

В продолжение темы современных представлений об инстинкте
С.П. Харитонов
Резюме. Основная задача данного сообщения – описать выделенные, или классифицированные, автором три стратегии принятия птицами решения и том, гнездиться в данном месте или нет. На основании опыта работы с различными видами околоводных птиц — в 1989–1992 гг. с чайковыми птицами Восточного Сиваша, Херсонской области, Украина; с белыми совами (Nyctea scandiaca) и гусеобразными птицами тундры в 2000–2018 гг. в окрестностях бухты Медуза (73°21′ с.ш., 80°32′ в.д.), северо-запад п-ова Таймыр; с озёрными чайками (Larus ridibundus) в 2010–2018 гг. на оз. Киёво в Московской области — обсуждается вопрос о механизмах принятия решения о месте расположения гнёзд у птиц.
Выявлены три стратегии поведения при принятии решения о том, гнездиться или не гнездиться в данной местности. Первая — рационально-вероятностная. Такая стратегия обнаружена у 4-х видов гусей, они оказались способны оценивать численность песцов на разных участках тундры, а чёрные казарки могут оценивать ещё и численность леммингов. Вторая стратегия — ориентации преимущественно на сиюминутные раздражители — состоит в том, что при выборе места гнездования превалирует реакция на небольшой набор внешних сиюминутных раздражителей (релизеров), что отмечено у ряда видов чайковых Приазовья.
Третья — стратегия следования консервативным поведенческим установкам («традициям») — оказалась присуща озёрным чайкам в условиях длительного существования постоянных водоёмов. Нередко птицы следуют традициям, даже если рационально-вероятностная стратегия представляется более выгодной. Степень следования трём вариантам стратегии различается у разных видов и даже разных популяций одного вида. Показано, что при помощи целенаправленного искусственного применения небольшого числа внешних раздражителей можно поменять поведенческие установки птиц, перенастроить выбор места гнездования, то есть искусственно «включить вторую стратегию». Обычных природных раздражителей для этого может быть недостаточно, приведён пример использования супер-раздражителей.

Введение
Содержание
Согласно современному подходу к изучению поведения животных — когнитивной этологии — у них есть способность к размышлению, оценке ситуации и выбору наиболее верной/выгодной стратегии поведения (Резникова, 2005). В европейской концепции, основанной К. Лоренцом и Н. Тинбергеном, считается, что поведением управляют одномодальные (однофакторные) раздражители (Дьюсбери, 1981). К. Лоренцом разработана так называемая гидравлическая модель поведения животных. В этой модели «бачок» (организм животного) постоянно наполняется «водой» (энергия). Существует «клапан», который может открываться только специфическими ключевыми раздражителями.
При воздействии этих раздражителей срабатывает врождённый спусковой механизм. По мере накопления энергии животное проявляет аппетентное поведение, т. е. начинает искать ключевой раздражитель. Если раздражитель «откроет клапан», «вода выливается», питая энергией соответствующее врождённое поведение: это консуматорный, или завершающий, акт (Дьюсбери, 1981). Чем больше энергии накопилось и чем больше времени животное затратило на аппетентное поведение, тем более слабое воздействие ключевого раздражителя запускает завершающий акт. Хороший тому пример — при долгом отсутствии самки самец каменки может начать «спариваться» с камнем (Панов, 1978).
Если в европейской концепции считается, что животным управляют однофакторные раздражители, то согласно американской концепции животные руководствуются рационально-вероятностной стратегией поведения (25.10.2010 г., Peter Moller, The City University of New-York, обзорный доклад на Этологическом семинаре ИПЭЭ РАН – описание стратегии приведено ниже). Если ссылаться на описанную выше гидравлическую модель, то по представлению современной американской школы поведения в животном действуют не один, а много таких гипотетических «бачков».
Для иллюстрации рационально-вероятностной стратегии поведения опишем приведённый Питером Моллером эксперимент. Пусть в пруду плавает 30 уток. На одной стороне пруда человек каждые 10 секунд кидает кусок хлеба весом 5 г, а на другом — 5 г каждые 20 сек. Утки распределяются в соотношении два к одному: 20 уток будут там, где хлеб кидают раз в 10 сек, а 10 уток — там, где раз в 20 сек. Утки распределились по пруду так, чтобы каждая могла получать примерно одинаковое и максимальное количество пищи. В этом эксперименте предсказать поведение каждой утки невозможно (неизвестно, какая именно птица поплывет к первому или второму экспериментатору), но в сумме получается полностью предсказуемый результат: утки распределяются именно в таком соотношении, которое указано выше.
Животные обычно не имеют полной информации об окружающем их мире, но должны проводить оценку условий среды и принимать решения не только на текущий момент, но и на будущее. Рационально-вероятностное поведение как для индивидов, так и для коллективов, хорошо описывается теорией игр — от детерминистской к вероятностным (probabilistic) моделям, через представление о «рационально-вероятностном решателе» (Kamamba et al., 2007a, 2007b).
Когда мы используем вероятностные модели, то разрыв между простым и сложным поведением исчезает. Собственно, это и называется стратегией поведения. Кроме рационально-вероятностной, мы выделяем еще две стратегии поведения: стратегия ориентации преимущественно на сиюминутные раздражители и стратегия следования консервативным поведенческим установкам («традициям»).
Методика
Работу проводили на протяжении многих лет с различными видами околоводных птиц. В 1989–1992 гг. с чайковыми птицами Восточного Сиваша, Херсонская обл., Украина; в 2000–2018 гг. с белыми совами (Nyctea scandiaca) и гусеобразными птицами в тундрах в окрестностях бухты Медуза (73°21′ с.ш., 80°32′ в.д.), северо-апад п-ова Таймыр; в 2010–2017 гг. с озёрными чайками (Larus ridibundus) на оз. Киёво, Московская обл. На Восточном Сиваше осуществляли наблюдения за формированием колоний морских голубков (L. genei), а также малых (Sterna albifrons), речных (S. hirundo) и пестроносых (Thalasseus sandvicensis) крачек. На п-ове Таймыр наблюдали за формированием гнездовых поселений гусеобразных и белых сов и картировали места расположения гнёзд этих видов при помощи GPS-навигатора. Кроме того, удалось провести два эксперимента (один из которых длился четыре года, второй — один гнездовой сезон) по изучению реакции белых сов и чёрных казарок (Branta bernicla) на макеты, изображающие белых сов, заблаговременно выставленные в тундре. На оз. Киёво в течение пяти лет проведена серия экспериментов с различными зрительными и акустическими раздражителями с целью восстановления колонии озёрных чаек, которая прекратила своё существование на этом озере за 15 лет до начала наших экспериментов.
Результаты
Поведение гусей в тундре и принятие ими решения о способе гнездования
Абиотические условия тундр (погода, уровень снежного покрова и проч.) переменчивы, но в целом относительно хорошо предсказуемы (предсказуемы пределы изменчивости температуры окружающей среды, по многолетним периодам предсказуемы предельные уровни снега и т.д.). Биотическая среда, в частности — уровень хищничества главного для птиц хищника, песца, — гораздо менее предсказуема. В серии работ было показано, что белолобые гуси (Anser albifrons), чёрные и краснозобые казарки (Branta ruficollis) могут оценивать численность песца в каждый конкретный сезон, а чёрные казарки могут оценивать и численность леммингов (Сыроечковский, 1999; Харитонов и др., 2008, 2013).
Последнее им необходимо, чтобы выбрать верную дистанцию от гнезда белой совы для постройки своего гнезда: необходимо загнездиться достаточно близко, чтобы находиться в защитной зоне вокруг гнезда белой совы (самцы этого вида хорошо защищают свою территорию от песцов) и, в то же время, достаточно далеко, чтобы не пострадала кладка и не быть съеденными самим при нападении самки совы, если численность леммингов окажется недостаточной для питания совиного выводка. Нами было показано, что чёрные казарки располагают свои гнёзда вокруг гнезда совы вполне рационально. Среднее расстояние от гнезда совы до гнёзд чёрных казарок было связано с численностью леммингов и песцов в районе гнездования (Харитонов и др., 2008, 2013). [Сильно ранее было показано, что в зависимости от обилия грызунов сходным образом усиливается/ослабляется отводящее поведение самок куриных птиц, в т.ч. по степени ритуализации. Прим.публикатора]
В этом случае мы наблюдаем такую стратегию при выборе гнездового места, которую можем рассматривать как рационально-вероятностную. Однако, часть чёрных казарок поступали иначе, ориентируясь только на небольшой набор зрительных раздражителей. Именно этой причиной мы можем объяснить возникновение поселений казарок из 1–4 гнёзд как возле гнёзд зимняков (Buteo lagopus), которые являются плохими защитниками от песца, так и вокруг токовой точки самца тундряной куропатки (Lagopus mutus), видимо напоминающего своей окраской самца белой совы, и даже около белого пенопластового ящика из-под метеозонда (Харитонов и др., 2008). Выбор места для гнездования с опорой только на небольшое число внешних, вероятно ключевых раздражителей (релизеров) оказывается инадаптивным — все такие гнезда были разорены песцами.

Гусак чёрной казарки (Branta bernicla nigricans) защищает гнездо. Остров Врангеля, среднее течение реки Тундровая, начало июля 1970 г.
Дополнительный материал для оценки роли релизеров дали наши полевые эксперименты. Роль релизеров играли макеты белых сов, предварительно выставленные в тундре в 2012 г. С их помощью удалось повлиять на выбор места гнездования двух пар белых сов в 2016 г. и вызвать реакцию пары чёрных казарок, выбиравших место для устройства гнезда. Макеты белых сов были изготовлены из отшлифованных морем деревянных болванок, покрашенных в белый цвет с нанесёнными чёрной краской изображениями глаз и клюва.
Макеты не были точной копией белых сов, поскольку при их изготовлении мы руководствовались наблюдениями одного из основателей этологии Н. Тинбергена. Он выяснил, что важным является не точное соответствие модели общему виду особи, а соответствие макета по цвету или форме некоторым частям её тела, которые, будучи утрированными, играют роль ключевых раздражителей (Тинберген, 2012). В его экспериментах самец зарянки (Erithacus rubecula) бросался в драку c «соперником», роль которого играл кусок ваты, окрашенный в оранжевый цвет, как грудь самой зарянки. А самцы гуппи (Poecilia reticulata) с красной грудью реагировали на проходящие за окном трамваи красного цвета (Тинберген, 2012).
Для того чтобы понять ход и результат нашего эксперимента, необходимо остановиться на особенностях гнездования белых сов на Таймыре. Они гнездятся при высокой численности леммингов, вроде бы случайно распределяясь по тундре. Однако, нам удалось выявить так называемые территориальные ячейки белых сов: участки без чётких границ, поскольку у охраняемых территорий птиц обычно четких границ не бывает, на которых в разные года гнездятся белые совы. Если они решили загнездиться в данном районе тундры в конкретном году, то это будет не случайно выбранное место, а одна из указанных территориальных ячеек (Харитонов и др., 2005). При этом непонятно, чем руководствуются белые совы, каждый раз «попадая» в одни и те же ячейки, хотя между двумя последовательными гнездованиями на них может пройти более 10 лет, и пары гнездящихся здесь в разные годы сов очевидно разные. Место расположения прошлого гнезда в этих случаях не играет роли: совы гнездятся либо на плоских камнях, после чего от гнезда не остаётся ни малейшего следа, либо, если гнездо располагалось на траве, то птенцы так утаптывают это место, что оно на многие годы становится непригодным для устройства следующего гнезда.
В ходе четырёхлетнего эксперимента на одной из таких территориальных ячеек был поставлен макет «вертикально сидящий самец», а на другой — макет насиживающей самки; вокруг неё на расстоянии от 8 до 150 м были размещены ещё 5 макетов самцов. В 2016 г. численность леммингов была низкой; при такой численности этих грызунов нам никогда не удавалось наблюдать гнездование белых сов.
Тем не менее, одна пара сов загнездилась непосредственно в том кластере, где стоял макет самца (правда, так, что гнездо со стороны макета было отгорожено небольшим пригорком, что препятствовало прямой видимости макета от гнезда). Вторая пара сов загнездилась в районе той ячейки, где были выставлены макеты самки и пяти самцов. Очевидно, пара не могла загнездиться ни в той же самой ячейке (слишком велика виртуальная конкуренция с «целыми пятью самцами»), ни совсем рядом — при малой численности леммингов белые совы рассредотачиваются по территории. В результате, вторая пара загнездилась «через ячейку» от той, в которой были выставлены 6 макетов, то есть вторая пара «ушла подальше» от ячейки, «перегруженной» зрительными раздражителями. Если использовать ту же терминологию, то можно считать, что первая пара сов оказалась ими «недогружена»: она загнездилась в той же ячейке, где был выставлен «одиночный самец» (рис. 1). Ни в каких других ячейках данного района белые совы в 2016 г. не гнездились.
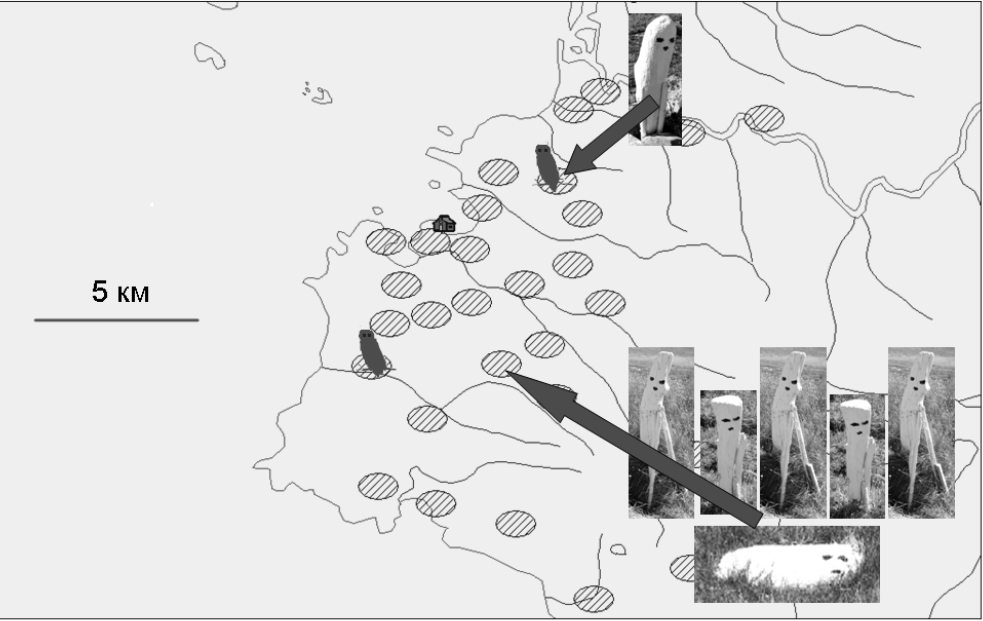
Рис. 1. Расположение территориальных ячеек, в которых загнездились белые совы в 2016 г.: заштрихованные овалы – многолетние территориальные ячейки белых сов (объяснение в тексте), фото – выставленные в двух ячейках макеты (ячейки указаны стрелками). Силуэты сов – места гнездования двух пар белых сов.
Наблюдения и ещё один эксперимент, проведённые в 2018 г., показали, что белые совы действительно реагируют на используемые нами довольно грубые макеты особей своего вида. 7.06.2018 г. на плакоре в тундре была установлено два новых макета, имитирующих насиживающую самку и стоящего в 50 см от неё самца. Уже на следующий день в 120–150 м от макетов мы обнаружили самца белой совы, который, вопреки ожиданию, при вспугивании не покидал окрестностей макетов, а лишь перелетал с места на место. Всего за следующие 7 дней в ближайших окрестностях макетов были отмечены три белых совы: два самца и одна самка. Хотя птицы не были помечены, самцы легко опознавались по цвету и характеру расположения незначительной пятнистости на оперении. Самец, который появился у макетов первым, был почти белым, с очень малым количеством пятен.
Второй самец имел на спине строчки пятен рыжеватого цвета, что, скорее всего, свидетельствует о его более молодом возрасте. Самку наблюдали лишь однажды, 9.06. Она приблизилась к макетам на расстояние менее 100 м и провела там более 2 часов. Ни один из двух самцов за время наших наблюдений так близко к макетам не приближался.
Первый самец провел около макетов 7 дней, перемещаясь вокруг них практически по кругу на расстоянии 100–800 м. Второй самец отмечался 4 дня и перемещался по полукругу на расстоянии 800–1500 м от макетов, то есть явно ориентировался на положение первого самца.
Вечером на седьмой день присутствия на обсуждаемом участке первого самца макет упал из-за подтаивания почвы, и на следующее утро первый самец исчез из района наблюдений (второй сделал это раньше). Ещё один пример очевидной реакции совы на макет зарегистрирован 15.07.2018 г., когда в непосредственной близости одного из макетов самца, на которого, по нашему мнению, была отмечена реакция гнездящейся пары сов в 2016 г., найдены более десятка перьев, принадлежавших белой сове, в том числе два маховых.
Находка свидетельствует о том, что птица провела здесь не один час. При этом не только на окружающих холмиках не было найдено перьев белой совы, но и за все годы наблюдений не было случая, чтобы удалось видеть столь большое количество перьев совы, свидетельствующее о долгом нахождение птицы на одном месте.
Описанные эксперименты и наблюдения убедительно показали, что самцы белых сов, которые, собственно, и «держат» территории, при заселении какого-либо участка тундры вполне могут ориентироваться друг на друга и выбирать положение будущего гнезда в зависимости от распределения соседних особей. Убедительность этих выводов усиливается ещё и тем, что численность леммингов в 2018 г. в районе работ была низкой, и белые совы не только не гнездились здесь, но и помимо встреч рядом с макетами отмечались лишь трижды и не задерживались в местах встреч.
Белые совы могут ориентироваться на положение других особей и без прямых видимых поведенческих взаимодействий. Проявлялось это в том, что каждая из трех наблюдаемых около макетов сов держалась только в определенном области по отношению к макетам: один самец перемещался в кольцевой области в непосредственной близости от макетов, второй самец на этот район ни разу не претендовал, хотя стремился к макетам, тоже перемещался по кольцевой области вокруг них, но – только за пределами области первого самца. Никаких видимых взаимодействий между самцами не отмечено. Самка была ближе всего к макетам, в той области, куда при ее присутствии не заходил даже первый самец. Взаимодействий тоже не отмечено.
Кроме реакции белых сов на макеты, в 2016 г. удалось наблюдать и сфотографировать реакцию пары чёрных казарок на макет насиживающей самки белой совы. Пара казарок держалась в этом районе более недели и демонстрировала намерение построить гнездо — примерно в 10 м от макета насиживающей самки совы (рис. 2). Именно так чёрные казарки поступают, формируя свои колонии: большинство их гнёзд располагается вокруг насиживающей самки белой совы.

Рис. 2. Закладка места гнезда парой чёрных казарок в 10 м от макета насиживающей самки белой совы. Качество снимка связано с дальностью съёмки (более 800 м)
Почему всё же пара чёрных казарок не загнездилась около макета? Вероятное объяснение этому — возможный механизм оценки казарками численности леммингов. Если численность песцов, очевидно, можно оценить за счёт прямых наблюдений, то численность леммингов чёрные казарки, по нашим представлениям, могут оценить только по активности самца белой совы: если часто подносит самке леммингов, то, значит, их много, можно гнездиться около этого гнезда совы. Если самец редко приносит леммингов, значит их мало, гнездиться рядом с гнездом совы опасно, можно пострадать. Поскольку макеты сов леммингов не приносили, казарки, скорее всего, решили, что численность леммингов столь мала, что гнездиться около данной пары сов опасно.
Отметим, что возле одной из двух упомянутых пар сов другая пара чёрных казарок в 2016 г. всё-таки загнездилась. Однако, загнездилась она на расстоянии 700 м от гнезда совы, что почти совпадает с граничным положением (680 м), на котором можно считать казарок ассоциированными с белыми совами (Харитонов и др., 2008). Получается, что указанная пара казарок загнездилась в ассоциации с белой совой, но при этом максимально ей «не доверяла». Краснозобые казарки при той же численности леммингов гнездятся ближе к белым совам, чем чёрные. Например, в том же 2016 г. две пары краснозобых казарок устроили гнёзда в 123 и 251 м от гнёзд сов. В другие годы краснозобые казарки также гнездились к совам в среднем ближе, чем чёрные. Причина может лежать в неверной оценке этими птицами опасности, исходящей от совы при малой численности леммингов, или же — в большей подвижности и большей агрессивности краснозобых казарок по сравнению с чёрными, что даёт им возможность лучше защищать своё гнездо при возможном нападении белой совы.

Итак, удалось выявить по крайней мере два фактора, на которые опирается рационально-вероятностное поведение гусей при выборе места гнездования. Механизм действия первого фактора можно считать понятным: это непосредственная оценка казарками численности песцов в конкретном районе. Второй фактор, численность леммингов, предположительно оценивается через наблюдение чёрными казарками за кормовым поведением сов. Оба этих фактора, по-видимому, служат для казарок сигналом пригодности данной гнездовой территории совы для гнездования в конкретном сезоне.
Выбор места гнездования чайковыми птицами в условиях нестабильных биотопов
В этих условиях наблюдается примат реакции на кратковременно существующие раздражители.
Обширные мелководные южные водоёмы (например, Сиваш) — очень нестабильная среда, где существует постоянная угроза затопления мест гнездования при нагонных ветрах. В этих условиях большое значение при выборе места гнездования играет фактор случайности и наличие визуальных раздражителей (куртины и пучки травы, гнездящиеся конспецифичные особи и т.п.). Если затопление гнездовий всё же происходит, ряд видов птиц «обучается» и начинает поступать более рационально: морские голубки и пестроносые крачки в течение данного сезона гнездятся повторно уже только на локальных возвышенностях.
Малые и речные крачки продолжают поступать по-прежнему, т.е. вновь строят гнёзда в потенциально затопляемых местах (Kharitonov, 1999). Не меняют места размножения после затопления гнёзд и чёрные водорезы (Rynchops niger). В Техасе эти птицы гнездятся на островах у самого уреза воды, и их гнёзда смывает штормом, если он бывает в гнездовое время. Через две недели после смыва водорезы гнездятся повторно на точно тех же местах и в том же количестве (White et al., 1984).
Условия среды могут выглядеть для вида весьма привлекательными, однако на самом деле являются неподходящими. Показательный пример из этой области мы наблюдали также на Восточном Сиваше. Малая крачка там гнездится повсеместно, одиночными парами и колониями. Наиболее предпочитаемое место гнездования для этих птиц — это юго-западный берег острова, на котором мы проводили работу; здесь достигается наибольшая плотность гнездования этого вида. При этом колония всегда смывается нагонной водой, и, тем не менее, малые крачки в наибольшей степени стремятся именно сюда. В данной популяции больше половины особей (судя по ряду признаков, — наиболее «качественных» особей) из года в год упорно гнездятся на месте, где успех размножения равен нулю из-за постоянно возникающих нагонных ветров, неизменно разрушающих колонию (Kharitonov, 1999).
Подобным образом малая крачка ведёт себя и в Австралии, а близкий к ней вид — американская малая крачка (Sterna antillarium) — в Северной Америке. Эти птицы гнездятся там, в частности, на плоских соляных равнинах, где воде стекать некуда, поэтому малейший дождь вызывает массовую гибель гнёзд (Winton, Leslie, 2003). В Австралии малые крачки гнездятся нередко возле самого уреза воды, отчего яйца заливаются при первом же высоком приливе, причину подобного явления авторы удовлетворительно объяснить не могут (Egan, Webb, 2000). На одном небольшом островке у побережья Австралии на песчаном пляже гнездятся 5 видов крачек, при этом каждый высокий прилив вызывает гибель большинства их гнёзд (Walker, 1993).

Аналогичные массовые затопления с ликвидацией большей части гнёзд черноголовых чаек (Larus melanocephalus), пестроносых, речных и малых крачек отмечены в Венецианской лагуне (Scarton, Valde, 1997). У чаек-хохотуний (Larus cachinnans) на о. Огурчинском в Каспийском море также отмечено весьма странное, с точки зрения адаптивности, явление. Там часть колоний чаек-хохотуний образуются в прибойной полосе, затем эти колонии смываются штормами. При этом имеется избыток мест для гнездования в защищённых от затопления местах (Панов и др., 1990).
Стратегия выбора места гнездования на основе консервативных поведенческих установок
В условиях среды, постоянной на протяжении некоего длительного промежутка времени (десятилетия), у популяций птиц могут вырабатываться консервативные поведенческие установки («традиции») по выбору место гнездования. В последнем случае птицы следуют традициям даже в том случае, когда, казалось бы, руководствоваться рационально-вероятностной стратегией было бы лучше, как с точки зрения кормности окружающей территории, так и с точки зрения защищённости от хищников и от беспокойства со стороны человека. Консервативная стратегия предполагает отказ от многих преимуществ рационального поведения, как-то гнездование в более защищенных местах и в местах, расположенных близко к источникам обильного корма. Пример: озёрные чайки имеют поведенческую установку — стремление гнездиться в центре колонии, хотя там успех размножения меньше, чем на периферии. При массовом разорении гнёзд в центре колонии птицы на её периферию не возвращаются, а покидают данную колонию вовсе (Харитонов, 1993).
На постоянных водоёмах к северу от Москвы при резком уменьшении кормности окружающей колонию местности озёрные чайки не меняют место колонии на расположенное ближе к источникам корма, а продолжают летать за кормом на очень большие расстояния.
Очевидно, этим птицам важнее гнездиться пусть и в менее удобном, но знакомом месте. Озёрные чайки в этом районе Московской области имеют тенденцию использовать место, в котором они в этом году поселились, и в дальнейшем.
В других местах бывают разливы и временные затопления мест гнездования (например, в пойме р. Москвы). В этих условиях чайки частично придерживаются консервативных поведенческих установок — гнездятся в немногочисленных постоянных колониях поймы. Другая часть чаек ведёт себя как морские голубки и пестроносые крачки на Сиваше — начинают ориентироваться на небольшой набор внешних коротко существующих раздражителей: гнездятся в тех местах, где из-под разлива появляются островки, поначалу окружённые водой. Таким образом, проявляется слабо исследованное этологией явление: поведение животных в разных популяциях одного вида, обитающих в разных географических районах, может отличаться столь же сильно, как оно отличается у разных видов. Разливы в Виноградовской пойме р. Москвы были частыми в 1980-е гг., но сейчас они там редки, и озёрные чайки опять руководствуются консервативными поведенческими установками: гнездятся в двух постоянных колониях, причём в одной из них гнёзда расположены над водой на ветвях кустов. Строить такие гнезда чайкам куда сложнее, чем гнёзда на поверхности земли, посещать и покидать такие гнёзда тоже сложно, так как озёрной чайке — птице открытых пространств — приходится миновать при этом переплетение ветвей, тем не менее, традиция «не позволяет» им сменить место гнездования на более удобное. Такие места в пойме есть: например, место одной из постоянных в прошлом колоний, которые чайки покинули в засушливый период (1990-е гг.); сейчас там условия восстановились, но озёрные чайки туда не возвращаются, хотя следование рационально-вероятностной стратегии несомненно облегчило бы чайкам существование в случае возвращения к гнездованию в этом месте.
Поведение озёрных чаек и в данном случае указывает на то, что переключение на реализацию стратегии, предусматривающей ориентацию на небольшое число внешних коротко существующих раздражителей происходит легко: весной 2018 г. разлив был довольно высоким, и чайки «ушли» с кустов на расположенный рядом временный островок; на кустах загнездились немногие чайки. Такие островки после падения уровня воды соединяются с сушей, и колония становится доступна для наземных хищников. Зрительный раздражитель — «островок» — даёт чайкам неверную информацию о конкретных условиях среды, и реакция на этот раздражитель чаек часто подводит (Харитонов, 1998).
Консервативные поведенческие установки известны и для других групп животных. Например, у ушастых тюленей (Otariidae) наблюдается приверженность к одним и тем же репродуктивным пляжам. На этих пляжах тюлени страдают от переуплотнения, хотя рядом имеются совсем пустые пригодные места (Панов, 1973).

Спаривание пестроносых крачек
«Можно видеть, как вокруг этой пары толпятся сразу несколько других крачек. Они не только кричат, но еще и пытаются схватить самца клювом за перья. Перед спариванием партнеры сами невольно привлекают внимание окружения: они возбуждены, вокализируют и/или принимают заметные позы. Собственно спариванию предшествует садка самца, когда он запрыгивает на спину самки, удерживая равновесие крыльями. В этот момент он наиболее уязвим к действиям своих соседей. Слишком настойчивое внимание последних может заставить самца спрыгнуть с самки.
По моим наблюдениям, сделанным на Азовском море, значительная часть попыток спаривания может прерываться таким образом. Интересно, что в ряде случае партнеры специально отходят на несколько метров от гнезда, если там есть места, где плотность крачек поменьше. Но и это не помогает. Заметив известную активность партнеров, товарки специально подбегают к ним, преодолев до десятка метров. Так что наладить личные отношения в многотысячном коллективе оказывается не так просто. И это — лишь одно из проявлений дезорганизации, которые можно наблюдать у колониальных видов птиц».
Практическое управление выбором места гнездования озёрными чайками
Во время восстановления колонии озёрных чаек на оз. Киёво (Московская обл.) оказалось, что необходимо не просто привлечь чаек на озеро, но и «сломать» их традиции — выработавшуюся за много лет приверженность (термин наш, Харитонов и др., 2016) к другим колониям. В 1980-е гг., когда на оз. Киёво ещё существовала большая колония озёрных чаек, основными кормными местами служили две свалки бытового мусора: Лобненская в 4.5 км от колонии и Долгопрудненская — в 12.5 км от колонии. Интересно, что в эти годы всего в 3 км от Долгопрудненской свалки уже существовал водоём (близ станции «Марк» Савёловской железной дороги), куда более удобный для гнездования озёрных чаек, чем берег оз. Киёво. Кроме близко расположенного источника обильного корма, сплавины на том водоёме вполне выдерживали гнёзда озёрных чаек, но проваливались под тяжестью человека. В отличие от оз. Киёво, где сплавины человека держат, и потому люди время от времени беспокоили чаек. На водоёме у станции «Марк» люди и бродячие собаки чаек не беспокоили. Тем не менее, в стабильной ситуации озёрные чайки в течение ряда лет этот более удобный водоём не осваивали. Его заселение чайками началось только тогда, когда началось разрушение гнездового биотопа на оз. Киёво. Впервые чайки загнездились у станции «Марк» в 1984 г., на следующий год после первого (в 1983 г.) массового разорения гнёзд на оз. Киёво (Харитонов, 1993). Численность птиц в колонии озёрных чаек на водоёме у станции «Марк» за несколько лет (учет проведен только в 1996 г.) достигла 5000 пар (Зубакин, 1998).
В 2011 г., когда начались мероприятия по восстановлению колонии чаек на оз. Киёво, на сплавинах в качестве раздражителей были выставлены макеты озёрных чаек сходного с ними размера и проигрывались чаячьи крики. Примечательно, что в 2012 г. была закрыта Долгопрудненская свалка, и чайки с водоёма у станции «Марк» вынуждены были летать за кормом уже на Икшинскую свалку в 24 км от своей колонии. При этом их кормовые перелёты пролегали прямо над оз. Киёво. Однако, теперь «традиция» озёрных чаек уже поддерживала у них установку на гнездование у станции «Марк», оз. Киёво стало для них новым водоёмом. Даже такое преимущество, как возможность летать за кормом в два раза ближе, их «не прельстила», не помогали также выставленные на оз. Киёво макеты чаек и демонстрация голосов.
Для того, чтобы «пересилить» традицию озёрных чаек (приверженность теперь уже к колонии у станции «Марк»,) пришлось применить супер-раздражители в виде макетов чаек удвоенного по сравнению с реальным размера (Харитонов и др., 2016). Традиция менялась с большим трудом: численность чаек на восстанавливаемой колонии оз. Киёво растёт очень медленно: в 2013 г. здесь были построены 2 гнезда, в 2014 — 100; в 2015 — 600; в 2016 — 950; в 2017 — 1250; в 2018 г. — 1450 гнёзд.
Обсуждение
Стратегия — это сознательная или бессознательная система действий живых организмов по организации (организационному обеспечению) своего жизненного цикла. В реальной жизни у птиц отмечены, по крайней мере, три стратегии поведения при принятии решения о том, гнездиться или нет в данной точке: 1) рационально-вероятностная; 2) стратегия преимущественной реакции на раздражители (возможно ключевые): поведение при выборе места гнездования фактически управляется раздражителями; 3) консервативными поведенческими установками.
Рационально-вероятностная стратегия выглядит как просчитывание всех возможных плюсов и минусов своего поведения, при этом птицы учитывают не только природные условия, но во многих случаях могут оценить и свои возможности. Кроме описанного выше примера с оценкой численности песцов и леммингов у чёрных казарок, хорошим одним примером рационально-вероятностного поведения служит гнездование слабых особей в зоне повышенной опасности для данного вида. Самая слабая пара белых гусей (Anser caerulescens), на которую агрессивно реагировали все соседи, загнездилась в 2 м от палатки наблюдателя, то есть в том месте, куда остальные гуси боялись заходить (Харитонов, 1988). В обширной колонии тихоокеанских чёрных казарок (Branta bernicla nigricans) самка, потерявшая партнёра (а значит — и защиту от вторгающихся особей), устроила гнездо всего в нескольких метрах от наблюдательной вышки, к которой другие чёрные казарки боялись приближаться (наши наблюдения в дельте Юкона на Аляске в 1993 г.).
Виды, следующие рационально-вероятностной стратегии при выборе мест гнездования, могут переходить к использованию и двух других упомянутых выше стратегий. Такому переключению отчасти может способствовать, очевидно, не только конкретные условия среды, но и то, что две последние стратегии, видимо, легче в исполнении, чем рационально-вероятностная. Реакция на небольшое число внешних раздражителей, как уже было сказано, может лишать птиц возможности использовать, казалось бы, наилучший выбор места гнездования.
Консервативные поведенческие установки, на наш взгляд, также нередко оказываются инадаптивными: образно выражаясь, птицам не надо «думать», а нужно продолжать действовать так, «как жили предки». Причём все это происходит, несмотря на очевидные недостатки двух последних упомянутых стратегий. Иррациональное поведение для ситуаций, когда требуется сложная оценка условий — это «падкость на небольшое число внешних сиюминутных раздражителей». Те же черные казарки реагируют на отдельные поведенческие раздражители, отчего могут гнездиться в совсем неподходящих и даже опасных местах.
Особенность консервативных поведенческих установок в том, что птицы, следуя им, не считаются со значительными дополнительными физическими усилиями. Объём этих дополнительных усилий опровергает общепринятый взгляд на биологические объекты. Считается, что живые организмы стремятся к минимальным затратам энергии. Однако уже многократно показано, в том числе и в приведённом нами случае далёких полётов озёрных чаек за кормом, что это вовсе не так: живые организмы готовы проявлять избыточную активность, и у них может отсутствовать стремления к расходованию минимума энергии.
Видимо, все три обсуждаемые стратегии могут реализовываться в поведении любых птиц. Всё дело в степени выраженности той или иной стратегии. Вторая из них — реакция на сиюминутные раздражители — обладает той особенностью, что может «пересиливать» две другие стратегии и направлять поведение птиц в новое русло. Раздражители могут поменять поведенческие установки озёрных чаек, их использование даёт нам возможность повлиять на их решение гнездиться в новом для них месте. При этом оказалось, что обычных, встречающихся в природе раздражителей для «перенаправления» этих чаек на новое место гнездования может быть недостаточно; могут потребоваться т.н. супер-раздражители, которые в природе не встречаются (Харитонов и др., 2016).
Представляется, что раздражители, или релизеры, могут быть полезны для скорых решений, но для реального благополучия популяции нужны длительные решения, то есть — рационально-вероятностная стратегия (с одной стороны, про это уже сказано, с другой – в чём же тогда полезность скорых решений, если мы знаем, что они неполезны? – рец.). Хотя релизеры считаются явлением адаптивным (а что существующее не считается адаптивным?), в реальной жизни при выборе места гнездования они нередко подводят, «отвлекают» птиц от рационально-вероятностной и консервативной стратегий, «заманивают» в дефективные условия для размножения.
Все эти явления настоятельно требуют своего изучения. На наш взгляд, в науке о поведении очень быстро произошёл поворот от изучения бессознательного поведения к выяснению умственных способностей и социального поведения животных. От классической этологии наука семимильными шагами направилась в сторону когнитивной этологии и социобиологии. При этом многие «доумственные» особенности поведения оказались лишь обозначены, но слабо исследованы. В частности, откровенно недостаточно изучена реакция на раздражители и особенно — на супер-раздражители. В классической этологии есть понятие о накопленной энергии, аппетентном поведении и о ключевом раздражителе, выпускающим энергию. Однако нет понятия о том, что раздражитель может запускать накопление энергии (например, чайки гнездиться не хотят, но раздражитель, возможно, «заставляет» их хотеть).
Заключение
Многолетние наблюдения за птицами разных видов из разных систематических групп показали наличие у них по крайней мере трёх стратегий при выборе места для гнездования. У разных видов эти стратегии имеют разную степень выраженности. Наиболее простой и, видимо, требующей наименьшей мозговой активности является стратегия реакции на небольшое число внешних сиюминутных раздражителей. Более сложной можно считать стратегию консервативных поведенческих установок («традиций»). Самой эффективной с точки зрения выживания вида или популяции выглядит рационально-вероятностная стратегия выбора места гнездования.
Именно последний способ действий выглядит адаптивным. Однако, как это ни странно, инадаптивное поведение при выборе места гнездования распространено у птиц довольно широко. По непонятным причинам рационально-вероятностная стратегия обнаруживается пока у относительно малого числа видов птиц. С утилитарной, практической точки зрения роль внешних раздражителей состоит в том, что при их помощи мы можем заставить птиц отойти от рационально-вероятностного характера организации их поведения и от консервативных поведенческих установок и направить поведение птиц в необходимое (человеку) русло.
Автор выражает искреннюю благодарность директору ФГБУ «ЗаповедникиТаймыра» В.В. Матасову за предоставленную возможность работать на северо-западном Таймыре и поддержку данных исследований. Большую признательность выражаем старшему инспектору отдела охраны «Заповедников Таймыра» И.Н. Корниенко, который непосредственно помогали моим исследованиям.
Список литературы
Дьюсбери Д.М. 1981. Поведение животных. Сравнительные аспекты. М.: Мир. 480 с.
Зубакин В.А. 1998. Распределение и численность чайковых птиц Московской области // Орнитология. М.: Изд-во МГУ. Вып. 28. С. 66–75.
Панов Е.Н. 1973. Организация и эволюция популяционных систем // Вопросы философии, No 11. С. 132-139.
Панов Е.Н. 1978. Механизмы коммуникации у птиц. Москва: Наука. 304 с.
Панов Е.Н., Зыкова Л.Ю., Грабовский В.И. 1990. Биология гнездования, поведение и таксономия хохотуньи Larus cachinnans. 1. Пространственная структура и поведенческие аспекты гнездовой биологии // Зоологический журнал. Т.69. No12. С. 92-105.
Резникова Ж.И. 2005. Интеллект и язык животных и человека. Основы когнитивной этологии. М.: Академкнига. 520 с.
Сыроечковский Е.Е.-мл. 1999. Чёрная казарка (Branta bernicla) в России: экология, распространение, проблемы охраны и устойчивого использования // Автореф. дис. … канд. биол. наук. М.: ИПЭЭ РАН. 35с.
Тинберген Н. 2012. Поведение животных, Москва: АСТ-ПРЕСС. 192 с.
Харитонов С.П. 1993. Гнездовой консерватизм (гнездовая филопатрия) и переселения озёрных чаек Larus ridibundus в пределах колонии // Русский орнитологический журнал. Т. 2. Вып. 3. С. 361-82.
Харитонов С.П. 1998. Верность месту гнездования и особенности переселений озёрных чаек (Larus ridibundus) в пределах группы колоний // Известия Российской Академии наук. Серия биологическая. No2. С. 250-257.
Харитонов С.П., Бубличенко А.Г., Коркина С.А. 2005. Экология гнездования белых сов на северо-западном Таймыре: сопоставление с фазами динамики численности леммингов и пространственное распределение // Совы Северной Евразии / под ред. Волков С.В., Морозов В.В., Шариков А.В. Москва: Рабочая группа по совообразным. С. 23-31.
Харитонов С.П., Волков А.Е., Виллемс Ф., Клейф Х. ван, Клаассен Р.Х.Г. и др. 2008. Колонии черных казарок возле белых сов: расстояния между гнездами в зависимости от численности леммингов и песцов // Известия РАН. Серия Биологическая. No 3. С. 313-323.
Харитонов С.П., Эббинге Б.С., Фоу Дж. Де. 2013. Колонии черных казарок возле белых сов: зависимость расстояния между гнездами от плотности размножающихся песцов // Известия РАН, серия биологическая. No 1. С. 53-59.
Харитонов С.П., Красильников Ю.А., Звонов Б.М., Золотарев С.С. 2016. Восстановление исчезнувшей колонии озёрных чаек (Larus ridibundus) (Charadriiformes, Aves): роль раздражителей в формировании новых колоний птиц // Поволжский экологический журнал. No 4. С. 476-492.
Egan K. Webb G. 2000. A useful strategy for the management of eggs dislonged from the nests of little terns Sterna albifrons // Corella V. 24. No 12. P. 13-14.
Kabamba P.T., Lin W.-C., Meerkov S.M. 2007a. Rational Probabilistic Deciders. Part I: Individual Behavior // Mathematical Problems in Engineering. Article ID 35897. 31 p. doi:10.1155/2007/35897.
Kabamba P.T., Lin W.-C., Meerkov S.M. 2007b. Rational Probabilistic Deciders. Part II: Collective Behavior // Mathematical Problems in Engineering. Article ID 82184. 34 p. doi:10.1155/2007/82184.
Kharitonov S.P. 1999. Environmental factors affecting the breeding success in several gull and tern species at Sivash Lake, the Azov Sea // The Ring. No 1. P. 141.
Scarton F., Valle R. 1997. Laridae e Sternidae nidificanti in laguna di Venezia: anni 1995–1996 // Riv. Ital. Ornitol. V. 67. No 2. P. 203–204.
Walker T.A. 1993. Seabird Islands. Purtaboi Island, Great Barrier Reef, Queensland // Corella. V. 17. No 5. P. 146-148.
White D.H., Mitchell C.A., Swineford D.M. 1984. Reproductive success of Black Skimmers in Texas relative to environmental pollutants // J. Field Ornithology. V. 55. No 1. P. 18-30.
Winton B., Leslie D.M. Jr. 2003. Nest sites and conservation of endagered Interior Least Terns Sterna antillarium athalassos on an alkaline flat in the south-central Great Plains (USA) // Acta ornithologica, V. 38 No 2. P. 135-141.
Вестник ТвГУ. Серия «Биология и экология». 2019. No 1(53) (С материалами 1го Всероссийского орнитологического конгресса).
Рекомендуем прочесть
 Эксперимент по восстановлению исчезнувшей колонии озёрных чаек
Эксперимент по восстановлению исчезнувшей колонии озёрных чаек О недейственности активных форм коллективной защиты колониальных птиц
О недейственности активных форм коллективной защиты колониальных птиц Что не так с социобиологией?
Что не так с социобиологией? Социальная организация гусеобразных (Аnseriformes): базовые принципы и основные направления эволюции
Социальная организация гусеобразных (Аnseriformes): базовые принципы и основные направления эволюции Сигнальное поле как регулятор: от «памяти» и «наследственности» до эволюции
Сигнальное поле как регулятор: от «памяти» и «наследственности» до эволюции Красивое доказательство «семантичности» демонстраций у птиц
Красивое доказательство «семантичности» демонстраций у птиц Птицы-печники против «общих мест» эволюционной экологии
Птицы-печники против «общих мест» эволюционной экологии Кооперативное размножение у врановых птиц
Кооперативное размножение у врановых птиц
