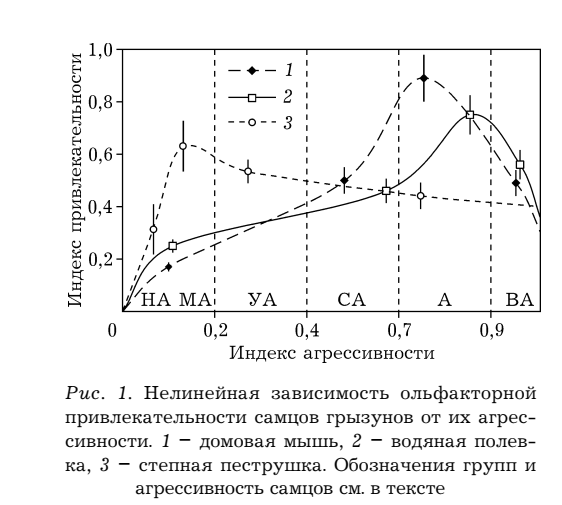
А самки при выборе оценивают не столько «мощь» самцов в конкуренции между собой (более или менее сцепленную с агрессией), сколько «обращение» с ними самими, с агрессией и активностью коррелирующее отрицательно, и положительно — с интенсивностью брачных демонстраций без сближения, и аффилиативного поведения, провоцирующего более тесное сближение без риска срыва или агрессии. При возможности выбора они предпочтут побеждённых, не победителей , чем обеспечивают успех рейдеров у благородных оленей и т. д. клептогамных самцов-«неудачников» (стратегия, альтернативная к таковой «центральных» самцов на току, территориальных рогачей у благородных оленей и пр.).
Недавно общественность возбудилась по поводу подробного выбора у самок сверчков, хотя он — общее место у многажды более близких к нам существ (включая предпочтение самцов с большей отзывчивостью на ответные реакции самки, более способных подстраиваться к её ответам сравнительно с более мотивированными, активными, больше демонстрирующими и пр., см. данные Гейл Патричелли по фиолетовому шалашнику, 1 и 2).
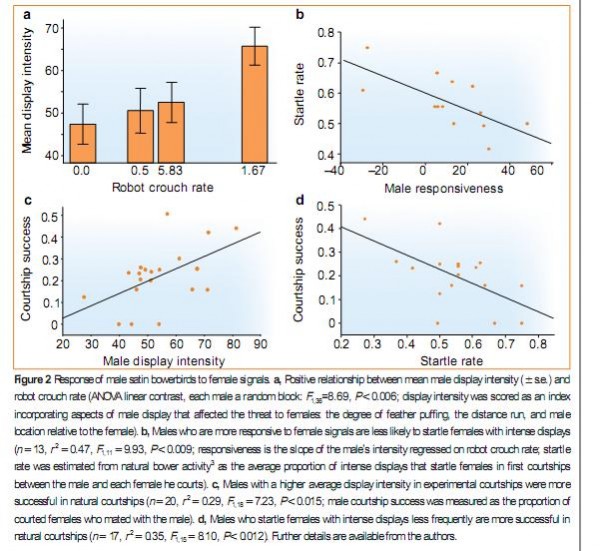 И эта особенность самок значимо ограничивает сверху способность полового отбора «двигать вперёд» признаки, важные для победы в конкуренции самцов, будь то агрессия у грызунов или длина хвоста у ткачиков р.Euplectes, превращает его в стабилизирующий, «удерживающий» адаптивный компромисс между интересами обоих полов (частью чего является вышесказанное про равную раскладку затрат)
И эта особенность самок значимо ограничивает сверху способность полового отбора «двигать вперёд» признаки, важные для победы в конкуренции самцов, будь то агрессия у грызунов или длина хвоста у ткачиков р.Euplectes, превращает его в стабилизирующий, «удерживающий» адаптивный компромисс между интересами обоих полов (частью чего является вышесказанное про равную раскладку затрат)
6. Рассматривая ряд близких видов с прямо противоположными взаимоотношениями партнёров: от постоянных пар с отцовской заботой о потомстве, до встреч партнёров только для спаривания с заботой одной лишь самки, через промежуточное состояние где у разных самцов и самок степень долговременности отношений, отцовской заботы и постоянства связи сильно варьирует индивидуально, мы видим что у них практически общая экология, как у полёвок р.Microtus и хомячков Peromyscus. На одном полюсе нет много больших затрат самок на репродукцию, которая бы требовала помощи самцов, при равенства этих затрат в парах. На другом полюсе самцы не спариваются сильно больше в сравнении с первыми, т.к. те много заботятся о потомстве, но отнюдь не хранят верность партнёру (или не всегда хранят, невязка генетической и социальной моногамии у птиц и млекопитающих встречается на каждом шагу). Такого рода различия не объяснить ни бейтмановскими градиентами, ни манипуляцией родительским складом.
7. Вопреки предсказаниям правила Бейтмана, большая конкурентность самцов, их высокий ранг и посредник того и другого – тестостерон – часто связан с большей родительской заботой и более устойчивыми отношениями с партнёром, а не с неверностью и максимизацией спариваний, см. 1-2. Данный эффект отмечен не только у моногамных видов с постоянными парами, но и у видов с изменчивой системой отношений, вроде обыкновенной полёвки Microtus arvalis: там есть полный диапазон вариантов,от соединения полов только для спаривания, после чего самка гонит самца и заботится о потомстве сама, до устойчивых пар и «заботливых отцов». О том же самом говорят данные М.Л.Бутовской по охотникам-собирателям хадза, где лидеры по телосложению и гормональному профилю «более феминны», и больше времени проводят с детьми.
Что ещё хуже для асимметричной модели с «правилом Бейтмана», «теневая сторона» социальности делается «лицевой» через ряд промежуточных этапов (этим поддерживается внутривидовой социальный полиморфизм) в зависимости от экологических обстоятельств, что подконтрольно естественному, а не половому отбору. Скажем, у на первый взгляд моногамных пёстрых дятлов и многих видов певчих воробьиных (включая «парадные примеры» асимметричности полового отбора), реальная система взаимоотношений полов в популяции — полигинандрия, где оба пола активно ищут спариваний и в паре и вне её, а тип отношений в каждой отдельной группировке зависит от местных условий. См. презентацию к докладу.

Токующие травники, самец и самка
9. Хороший пример, что развитие научных направлений означает не только приращение знание, но и забывание части «добытого» — симметричная теория полового отбора Джулиана Хаксли, см. его статьи о поведении чомги Podiceps cristatus 1914, краснозобой гагары Gavia stellata 1923, травника Tringa totanus 1916 гг. Сегодня (увы!) господствует асимметричная модель Бейтмана — Трайверса, с самцами, конкурирующими друг с другом в стремлении к максимальному числу спариваний, и самками, пассивно выбирающими по плодам такой конкуренции — «лучших» по генам и/или родительской заботе, и их этой «войн полов», мол, складывается всё наличное разнообразие социальных систем, в зависимости от количественного выражения этой конкурентности и этого выбора.

(Она же приписывает себе в предшественники самого Дарвина. Великого человека данные гг. используют как свадебного генерала: сам он не исследовал поведение животных в сообществах, лишь собирал рассказы, «джентльменов, заслуживающих уважения», всё это было в донаучный период поведенческих исследований. И часто упоминаемое «Выражение эмоций у человека и животных» это собрание мнений, часто с прозрениями, но не фактов.
Асимметричная модель Бейтмана-Трайверса, правда, опровергается фактами, давно собранными, известными всем интересующимся проблемой… однако ж «держится на плаву», не отвергаться научным сообществом в силу вненаучных причин, связанных с соответствием идеологии современного общества. Что верно и для других обобщений социобиологов; проверку эмпирией выдержал лишь принцип гандикапа. Здесь важно то, что с начала ХХ века до подъёма социобиологии в 1970-х господствовала другая модель, созданная «дедушкой этологии» Джулианом Хаксли в начале ХХ в..
Она, наоборот, симметрична. На стадии тестирования потенциальных партнёров оба пола «вкладываются» в оценку ухаживательных демонстраций и брачных украшений друг друга, но выбирают не самых интенсивно демонстрирующих, или с самыми длинными хохлами и воротниками, а «чуть длинней», чем у себя. Т.е. выбор здесь направляется не максимизацией, а оптимизацией по двум критериям, первый, как в асимметричной модели, развитость демонстраций (большая ритуализация) и развитость украшений (длина и пр. внешний вид), а второй — взаимное соответствие обоих партнёров, как в плане поведения, так и в плане мотивации (то же мы видим в выборе у грызунов, копытных и много ещё у кого).
Зачем нужно выбирать не «самого лучшего», а «сходного, чуть получше себя»? Тестирование потенциальных партнёров после нескольких отвержений заканчивается образованием пары, и с т.з. асимметричной модели, этого достаточно — самец должен трахаться и бежать к следующей, а самка — сразу заботиться о потомстве, включая более или менее успешные попытки его удержать. Так чёрта с два!
У моногамных видов дальше следует довольно длительный период ухаживания без копуляций (до 3 недель-месяца, см. как у большого пёстрого дятла Dendrocopos major), у других групп тоже довольно долго, см. статью Карла-Адама Вахтмайстера (2001). В ходе него птицы регулярно токуют, всё больше сближаясь со всё более эффективными демонстрациями, в результате чего риск срыва следующих токований страхом или агрессией (резко усиливающихся при слишком близком сближении, почему копуляция реализуема отнюдь не сразу, да и потом она осуществляется очень неточно, и эти ухаживания важны именно как способ повышения точности соединения клоак при спаривании).
Одновременно растёт устойчивость пары как нечувствительность обоих партнёров к ухаживанию холостых третьих птиц, — и только когда оба этих процесса дойдут до максимума, идут спаривания, завершающиеся яйцекладкой. А вот если погодные или биотопические условия будут плохими настолько, что это «сближение с демонстрированием» придётся таки сокращать, или хотя бы лишь демонстрирование, дело плохо. Пара не распадётся, её сочлены не сбегут в поисках нового, лучшего места для размножения, выстроят гнездо, но отложат туда 1-2 яйца, или не отложат вовсе. Так часто бывает у тех пар большого или малого пёстрого дятла, кто зимовал далеко от гнездовых биотопов, прибыл туда поздно, в последнюю неделю апреля, образовывал пары «в сокращённом режиме», и времени на ухаживание в образовавшейся паре просто не было.
Из: C-A.Wachtmeister (2001)
Всё происходящее радикально несовместимо с асимметричной моделью полового отбора, в рамках которой
а) непонятен разрыв во времени между образованием пары и «действенными» копуляциями,
б) необъяснима функциональная роль регулярных и интенсивных ухаживаний в образовавшихся парах, обычно сопровождаемых специальными ритуалами, обычно более заметными и выразительными, чем при тестировании потенциальных партнёров. Это разного рода дуэты у попугаев и кустарниковых сорокопутов, «поза пингвина» у чомг, аналогичные действия у гагар, триумфальная церемония у гусей и ряда уток и пр., см.табл.1 выше. Пара-то уже образовалась, и с т.з. асимметричной модели надо не выстраивать отношения, а сразу начать размножаться, но нет…
К.-А.Вахтмайстер пробует объяснить несовместность а-б с социобиологическими построениями о «войне полов», и в результате закономерно приходит к симметричной модели, похожей на Хакслианскую, о чём см. ниже;
в) репродуктивный потенциал самцов и самок определяющий приспособленность есть не атомарная, а реляционная характеристика: это не константа, увеличиваемая числом спариваний у первых и вложениями в заботу об их последствиях у вторых, но переменная, зависящая от степени реализации; а эта последняя определяется вложениями обоих во взаимное согласование поведения как раз на втором этапе ухаживания.
Нехватка социальной стимуляции ввиду дефектности коммуникации на этом этапе снижает успех размножения до нуля, что есть частный случай известного эффекта Олли.
г) Поэтому симметричная модель предполагает, что у таких видов оба пола, а не только самцы, будут предпочитать удлинённые брачные украшения, и отвергать укороченные. Да, это было показано на большой конюге — моногамном виде, с развитыми брачными украшениями (хохол перед клювом + косица, см.картинки), и при этом мономорфного, как чомги или гагары, с которыми работал Хаксли.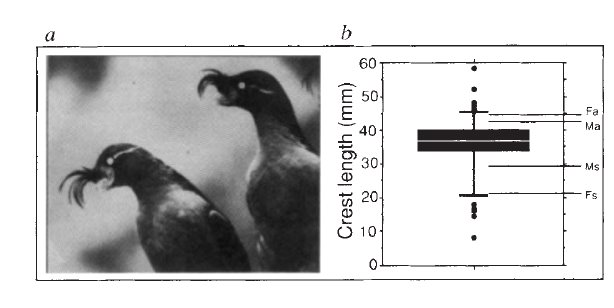 Исследования Яна Н. Джонса и Фионы Л. Хантер (1993, 1998, 2000) показывают что
Исследования Яна Н. Джонса и Фионы Л. Хантер (1993, 1998, 2000) показывают что
а) изменчивость украшений значимо больше, чем других деталей внешней морфологии, относящихся к линейным размерам, и связана с дифференциальной размножаемостью, а не выживаемостью птиц, т.е. если их контролирует какой отбор, но не естественный, а именно половой (привет отрицающему его Е.Н.Панову);

Из: Jones, Hunter, Fraser (2000)
б) искусственно удлиняя и укорачивая украшения, они получили избегание укороченных и предпочтение удлинённых у обоих полов.
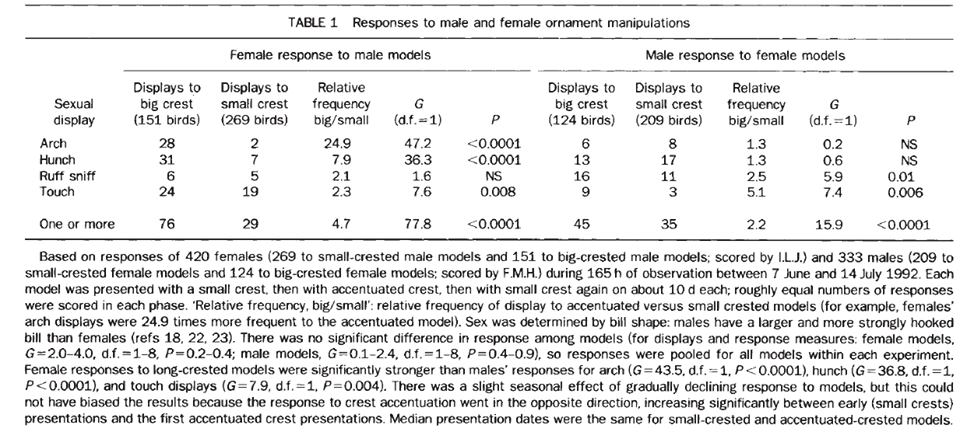
Из: Jones, Hunter, 1993.
Это — красивое подтверждение симметричной модели полового отбора Хаксли, согласно которой оба пола «вкладываются» в устойчивость отношений в паре, как к токованию «чужих», так и ко всяким средовым траблам, чем оно выше, тем полней реализуется репродуктивный потенциал обоих. Поэтому выгодно подбирать не лучшего, а сходного партнёра, чтобы твоё поведение было средством реализации чужой приспособленности ровно в той степени, в какой чужое поведение — твоей.
По ходу обмена брачными демонстрациями при постепенном сближении особи здесь конкурируют за то чтобы быть ведущим, а не ведомым в следующих актах коммуникации, чтобы хоть на немного, но эффективней использовать другого для своих нужд, чем он тебя (и брачные демонстрации выработались как способ «снять» противоречия, следующие из этой «войны полов» к выгоде для вида в целом — чтобы максимизировался репродуктивный выход всей популяции). Впрочем, последнее — мой сильно осовремененный пересказ, с учётом достижений социобиологов в области метода поведенческих исследований , не теории; сам Хаксли об этом не думал, что не делает его теорию хуже.
Можно спросить, а как же немоногамные виды, где пары временные и распадаются сразу после спаривания, вроде тетеревов, манакинов, диких индеек; их самцы собираются на токах, водят гаремы и не заботятся о потомстве.
Самки здесь получают дополнительную социальную стимуляцию, спариваясь с несколькими самцами, в т.ч. с субдоминантами, на разных токах, в виду господствующей там непрерывной череды поз и криков. А вклад самцов в заботу о потомстве состоит не столько в «хороших генах», сколько в обеспечении привлечённых током самок хорошей территорией для безопасного сбора корма вокруг места тока. Дело в том, что самцы на току исключительно уязвимы, тем не менее они демонстрируют часами, рискуя жизнью не меньше, чем радист разведгруппы в районе активности функабвера; коль они выбрали именно это место а не иное, значит местообитания вокруг тока просто превосходны в плане минимизации времени для кормёжки и безопасности. Принцип гандикапа Захави в действии — и равенство приспособленностей «лучших» и «худших» особей, при некотором проигрыше «средних», как менее специализированных в сторону той или другой стратегий!
Т.е. мы видим, что симметричная модель полового отбора Хаксли объясняет всю сумму относящихся к делу разнородных данных в разы лучше, чем асимметричная. Не зря именно с Хаксли полемизировал Ангус Бейтман в своей знаменитой статье 1948 г., где он или неверно обработал данные, или подогнал их под заранее заготовленный — т.е. идеологический — вывод. Он, собственно, поведением не занимался, кроме этих опытов для проверки важного для него общего принципа, и следующих, в 1949 г., для обоснования идеи поведенческих «препятствий к скрещиванию», также поставленной сейчас под сомнение.
Тем не менее, после преобладания в 30-60-х гг. (см. статью Хаксли 1938 г.) симметричная модель была прочно забыта, притом что Трайверс и другие фундаторы асимметричной модели (и социобиологии) все приведённые работы Хаксли честно цитировали. И приведённые данные в пользу неё и против конкурирующей пока что не помогли её «вспомнить»: такова сила идеологического компонента, поддерживающая социобиологов и других биологических редукционистов вопреки эмпирическим данным((( Притом что другие достижения Хаксли в области этологии — теорию ритуализации, например, идею двух форм выражения побуждений — ритуализированного в демонстрациях и в прямых действиях — зоологи помнят и всяко используют.



![Элементы брачного поведения красношейной поганки (по [Cramp, Simmons, 1977], с изменениями)](http://www.socialcompas.com/wp-content/uploads/2019/12/31.gif)