

Южный соловей. Сверху — внешний вид, внизу — гнездовой и зимовочный ареалы (зелёным и синим).
 Далеко не всегда изменения под действием естественного отбора адаптивны. Достаточно часто они “загоняют” вид или популяцию в “ловушку”, ведущую к вымиранию, как глобальное потепление — южных соловьёв, исследованных в Испании Remacha et al., 2020. В чём здесь «ловушка»? На зимовку вид совершает транссахарские перелёты и там в Африке, сильно страдает в годы прихода засух (учащающиеся в связи с потеплением, особенно на юге и Африканском роге) — должны вылетать раньше, их больше гибнет в эти годы, прилетает и гнездится у нас меньше и пр. Важный фактор снижения этих рисков — более длинные крылья, и вроде бы приспособление должно действовать в эту сторону?
Далеко не всегда изменения под действием естественного отбора адаптивны. Достаточно часто они “загоняют” вид или популяцию в “ловушку”, ведущую к вымиранию, как глобальное потепление — южных соловьёв, исследованных в Испании Remacha et al., 2020. В чём здесь «ловушка»? На зимовку вид совершает транссахарские перелёты и там в Африке, сильно страдает в годы прихода засух (учащающиеся в связи с потеплением, особенно на юге и Африканском роге) — должны вылетать раньше, их больше гибнет в эти годы, прилетает и гнездится у нас меньше и пр. Важный фактор снижения этих рисков — более длинные крылья, и вроде бы приспособление должно действовать в эту сторону?
Однако работа по двум популяциям из Центральной Испании показывает, что в последние 20 лет, когда потепление развёртывается вовсю и сдвигает фенодаты не только прибытия, но и отлёта с зимовок, в т.ч. у дальних мигрантов, соловьи меняются в противоположную — и невыгодную — для них сторону, их крыло укорачивается! что делает их ещё более уязвимыми в условиях ухудшения обстановки на зимовках и трассе пролёта.
Почему так получается? В связи с потеплением в Центральной Испании лето стало суше, засухи чаще, суровей и дольше, почему временное «окно» для успешного размножения сократилось, оптимальным стал меньший размер кладки. А, как назло, до всякого потепления, у перелётных птиц, и особенно дальних мигрантов, сформировался так называемый «пакет генов, связанных с миграцией» («migratory gene package«), фактически суперген, регулирующий не только перелёты, но и подгоняющий жизненный цикл к их требованиям. И здесь большая длина крыла ассоциирована с большим уровнем базального метаболизма, большим размером кладки и меньшей продолжительностью жизни — либо наоборот.
Поэтому местный отбор в пользу меньших размеров кладки и более длинной жизни имеет последствия, неблагоприятные для перелётов, в том числе уменьшение длины крыла. Так у североамериканских перелётных певчих дни за время с 1978 г. размер тела в целом стал меньше — правило Бергманна — а крылья длиннее.
И вот благодаря таким корреляциям, установленным в филогенетическом прошлом вида, отбор на приспособление к местным, испанским условиям, имеет губительную «отдачу» в виде увеличения уязвимости при перелётах (при тех же условиях потепления, что его вызвали). Это показывает относительность — и высокую инерционность — адаптации, и объясняет случаи т.н. инадаптивной эволюции, когда отбор эффективен в каждый данный момент, но организация не выдерживает изменений и дохнет (вещь довольно обычная в с/х практике, почему для предотвращения этого приходится отбор по хозяйственно важным признакам сочетать с отбором по приспособленности).
***
Содержание
Фенотипические изменения под действием глобального потепления отмечены многими авторами, число их ширится, как и репертуар затронутых ими признаков. Однако неясно, всегда ли в изменившейся среде обитания подобные сдвиги увеличивают приспособленность, т.е. всегда ли изменения идут в сторону улучшения работы признака в новых условиях (скажем, оптимизируя затраты на них). Крылья дальних мигрантов формировались естественным отбором так, чтобы минимизировать энергозатраты, и изменения крыльев в связи с потеплением должны идти в сторону нового минимума.
Другой вариант — форма крыла изменяется по корреляциям в связи с отбором по другим признакам, также идущим в сторону большей приспособленности в изменившейся среде, что, в конечном счёте, отклонит форму крыла от оптимума и увеличивает затраты на миграцию. Данные 20-летнего мониторинга (1995-2014 гг.) двух популяций южного соловья (Luscinia megarhynchos) из центральной Испании, показывают, наоборот, падение относительной длины крыльев (к размеру тела). В последние десятилетия условия гнездования там ухудшились из-за усиления летней засухи.
Однако длиннокрылые соловьи лучше выживают в первой в жизни миграции (полёт на зимовку и возвращение обратно); чем короче средняя длина крыльев особей, тем сильнее отбор, созданный на дифференциальной выживаемости на пролётных путях. Следовательно, наблюдаемая микроэволюция неадаптивна; конечно, возможно, что больший риск участия короткокрылых птиц в дальних миграциях уравновешивается их преимуществом при репродукции, но данных про это нет.
Считая, что фенотип целостен, а интеграция в него морфологии и жизненной стратегии у дальних мигрантов имеет генетическую основу, авторы родили гипотезу, что неадаптивное укорочение крыльев — коррелированная реакция на отбор в пользу меньших чем ранее репродуктивных вкладов гнездящихся пар при росте засушливости. Это разумно, ведь в этих условиях многочисленное потомство всё равно нечем кормить, но достаточно ли меньшего числа птенцов для сохранения хотя бы прежнего уровня жизнеспособности популяции в условиях неизбежного роста смертности более короткокрылых птиц на миграционных путях? [И судя по приводимым данным, нет. Здесь и далее прим.переводчика].
Глобальные изменения вызывают множество экологических и эволюционных ответов живых существ, включая изменение видовых ареалов, корректировку сезонных циклов и морфологические адаптации (Parmesan and Yohe 2003, Hoffmann and Sgrò 2011, Radchuk et al. 2019). Все такого рода изменения последних десятилетий по умолчанию трактовались как адаптации, созданные отбором, «подгоняющим» организмы к более теплому климату (Lo Cascio Sætre et al., 2017, Weeks et al., 2020), изменениям фенологии (Both et al., 2006) или большей антропогенной трансформации (Johnson and Munshi-South 2017). Однако наше понимание механизмов, лежащих в основе адаптации в быстро меняющемся мире, ограничено сложностью документирования эволюции, идущей сейчас в дикой природе, отсутствием знаний о наследуемости признаков и сложностью коррелирующих реакций, которые могут ограничить или свести на нет преимущество приспособленности адаптивных признаков (Merilä 2012, 2017).
Перелетные птицы — излюбленная модель изучения адаптаций к глобальным изменениям (Helm et al., 2013, Hahn et al. 2016, Vágási et al. 2016). Десятилетия исследований и мониторинга популяций птиц помогли накопить всесторонние знания об экологических и эволюционных детерминантах вариативности миграционного поведения и связанных с ним фенотипических чертах, от сезонной структуры годовых циклов до морфологических адаптаций (Helm et al. 2013, Hahn et al. 2016, Vágási et al. 2016). Сезонность первичной продукции [в более северных местообитаниях] обычно рассматривается основным фактором эволюции миграций, поскольку выгоды большей плодовитости при размножении в высокопродуктивной среде обитания, компенсируют плату в виде подъëма смертности при миграциях, созданных реагированием вида на изменения продуктивности (Thorup et al., 2017).
В таких обстоятельствах селективное преимущество могут получить особям, генетически запрограммированные не только на морфологию и поведение, более подходящую для миграций, но и на “более быструю” жизненную стратегию (Wikelski et al. 2003, Piersma et al. 2005, Van Noordwijk et al. 2006, Dingle and Drake 2007, Liedvogel et al., 2011). Адаптивная интеграция особенностей поведения, физиологии с характеристиками жизненной стратегии была названа “миграционным синдромом” (Dingle and Drake, 2007). Его можно рассматривать как частный случай синдрома характеристик, обеспечивающих “темп жизни”.
Последний описывает жизненную стратегию индивидов и популяций как положение внутри континуума переходов между полюсами градиента, соответствующими «быстрому» vs «медленному» темпам жизни (Ricklefs and Wikelski 2002, Wikelski et al. 2003, Dammhahn et al. 2018). Поэтому более дальние мигранты отличаются не только особенностями морфологии, вроде большей длины крыла, подходящей для быстрого полета (Tellería et al. 2001, Piersma et al. 2005), они также «живут быстрее» [т.е. являются более r- стратегами, тогда как мигрирующие на меньшие расстояния — «более -К«]: у них выше метаболизм покоя, больше размер кладки или меньше продолжительность жизни, чем у менее перелетных птиц (Gwinner et al. 1995, Pérez-Tris and Tellería 2002, Wikelski et al. 2003).
Сегодня предполагается общая генетическая основа разных приспособлений, обеспечивающих миграцию (Liedvogel et al., 2011). Эта гипотеза «пакета генов миграции» поддерживается
(1) данными о генетических корреляциях между названными признаками (Gwinner et al. 1995, Pulido 2007),
(2) популяций с разным уровнем миграционной активности при выращивании в контролируемых условиях лаборатории реализуют каждая фенотип, соответствующий «своему» уровню, и альтернативные между собой (Wikelski et al. 2003),
(3) данные признаки изменяются в ряду популяций, различающихся миграционной активностью, причём изменения разных признаков скоррелированы между собой. Так, удлинение крыла коррелирует с большей плодовитостью или повышенным уровнем базального метаболизма (Gwinner et al. 1995, Pérez-Tris and Tellería 2002, Wikelski et al. 2003);
(4) такие же корреляции между признаками отмечены внутри популяций, исследованных в данном отношении (например, более ранний прилёт коррелирует с большей плодовитостью; Both and Visser 2005, Teplitsky et al. 2011);
(5) эти же признаки согласованно изменяются во времени (Weeks et al. 2020).
Согласно парадигме пакета генов, влияющих на миграцию (на уровне признаков это можно назвать миграционным синдромом), экологические воздействия вне перелета и зимовки, сдвигающие «темп жизни» особей в данной популяции от прежнего оптимума к какому-то новому, меняют по корреляциям и остальные признаки синдрома. Эти скоррелированные изменения в одних случаях облегчают приспособление популяции к соответствующим изменениям, в других, наоборот, сдерживают их (Merilä 2012, Møller et al., 2017). Поэтому парадигма пакета мигрирующих генов удобна для интерпретации наблюдаемых фенотипических изменений, особенно при согласованном изменении разных признаков в направлении, которое трудно интерпретировать как адаптацию (Weeks et al., 2020).
Долговременное прослеживание особей в популяции даёт естественную возможность среду, связать определённые средовые воздействия с определёнными изменениями признаков по типу «фактор — отклик» (Clutton-Brock and Sheldon 2010, Karell et al. 2011, Tellería et al. 2013). Быстрый прогресс глобального потепления в последние десятилетия (IPCC, 2013) стимулировал у перелетных птиц микроэволюционные изменения таких признаков как размер тела, даты прибытия и откладки яиц, величина выводка, и ряд других (Both and Visser 2001, 2005, Yom-Tov et al. 2006, Salewski et al. 2014, Usui et al. 2017; Weeks et al., 2020).
Однако неясна природа этих фенотипических изменений: то ли это приспособление под действием отбора, подталкивающего данные признаки к оптимумам структуры и максимумам приспособленности, актуальным в новых условиях (и тогда эти признаки/изменения должны быть названы адаптивными)? Или же это скоррелированные реакции на отбор, действующий на другие признаки, или просто фенотипическая пластичность, когда вновь возникший предпочтительный фенотип не коррелирует с приспособленностью [это неверно: даже когда он не наследуется, но полезен животному здесь и сейчас, особи отбираются по способности его давать, здесь есть генетическая компонента]. Ответные изменения на воздействия могут даже сдерживать адаптацию, если включают в себя рост частоты неадаптивных признаков. Комбинированный анализ трендов фенотипических изменений и приспособленности индивидов может дать представление об экологических ограничениях адаптируемости перелетных птиц к быстрому изменению климата (Chown et al. 2010, Radchuk et al. 2019).
Южного соловья (Luscinia megarhynchos) можно использовать как модель для ответа на вопросы
1) меняется ли долговременно отношение длины крыла к длине (важное для дальних мигрантов) в ответ на средовую динамику в местах гнездования (т.е. прогресс глобального потепления);
2) адаптивны данные изменения или нет, во 2м случае они ухудшают способность к миграции [или увеличивают риск].
В поисках ответа авторы фиксировали изменения вариации морфологии крыла и зависимую от фенотипа выживаемость в двух рядом находящихся популяциях южных соловьев в центральной Испании, особи из которых прослеживались индивидуально в течение 20 лет (1995-2014 гг.). Также фиксировали местную средовую динамику: изменения экологических факторов, наиболее значимых как детерминанты мест гнездования соловьев за годы наблюдений и изменений фенодат прибытия и размножения. Это существенно, поскольку глобальные тренды климатических изменений не всегда проявляются на местном уровне (или проявляются измененными) (Wilbanks and Kates 1999), т.е. местные популяции широко распространенных видов в ответ на общий тренд потепления климата реагируют разнонаправленно (Hahn et al. 2016).
Временные сдвиги в фенологии весны могут сопровождаться адаптивными изменениями фенотипа в местных популяциях соловьев: длиннокрылые, т. е. быстрей мигрирующие особи выиграют при более раннем начале сезона размножения или укорочении этих сезонов с точки зрения фенологии распускания листьев или доступности насекомых (Hahn et al., 2016). И наоборот, укорочение крыльев может развиться в ответ на поздние весны, ослабляющие отбор на более ранний прилет (Alerstam et al., 2007). Сейчас Средиземноморье — горячая точка [глобального] изменения климата: налицо ярко выраженное потепление [при росте засушливости], уменьшении объема осадков весной и летом (Giorgi and Bi 2005, Giorgi 2006, Giorgi and Lionello 2008). Таким образом, проблема средиземноморских соловьев не во временном сдвиге пика продуктивности или изменении оптимальной даты прилёта [т.е. скорости весенней миграции], но все более засушливый сезон размножения.
Усиливающаяся летняя засуха влияет на птиц в конце сезона размножения, поэтому она может не вести к изменению оптимальных сроков прилёта, способствующих адаптивным изменениям длины крыла. Последнее при прочих равных благоприятствует особям с более медленным (и дешёвым энергетически) темпом жизни, поскольку засушливость сильно ограничивает репродуктивный потенциал средиземноморских птиц, ставя под угрозу жизнеспособность потомства, особенно если рост молодняка перекрывается с периодом водного стресса (Pérez-Tris et al. 2000, Carbonell et al. 2003, Charmantier et al. 2016).
Среди прочего, отбор на медленный темп жизни благоприятствует особям с небольшими кладками, в суровых условиях уменьшенный выводок вырастить легче, меньше родительских усилий требуется. Если у соловьёв, как и у других воробьиных, малый размер кладки и укороченные крылья относительно размера тела коррелируют между собой (Gwinner et al. 1995, Starck et al. 1995, Pérez-Tris and Tellería 2002, Baldwin et al. 2010), отбор может укоротить крыло не вследствие оптимизации морфологии, но из-за коррелированного ответа на отбор, благоприятствующий другому компоненту миграционного синдрома (Piersma et al., 2005, Van Noordwijk et al., 2006).
Тогда укорочение крыльев ухудшит миграционные способности, создавая угрозу выживаемости, отбираемый фенотип получает форму крыла, всё менее адаптивную для задач миграции. Необходим комбинированный анализ морфологической изменчивости и выживаемости, зависящей от фенотипа, чтобы отличить адаптивные морфологические изменения от неадаптивных трансформаций фенотипа (Radchuk et al., 2019), тем самым улучшая нашу способность правильно интерпретировать современные тенденции морфологической изменчивости у перелетных птиц.
Материал и методы
Объект исследования и данные кольцевания
Южный соловей — дальний мигрант, широко распространенный в Европе. Межпопуляционные различия в морфологии связаны с разной дальностью миграции и разной скоростью весеннего развития растительности в пункте назначения миграции (Hahn et al., 2016). Вид был удобен для целей исследования, ибо маховые молодых сохраняются до второго лета жизни, линяя по завершению первого годового круга миграций. Этот позволяет напрямую сравнить летные возможности молодых перед первой миграцией с таковыми вернувшихся весной из транссахарской Африки [т.е. прошедшими отбор миграционным путём]. Южные соловьи гнездятся преимущественно во влажных низменных лесах с конца апреля (первые кладки) до середины июля (самые поздние слетки делаются самостоятельны), обычно откладывая 4-5 яиц. Инкубационный период 13-14 дней, в 10-12 дней птенцы покидают гнездо, хотя начинают летать на 3-5 дней позже (Collar 2005).
Данные авторов собраны на двух станциях кольцевания, работающих еженедельно. Лас-Минас (40,2245°северной широты, 03,5475°западной долготы) расположен в зарослях тростника, птиц здесь кольцуют круглоголично с 1995 года. Преса–дель-Рей (40,3010°северной широты, 03,5414°западной долготы) — пойменный лес, где птиц кольцевания в 1998-2014 гг., только в названный выше сезон размножения (Дополнительный материал S1 и рисунок S2.1 в дополнительном материале S2). Для соловьёв это местообитания противоположного качества: оптимальны в лесу, пессимальны в зарослях тростника (Holt et al., 2010), что облегчает обобщение полученных результатов [на весь спектр видовых биотопов, от лучших до наиболее пессимальных].
Возраст соловьев определяли по состоянию оперения (Jenni and Winkler 1994), различая молодых (этого года вылупления), «новобранцев», «молодое пополнение» местной популяции (второй год жизни, первое размножение в её составе) и старожилов (птицы 2 лет и старше, далее «взрослые»). Пол в анализ не включали: он определяется у взрослых по пению на гнездовой территории или клоакальному выступу, но не у птенцов. У птиц измеряли максимальную ширину хорды крыла (с точностью до 0,5 мм), длину предплюсны (0,01 мм), оценивали жировые запасы (Kaiser, 1993) и взвешивали (с точностью до 0,01 г). Одних и тех же птиц меряли разные (многие) наблюдатели.
Чтобы не путать ошибку измерения, связанную с индивидуальностью наблюдателя, с морфологическими изменениями в онтогенезе, от одного наряда к другому, авторы вычисляли коэффициенты внутриклассовой корреляции (ri; Lessells and Boag, 1987), показывающий повторяемость длин крыла и предплюсны измеренных в один и тот же год у особей одного возраста и наряда. В обоих случаях она высока: длина крыла ri = 0.86, F324,543 = 17.94, P < 0.001; цевки; ri = 0.87, F331,555 = 18.93, P < 0.001.
Данные по гнездящимся птицам данной территории (взрослым или новобранцам) и родившимся здесь молодым отбирали по датам отлова и истории повторных отловов. Консервативная оценка отсутствия здесь мигрантов — с 15 мая по 15 июля для гнездящихся, 31 июля для молодых птиц. К местной популяции отнесены соловьи, пойманные хотя бы один раз в течение этого периода. Для каждого рассчитана средняя длина крыла и цевки по примерам в разных поимках за данный сезон. Подробней о формировании выборки данных см. допматериалы S1.
Средовые переменные [анализируемые в данной работе]
Долговременные изменения условий существования местных соловьёв весной и летом анализировались на основе данных о температуре и влажности в части центральной Испании, включающей оба места исследований, и ряда индексов, с помощью которых на основе спутниковых снимков растительности описывается её развитие и продуктивность. Пространственное и временное разрешение средовых переменной варьировалось сообразно характеру данных.
Весенняя продуктивность. Нормализованный индекс развитости растительности (NDVI) широко используется как мера ее продуктивности в работах по связи динамики численности разных видов животных с изменениями окружающей среды (Pettorelli et al., 2005). Использованы данные по NDVI из проекта анализа растительного покрова и его фенологии (VIP, vegetation index project), серия VIP07 (Дидан и др., 2016; доступны данные с 1982 по 2014 год), полученные в 2 близлежащих областях Центральной Испании (Допматериалы S2). Значения NDVI распределяли по разным видам землепользования, поскольку они определяют растительность, а она — соловьиную фенологию (Badeck et al., 2004). Способы фильтрации значений средовых переменных даны в допматериалах S2. Хол весны каждого года характеризовали: её началом и концом, определённым как дни с максимальным и минимальным изменением (приростом) значений NDVI соответственно), датой максимума продуктивности (день максимума NDVI), темпом развития растительности (разница максимального и начального NDVI данной весны, деленная на число дней между ними).
Летняя температура и влажность. Пик размножения соловьёв летом (июнь–июль в Ц. Испании, см. также Collar 2005). В Средиземноморье это засушливый период, вредно сказывающийся на развитии птенцов (Pérez-Tris et al., 2000). Лето каждого года исследований характеризовали данными температуры и влажности, полученными Национальным центром экологического прогнозирования (Kemp et al., 2012; временной интервал 1950-2014 гг., пространственное разрешение 2,5° широты или долготы), и анализироованными программой RNCEP пакета R. Для каждого лета рассчитаны: даты максимума температуры, их величина, дата минимума влажности, величина влажности воздуха в этот день, скорость роста жары и засушливости в этом сезон (разность значений температуры и влажности между их максимумами и моментами, когда первая стала расти, а вторая падать, деленные на число дней между этими моментами).
Вычисление индексов состояния среды. Вышеназванные характеристики весны и лета в разные годы наблюдений определены при помощи обобщëнных аддитивных моделей (GAM, пакет mgcv в R; Wood 2017). Gam — наиболее мощные из регрессионных моделей, где некоторые вводимые факторы имеют линейную, некоторые нелинейную связь с откликом. С её помощью определяли моменты максимума и минимума производной для аппроксимирующей кривой, полученной сглаживанием по 2хдневным интервалам вместе с её стандартными ошибками. Первый «ловит» начало, второй конец анализируемых сезонов (начало весны и повышения температуры и минимум (конец весны и начало снижения влажности).
Эти модели также использовали для определения даты максимума прогнозируемых изменений NDVI и температуры, и минимума влажности достигла своего минимума. Расчеты выполнены с использованием программного обеспечения R3.5.3 (R Development Core Team 2019). Доля дисперсии, объясненной рассчетной сеткой модели, варьировала для значений NDVI от 19,87 до 85,28% (среднее значение = 55,75%), для данных температуры от 75,42 до 89,46% (среднее значение = 84,90%) и от 34,21 до 69,39% для данных о влажности (среднее значение = 53,28%).
Далее авторы выполнили анализ главных компонент (PCA) для матрицы средовых переменных (использовали вращение varimax, пакет psych в R; Revelle 2019), чтобы выделить факторы, ответственные за главные тренды развития погоды весной и летом в период наблюдений. Координаты по осям этих компонент использовали как обобщённые индексы весенней и летней фенологии. Весну характеризовали
1) насколько она ранняя (противопоставляется поздней весне) и
2) скоростью её наступления (дружная vs затяжная весна).
1) позитивно связан с датой начала весны, датой максимума продуктивности и концом весны (доля объяснений дисперсии R2 = 0,55, коэффициенты факторной нагрузки ≥0,88). Дружность весны позитивно зависит от темпа развития растительности и отрицательно — с её сокращением при переходе к засушливомулету (R2=0,24, нагрузкa ≥0,65). Сходным образом лето описывали двумя факторами:
1) засушливостью (быстрое развитие жары и падение влажности противопоставляется медленному и с большей влажностью),
2) оно раннее vs позднее.
Первый объяснил 41% дисперсии, он позитивно связан с летним максимумом температуры и скоростью его достижения (нагрузка ≥0,78). Значимость фактора (величина R2) росла в более засушливые лета, и при скорейшем развитии засухи (нагрузки ≤-0,54). Второй объяснил 27% дисперсии; он положительно связан с датой температурного максимума и отрицательно — с минимумом влажности (нагрузки ≥0,62).
Фенология насекомых. Насекомые — главная пища птенцов, их обилие в период гнездования определяет успех размножения (Visser et al., 1998). Согласно рекомендациям Emmenegger et al. (2014) и Hahn et al. (2016), авторы характеризовали развитие биомассы насекомых суммой накопленных градусо-дней (СНГД) (Jarošík et al., 2011). Начало высокой доступности насекомых (FAhigh = начало сезона доступности пищи) считали когда накопленные температуры СНГД = 59,1°C/сут., это средняя температура появления ранних стадий личинок насекомых (Jarošík et al., 2011). В рассчетах СНГД суммированы только температуры выше 10,4°C, нижнего порога развития насекомых (Jarošík et al., 2011).
Морфологические переменные. Авторы анализировали тенденции долговременных изменений длины крыла и предплюсны — важных характеристик конструкции летательного аппарата перелетных птиц (Tellería and Carbonell 1999). Однако различия в данных измерениях в первую очередь зависят от размеров тела; для получения независимых от них мер длины крыла и предплюсны использовали анализ главных компонент (Rising and Somers 1989, Pérez-Tris and Telleria 2001). Первая ось пространства ГК (PC1) объяснила 61,2% дисперсии данных промеров, координаты по ней положительно коррелировали со значениями обоих (нагрузка = 0,78). Вторая ось РС2 объясняет оставшиеся 38,8% дисперсии; она положительно связана с длиной крыла (нагрузка = 0,62) и отрицательно — с длиной предплюсны (нагрузка = -0,62). Отсюда значения PC1 использовали как меру линейных размеров, PC2 как меру развития летательного аппарата (MFA), усиливающегося с удлинением крыла и ослабляющегося с удлинением предплюсны.
Обычно крыло у птиц удлиняется с возрастом, особенно за счёт первой полной линьки (De la Hera et al., 2014). Эта пластичность может быть объектом отбора (Perez-Tris и Telleria, 2001), создающего морфологические изменения. Межиндивидуальные различия в длине крыла выявлены дисперсионным анализом её в разном возрасте [для оценки их значимости]. Он убедил авторов, что износ перьев значимо не укорачивает крыло в период от вылупления до второго года жизни, т. е. износ перьев не влияет F1,32 = 1,30, P = 0,26. Тогда индивидуальную пластичность оценивали средним приростом длины крыльев взрослых соловьев относительно длин, когда они были молодыми.
Фенологические переменные. Исследуя зависимость между морфологическими изменениями и фенологией весны/лета, авторы анализировали тренды изменений дат первого отлова гнездящихся птиц и птенцов (прокси соответственно даты прилёта и вылета птенцов). Здесь учли только данные из Лас-Минаса, поскольку в Преса-дель-Рей ловить начали лишь в апреле, когда часть птиц уже прилетела. Чтобы исключить систематическую ошибку, созданную возможным отклонением среднегодовых дат прилета (она возникает, если рано прилетающих особей в действительности ловят позже, и эти даты накапливаются), учитывали даты первого отлова самых ранних 20% особей (от 2 до 6 птиц в зависимости от года). Тот же анализ повторили для данных по всем гнездящимся птицам, пойманным до 15 мая; 15 мая примерно соответствует концу первой волны отловов (их частота распределена бимодально, спад между пиками соответствует гнездовому периоду).
Аналогично средние даты вылета для разных лет определяли по дням первых отловов слетков, сравниваемых с распределением дат отлова всех слетков. Интервал между прибытием и размножением определяли разностью дат отлова самых ранних слетков в Лас-Минасе и 20% самых ранних гнездящихся птиц. Определение дат прилета и вылета слётков чувствительно к величине интервалов между этими двумя периодами отловов в разные годы, поэтому в анализе фенологических изменений исключены годы, когда были перерывы в отлове в моменты, обычно важные для птиц (иногда это вызвано плохой погодой). Все даты переведены в юлианские (1 января = 1).
Анализ временных рядов. Авторы анализировали ряды изменений экологии, морфологии и фенологии соловьёв, оцененных соответствующими переменными. Работали с аппроксимируюшими кривыми изменений, полученными GAM с функцией сглаживания по года на основе метода ограниченного максимального правдоподобия (пакет mgcv в R; Wood, 2017). Эффективные степени свободы (edf) показывают степень сглаживания. Когда лучше соответствуют нелинейной зависимости (edf > 1), также строили общие линейные модели регрессии (при необходимости смешанные) для дальнейшего изучения линейного тренда. Отдельно оценивали автокорреляцию остатков, строили графики, чтобы определить лучшие модели в разные годы (чем меньше остатки коррелируют и ближе к шуму, тем лучше).
Альтернативные методы, вроде расчета функций автокорреляции, применять было нельзя — они требуют постоянства интервалов между элементами ряда. Анализировали тренды изменений MFA, длин крыла, предплюсны, РС1 и РС2, используя сглаживание по годам (а также связь года и возраста, когда это существенно). Индивидуальность особей интерпретировалась как случайно действующий фактора анализа. Возраст и место исследований (в моделях, использующих данные по двум населенным пунктам) включались как фиксированные эффекты, вместе с их взаимодействием, если оно значимо.
Разные формы землепользования учтены как фиксированные эффекты влияющие на индекс NDVI. Статистическая значимость эффектов оценена логарифмом теста отношения правдоподобия (LRT) с использованием распределения хи-квадрат с df = 1 для смешанных моделей и F-статистики для простых моделей. Долговременные изменения средовых переменных смоделированы для всех 20 лет ночного мониторинга соловьев. Чтобы выявить закономерности их изменений в районах исследований, авторы проанализировали их тренды и за более длинный период, с разной продолжительностью для каждой из них в зависимости от временного охвата наличных данных.
Фенотип-зависимые пополнение и выживаемость в популяции. Авторы оценили дифференциальную выживаемость разных фенотипов и дифференциальное пополнение ими местных популяций соловьёв [и через это естественный отбор], используя два подхода. Первый — сравнение MFA впервые отловленных слетков и молодого пополнения контролируемых популяций, второй — оценка различий в среднеожидаемой продолжительности жизни (СОПЖ) у птиц с разным MFA, оценка зависимости MFA и СОПЖ. Первый оценивает зависимую от MFA выживаемость в момент максимальной элиминации в первую осень и зиму жизни молодых, когда они первый раз мигрируют.
В сравнении MFA слетков и молодого пополнения местных популяций использовали технику дисперсионного анализа (ANOVA). Давление отбора за этот период рассчитали как разность MFAп — MFAс, деленную на стандартное отклонение MFAс, где MFAп — средние MFA у соловьев «молодого пополнения», MFAс — оно же для птиц той же когорты, измеренное, когда их ловили слëтками.
Такой рассчет оценивает интенсивность отбора через изменение средних значений отбираемых признаков в единицах стандартного отклонения (Hereford et al., 2004). Соловьи, выжившие [и прибывшие в местную популяцию] включают как повторно пойманных б. слетков, так и не окольцованных птиц, без значимых различий по MFA между ними. Следовательно, поставленные сети ловят почти всех соловьёв, гнездящихся в данном месте или поблизости, и гнездовая дисперсия вместе с натальной не влияет на результаты. LRT-тест для смешанной линейной модели с конкретной популяцией как фиксированным и годом наблюдений как случайным фактором: χ2 = 2,12, P = 0,15.
Дальше использовали регрессионный анализ интенсивности отбора как отклика, зависящего от фактора «год рождения каждой когорты птиц» (исключая гнездовой сезон 2014 г., где отсутствовали данные о пополнении). Также анализировали зависимость интенсивности отбора от MFAс (место наблюдений было фиксированным эффектом). Дальше сравнили изменения средних MFA в размножающихся популяциях разных лет (рассчитанных по объединëнной выборке взрослых и птиц «молодого пополнения), со средним MFA слетков тех же лет, для приблизительной оценки наследуемости MFA.
Выживаниe соловьев на протяжении жизни в зависимости от MFA моделировали программой RMark (Laake and Rexstad, 2008), используя данные поимок-повторных отловов. Этот анализ требует присвоить конкретные значения MFA каждой отдельной особи, однако у перелетных птиц длина крыла растёт с возрастом (Perez-Tris and Telleria, 2001; De la Hera et al., 2014), и соловьи не исключение (см. тренды изменения морфологии ниже). Поэтому использование средних значений MFA разных возрастов исказило бы зависимость, ибо лишь выжившие делаются взрослыми, что искусственно увеличивает их их средние MFA.
Это препятствие обошли стандартизацией значений MFA трёх выделенных возрастов (сеголетки, молодое пополнение и взрослые). Дальше использовали именно стандартизированные по возрасту значения MFAst как характеристику особей, «оцениваемую» отбором. Её и использовали в анализе выживания, зависимого от фенотипа. В тестировании предположений об этих зависимостях использовали программу Mark 6.2 (White and Burnham, 1999), примененную к открытой популяционной модели Кормака-Джолли-Себера (CJS), в её рамках получены оценки текущей выживаемости (Φ) и вероятности повторного отлова в разные годы (p). Так протестировали все комбинации влияний 1) года наблюдений, 2) времени (линейный тренд), 3) возраста при первом отлове, 4) места наблюдений и 5) MFAst на выживаемость и а) константы б) времени (как действующего фактора), в) места наблюдений и г) усилий по сбору выборки на вероятность повторного отлова.
Усилия по отлову птиц включены в регрессионный анализ в качестве константы, различающейся до и после 2009 года в Преса-дель-Рей (когда общая длина тамошней паутинной сети выросла с 66 до 120 м). Использована стратегия выбора моделей с перебором всех комбинаций, следуя Doherty et al. (2012). При проверке предположений CJS, сперва оценили качество моделей прогоняя их программой Release в RMark (P > 0,99 во всех случаях). Мспользовали информационный критерий Акайке, скорректированный с учетом малого размера выборки (AICc), для отбора наилучших моделей (все модели с ΔAICc ≤ 2 были признаны одинаково хорошими; Burnham and Anderson, 2002).
Результаты
Средовая динамика
За годы мониторинга соловьев (рис.1) весна приходит всё раньше (линейные изменения, edf = 4,87, оценка = 0,023, SE = 0,009, F1,93 = 6,95, P = 0,01; рисунок 1), а скорость её развития циклически колеблется (edf = 6,19, F = 3,73, P = 0,001). За более долгий период (1982-2014 гг.) показана нелинейность изменений прихода весны (edf = 2,96, F = 3,19, P = 0,02; рис.1; в скорости развития весны он был не значим, проявился на уровне тенденции, P = 0,15). Оба параметра хода весны значимо зависели от форм землепользования (все значения P < 0,001).
Также фиксируется линейный тренд роста летней засушливости в 1950-2014 гг. (edf = 1,34, оценка = 0,017, SE = 0,006, F1,63 = 7,64, P = 0,007; см. рисунок 1; все остальные значения P ≥ 0,44). Начало сезона высокой доступности насекомых в 1950-2014 гг. колебалось со 101-го по 163-й день юлианского календаря. Фенология насекомых менялась лишь долгосрочно (edf = 2,58, F = 4,02, P = 0,01) по отрицательному линейному тренду (оценка = -0,156, SE = 0,080, на границе значимости F1,63 = 3,83, Р=0,055, см. рис.1, тренд изменений 1995-2014 гг. не значим).
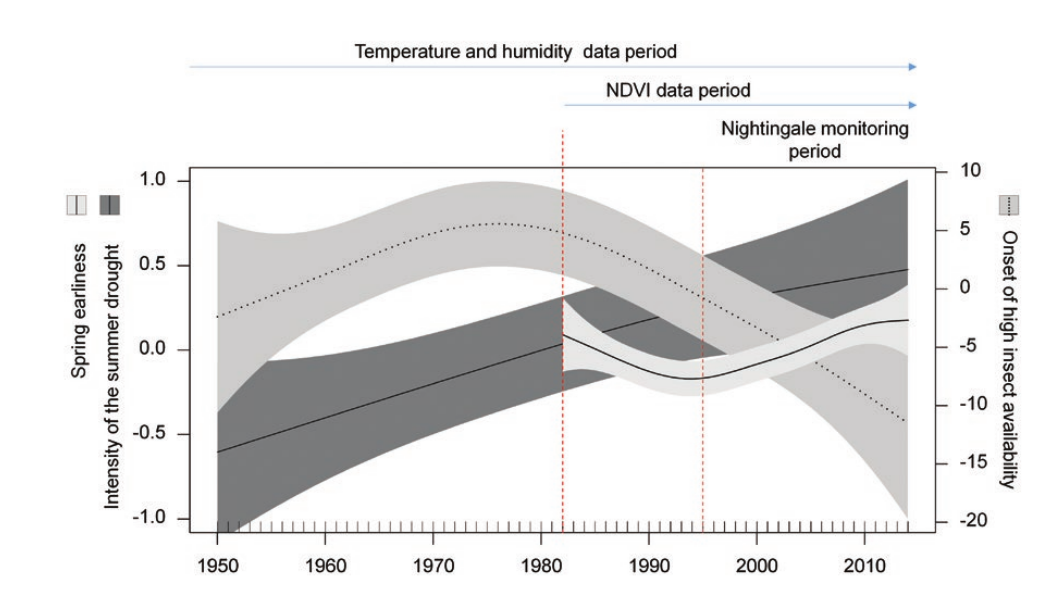 Рисунок 1. Долговременные изменения начала прихода весны (сплошная линия на светло-сером фоне), засушливости лета (сплошная на темно-сером) и периода обилия насекомых (пунктир на сером средней интенсивности), абсцисса — годы мониторинга разных параметров (сверху). Полосы — 95% доверительные интервалы, линии — регрессии.
Рисунок 1. Долговременные изменения начала прихода весны (сплошная линия на светло-сером фоне), засушливости лета (сплошная на темно-сером) и периода обилия насекомых (пунктир на сером средней интенсивности), абсцисса — годы мониторинга разных параметров (сверху). Полосы — 95% доверительные интервалы, линии — регрессии.
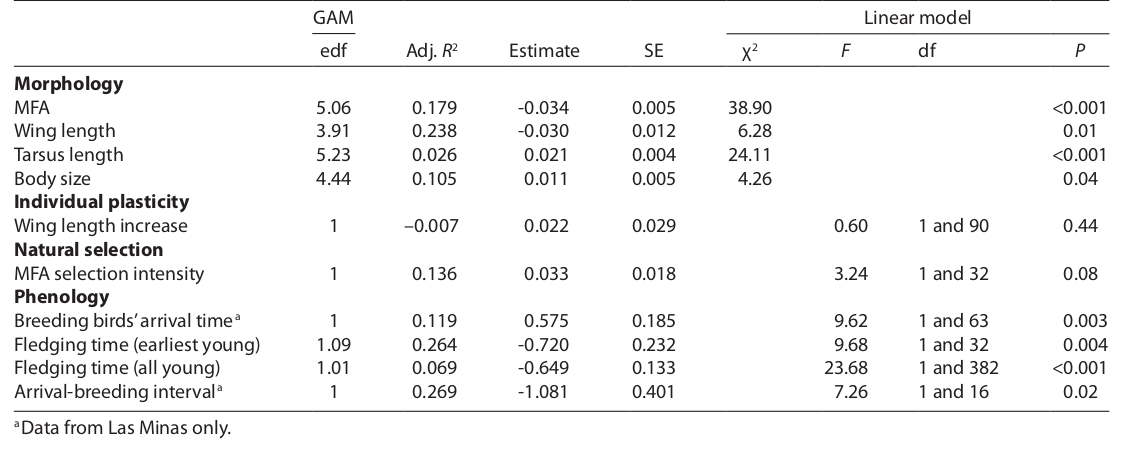 Таблица 1. Долговременный тренд морфологических изменений у южного соловья (значения и их вариация) на фоне динамики характеристик среды. Первый включает изменения длины крыла и предплюсны, в виде индексов структурных размеров, независимых от размеров линейных (PC1 в анализе главных компонент) и развитость летательного аппарата (MFA, PC2), птиц, растущую прямо пропорционально длине крыла и обратно — предплюсны (оба взяты относительно линейных размеров). Также анализируется тренд изменений пластичности по длине крыла (ее индивидуальное увеличение после первой полной линьки) и интенсивность естественного отбора по MFA в связи с первой в жизни миграцией. Фенологические переменные — прилёт гнездовой популяции (по датам отловов 20% самых ранних птиц за год, только для Лас-Минаса), срок вылета (по самым ранним датам отловов слетков, либо по всём пойманным молодым за сезон), промежуток между прибытием и вылетом молодых в днях (измерено только в Лас-Минасе).
Таблица 1. Долговременный тренд морфологических изменений у южного соловья (значения и их вариация) на фоне динамики характеристик среды. Первый включает изменения длины крыла и предплюсны, в виде индексов структурных размеров, независимых от размеров линейных (PC1 в анализе главных компонент) и развитость летательного аппарата (MFA, PC2), птиц, растущую прямо пропорционально длине крыла и обратно — предплюсны (оба взяты относительно линейных размеров). Также анализируется тренд изменений пластичности по длине крыла (ее индивидуальное увеличение после первой полной линьки) и интенсивность естественного отбора по MFA в связи с первой в жизни миграцией. Фенологические переменные — прилёт гнездовой популяции (по датам отловов 20% самых ранних птиц за год, только для Лас-Минаса), срок вылета (по самым ранним датам отловов слетков, либо по всём пойманным молодым за сезон), промежуток между прибытием и вылетом молодых в днях (измерено только в Лас-Минасе).
Тренды тестированы с использованием обобщённых аддитивных моделей (GAM), но линейные — также и с помощью линейных моделей. Для каждого тренда приведены влияние фактора (доля объясненной дисперсии, R2) и величины изменений во времени каждого из откликов со стандартными ошибками. При edf = 1, общие аддитивные модели регрессии эквивалентны линейным. Статистическая значимость линейного тренда (P) была получена с помощью тестов отношения правдоподобия (для моделей со смешанными эффектами, χ2) или статистики F (для линейных моделей с фиксированными эффектами).
Морфологические изменения
За годы наблюдений среднее MFA соловьев падало (таблица 1, рисунок 2). Менялись обе структуры: нескорректированная длина крыла падала (на 0,6 мм), при росте длины предплюсны на 0,4 мм (таблица 1). Линейный размер, оцененный по PC1 для 2 названных признаков, также вырос за эти годы, не сильно, но значимо (таблица 1). Тренды морфологических изменений значимо не отличались в разных возрастах (везде P > 0,12). Для MFA выявлена небольшая, но значимая зависимость между возрастом и местом исследований (LRT: χ2 = 7,27, P = 0,03, см. Доп материалы S3 о различиях). Место наблюдения не влияло на значения никаких других переменных (везде P > 0,12), но длины тела и крыльев выше у взрослых (P < 0,001).
У некоторых птиц крылья стали длинней после первой полной линьки (82,02 ± 0,20 мм vs 84,04 ± 0,18 мм; влияние линьки значимо, ANOVA F1,92 = 180,11, P < 0,001). Не выявлено каких-либо изменений этого приращения [созданного пластичностью] за годы наблюдений (таблица 1).
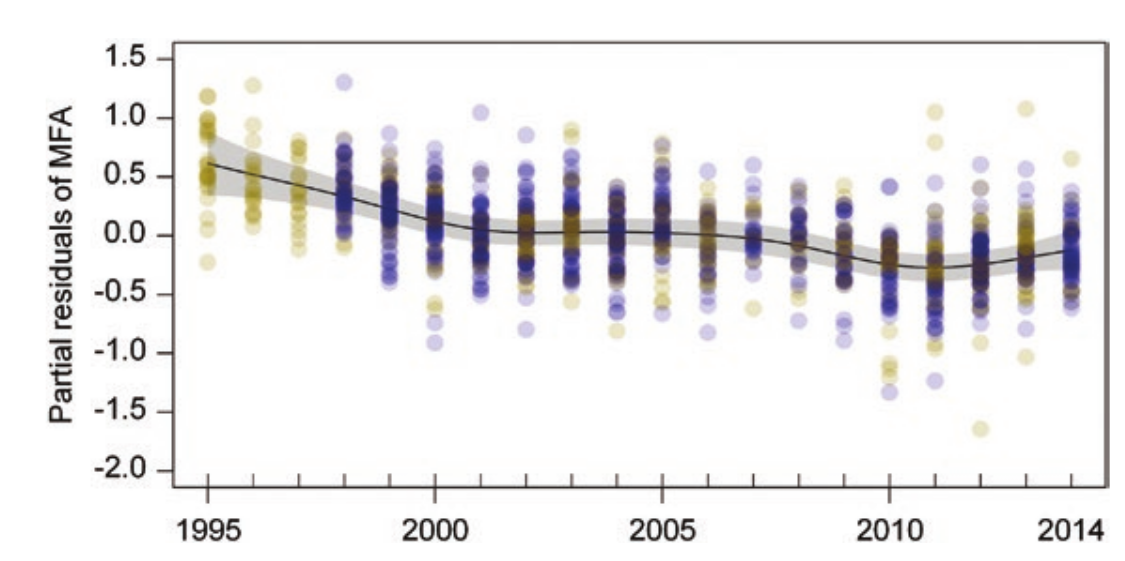 Рисунок 2. Долговременные изменения летательного аппарата южных соловьев (MFA, растёт с удлинением крыла и укорочением ц цевки относительно тела, 1995-2014 гг., еdf = 5,06). График — наилучшее соответствие нелинейной регрессии с 95% доверительными интервалами. Желтые кружки везде — Лас-Минас, синие — Преса-дель-Рей.
Рисунок 2. Долговременные изменения летательного аппарата южных соловьев (MFA, растёт с удлинением крыла и укорочением ц цевки относительно тела, 1995-2014 гг., еdf = 5,06). График — наилучшее соответствие нелинейной регрессии с 95% доверительными интервалами. Желтые кружки везде — Лас-Минас, синие — Преса-дель-Рей.
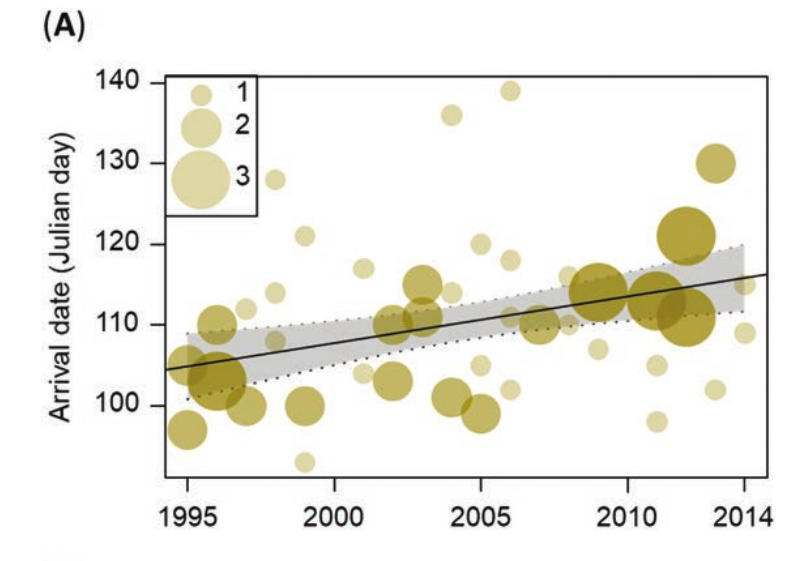
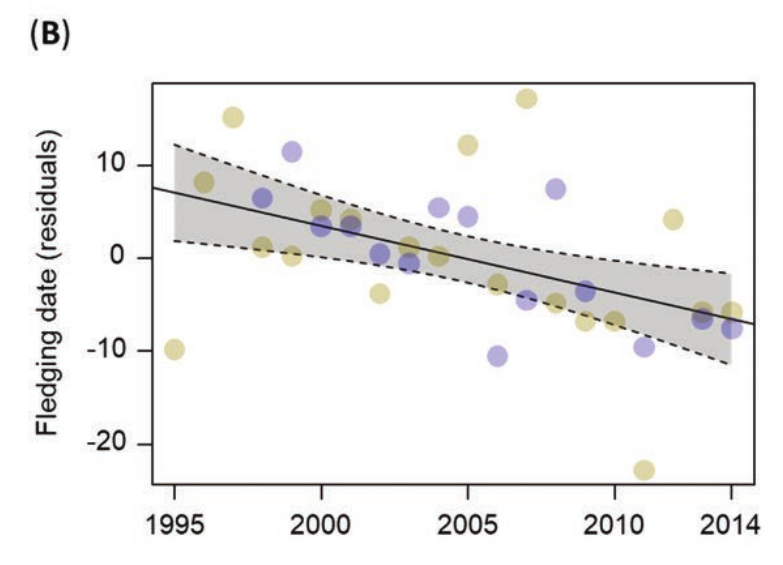
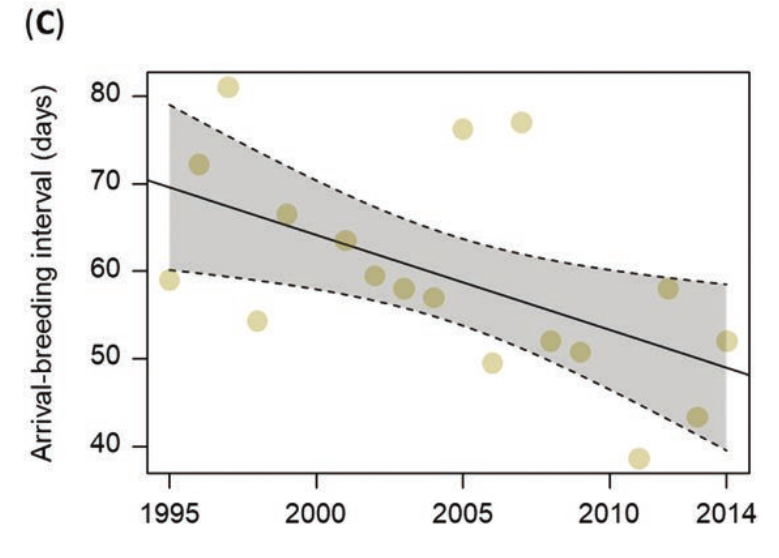 Рисунок 3. Изменения фенодат южного соловья в 1995-2014 гг. (А) Прилёт, по первым отловам 20% самых ранних гнездящихся птиц (только в Лас-Минасе, размер круга показывает размер выборки; 1-й день = 1 января), (В) вылет слетков, остаток от средних дат поимки первых молодых, вылетевших в данном районе и (C) интервал между прилетом взрослых и вылетом слетков. График показывает наилучшее соответствие линейной регрессии с 95% доверительными интервалами (линия и полоса).
Рисунок 3. Изменения фенодат южного соловья в 1995-2014 гг. (А) Прилёт, по первым отловам 20% самых ранних гнездящихся птиц (только в Лас-Минасе, размер круга показывает размер выборки; 1-й день = 1 января), (В) вылет слетков, остаток от средних дат поимки первых молодых, вылетевших в данном районе и (C) интервал между прилетом взрослых и вылетом слетков. График показывает наилучшее соответствие линейной регрессии с 95% доверительными интервалами (линия и полоса).
Изменения фенологии
Даты прибытия 20% самых ранних особей в Лас-Минасе варьировали между 12 апреля 1995 года (n = 4) и 2 мая 2013 года (n = 3), т. е. 20 дней между самым ранним и поздним годами. За годы наблюдений они стал прилетать позже, в среднем на 10,9 дней (таблица 1, рисунок 3А). Эта же величина получается, если её рассчитать по средним датам первого отлова птиц гнездовой популяции, пойманным до 15 мая. В их анализе использовали простые регрессионные модели из-за несходимости в смешанных и близости случайных отклонений к нулю.
Даты первых отловов слетков варьировали между 28 мая 2011 года и 7 июля 2007 года (40 дней между самыми ранним и поздними). Выявлена тенденцию к более раннему вылету; она фиксируется как по датам первых отловов молодых (сдвиг на 13,7 дней вперёд, таблица 1, рисунок 3B), так и по всем датам отловов слетков (сдвиг на 12,3 дня вперед; таблица 1). В Преса-дель-Рей соловьи в среднем вылетали раньше, чем в Лас-Минасе (разница = -3,494, SE = 1,489, F1,382 = 5,51, P = 0,02), но по первым отловам слетков отличий не было (F1,32 = 3,41, P = 0,07). Период между прилетом взрослых и вылетом молодых менялся в Лас-Минасе от 38,6 дней в 2011 году до 81 дня в 1997 году; 42,4 дня между самым коротким и длинным сезонами репродукции. За годы наблюдений последний значимо сжался, на 20,5 дней (линейные изменения, таблица 1, рисунок 3С). Здесь исключены из анализа годы пробелов в выборке (для дат прилёта и интервала между ним и концом размножения это 2000 и 2010 годы в Лас-Минасе, для дат отлова первых слетков 2010 и 2012 годы в Преса-дель-Рей). Корреляции изменений фенологии, морфологии и средовых переменных показаны в допматериале S4.
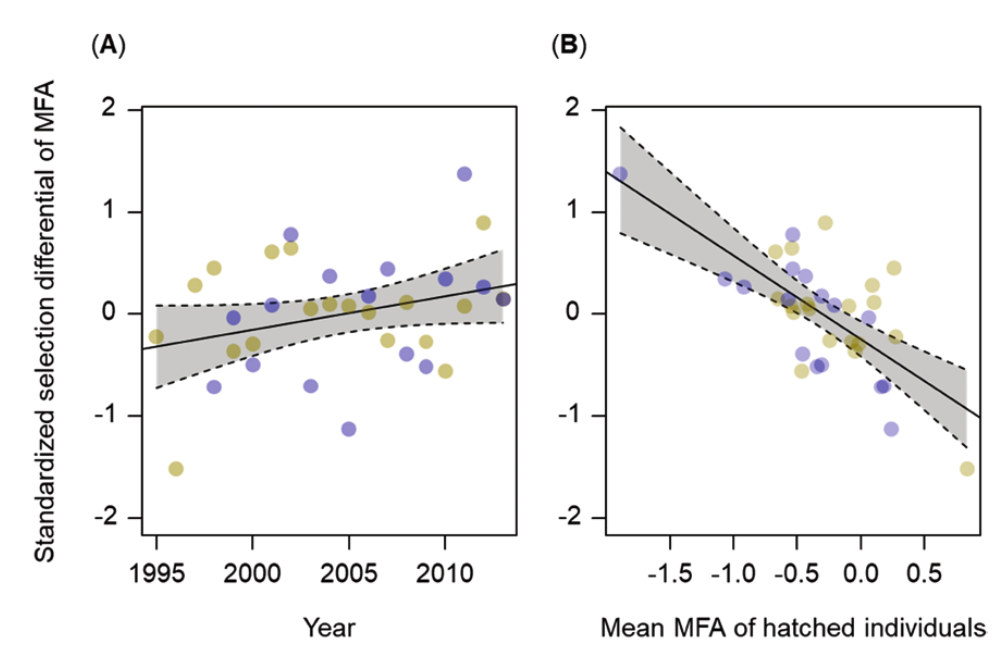 Рисунок 4. Погодовые изменения селективного пресса на соловьев первого года жизни. Показана дифференциальная выживаемость, в зависимости от MFA после двух первых в жизни миграций, в Африку и обратно (наилучшая линейная регрессия с 95% доверительными интервалами). (А) Динамика интенсивности отбора в 1995-2014 гг. и её связь со средними MFA в популяции немедленно после вылета (В). Удаление выбросов в B результатов не изменило.
Рисунок 4. Погодовые изменения селективного пресса на соловьев первого года жизни. Показана дифференциальная выживаемость, в зависимости от MFA после двух первых в жизни миграций, в Африку и обратно (наилучшая линейная регрессия с 95% доверительными интервалами). (А) Динамика интенсивности отбора в 1995-2014 гг. и её связь со средними MFA в популяции немедленно после вылета (В). Удаление выбросов в B результатов не изменило.
Зависимые от фенотипа воспроизводство популяции и выживаемость особей
В целом, соловьи «молодого пополнения» местной популяции, отловленные весной после своей первой в жизни зимовки в Африке, имеют значимо большие MFA, чем сеголетки, отловленные до отлёта туда (ANOVA: F1,692 = 4,73, P = 0,03), что указывает на прошедший отбор в пользу более длиннокрылых и короткоплюсневых птиц. Его интенсивность связана с первой миграцией и имеет тенденцию расти за годы наблюдений, хотя и незначимо (таблица 1, рисунок 4А).
Однако давление отбора здесь росло в когортах с уменьшенным MFA (регрессия = -0,886, SE = 0,156, F1,32 = 32,46, P < 0,001; рисунок 4B). MFA размножающихся соловьёв позитивно связан с MFA их слëтков (оценка = 0,472, SE = 0,095, r = 0,76, F1,18 = 24,77, P < 0,001) [т. е. в значительной степени наследуем].
Наилучшие модели текущей выживаемости CJS (9 моделей с ΔAICc ≤ 2 по сравнению с моделью с минимальным AICc) чаще всего включают возраст при первом отлове, место наблюдений, линейный временной тренд и взаимосвязи между этими эффектами. Однако везде значимо было лишь место (таблица 2; см.все модели в допматериалах S5). Текущая выживаемость соловьёв выше в Преса-дель-Рей, чем в Лас-Минасе (модель с влиянием места и возраста: оценки эффектов = 0,543, SE = 0,024 vs 0,436, SE = 0,029). На обоих участках выживаемость слетков значимо ниже (оценки эффекта ± SE для Лас-Минаса: 0,360 ± 0,047, Преса-дель-Рей: 0,462 ± 0,048), чем у «молодого пополнения» популяции весной (Лас-Минас: 0,463 ± 0,038, Предварительная оценка: 0,568 ± 0,033) или взрослых птиц (ЛасМинас: 0,448 ± 0,040, Преса-дель-Рей: 0,553 ± 0,036). Отметим, что натальная дисперсия молодых и большая филопатрия птиц более старших возрастов вносят свой вклад в различия по текущей выживаемости.
Тем не менее, влияние возраста при первом отлове на текущую выживаемость осталось не значимым по тестам на соотношение правдоподобия, сравнивающим конкурирующие модели с этим эффектом или без него (P > 0,155). MFAST был включен в девятую модель с ΔAICc ≤ 2, и это значимо не влияло на выживаемость (β = 0,019, SE = 0,074; таблица 2).
 Таблица 2. 9 лучших CJS моделей, текущая выживаемость (Φ) и вероятность повторного отлова (p) соловьев как функция возраста на момент первого отлова, района наблюдений, года (линейная аппроксимация), MFAST (стандартизированный по возрасту показатель, который увеличивается с увеличением длины крыла и уменьшением предплюсны длина по сравнению с размером тела), а также усилий по отбору проб. Модели упорядочены по информационному критерию Aкайке с поправкой на малые размеры выборки (AICc). В таблице перечислены 9 моделей, которые показали наилучший баланс между соответствием данным и сложностью (те, у кого были самые низкие значения AICc; модели с ΔAICc ≤ 2 по сравнению с моделью с самым низким AICc, считаются одинаково хорошими). Лучшая модель, включая MFAST, выделена жирным шрифтом. Наилучшая модель, которая не учитывала влияние района наблюдений на Φ, также приведена в нижней части таблицы. Указано количество параметров в каждой модели (k) и веса Акайке (wi, на основе всех возможных моделей).
Таблица 2. 9 лучших CJS моделей, текущая выживаемость (Φ) и вероятность повторного отлова (p) соловьев как функция возраста на момент первого отлова, района наблюдений, года (линейная аппроксимация), MFAST (стандартизированный по возрасту показатель, который увеличивается с увеличением длины крыла и уменьшением предплюсны длина по сравнению с размером тела), а также усилий по отбору проб. Модели упорядочены по информационному критерию Aкайке с поправкой на малые размеры выборки (AICc). В таблице перечислены 9 моделей, которые показали наилучший баланс между соответствием данным и сложностью (те, у кого были самые низкие значения AICc; модели с ΔAICc ≤ 2 по сравнению с моделью с самым низким AICc, считаются одинаково хорошими). Лучшая модель, включая MFAST, выделена жирным шрифтом. Наилучшая модель, которая не учитывала влияние района наблюдений на Φ, также приведена в нижней части таблицы. Указано количество параметров в каждой модели (k) и веса Акайке (wi, на основе всех возможных моделей).
Обсуждение
Параллельно с глобальными изменениями [особенно климатическими] у многих перелётных птиц идут фенотипические изменения, хотя [в большинстве случаев] остаётся малоизученным, приспособительные они или вынужденные средовым трендом. За 20 лет мониторинга (1995-2014 гг.) крыло южных соловьев в Центральной Испании укорачивалось относительно тела, которое ещё и несколько выросло. Птицы не просто уменьшились: изменилась «конструкция» летательного аппарата (Dunn et al. 2017). Данный тренд равно зафиксируется в тростниковых зарослях и в лесу — местообитаниях, контрастирующих по качеству и структуре растительности (Holt et al., 2010).
Сравнение изменений морфологии с выживаемостью, зависящей от фенотипа, показывает ухудшение усредненной формы крыла с точки зрения миграционных способностей птиц. Во время первой в жизни миграции давление отбора благоприятствует длиннокрылым особям и, чем короче крыло, тки сильней в популяции отбор в пользу большей длинны крыла. Авторы не нашли доказательств лучшей выживаемости более длиннокрылых соловьëв в любом возрасте, однако у них выше шанс пополнить собой размножающуюся часть популяции — и вопреки всему этому крыло укорачивалось, а частота длиннокрылых птиц падала из поколения в поколение.
Следовательно, лучшие мигранты попали в явно невыгодное положение; данный факт исключает возможность укорочения крыльев из-за ослабления отбора, вызванного снижением миграционных затрат. Данные [повешенных на птиц] геолокаторов показывают зимовку соловьев в районе Сахеля независимо от пути полёта туда (Hahn et al., 2014), причём самый короткий путь именно у испанских птиц (Hahn et al., 2016). Однако единственный способ сократить дальность миграционного пути было бы зимовать, не пересекая Сахару, однако до сих пор ни разу не сообщалось о соловьях, зимующих севернее неё.
Изменение длины крыльев соловьев соответствовало средовым трансформациям [в местах гнездования]. [Из-за глобального потепления] весна в центральной Испании наступает всё позже, при усилении летней засушливости, т. е. сезон репродукции укорачивается. [Одновременно общее потепление] сдвигает вперёд даты начала размножения насекомых. Всё это соответствует общим тенденциям изменения климата в Средиземноморье (Giorgi and Bi 2005). Соответственно им прилет соловьев задерживается, но вылет птенцов сдвигается на более ранние даты. Скорее всего, этот сдвиг фенологии — реакция на усиливающуюся летнюю засуху (Carbonell et al., 2003). В более северных частях ареала летняя засуха не бывает проблемой, в Испании её последовательное усиление снижает приспособленность птиц, не успевающих вырастить птенцов до её максимума (Pérez-Tris et al. 2000).
[Поскольку] укорочение крыльев снижает выживаемость при первой миграции, его реализация в контролируемых популяциях, видимо, вызвана лучшей размножаемостью более короткокрылых птиц. Из концепции темпа жизни следует что экстремизация среды обитания создаёт предпочтение морфоэкологическим изменениям, уменьшающим затраты на выращивание потомства (Grant and Grant 2002, Ricklefs and Wikelski 2002). [Другое дело, что, снижая затраты и поддерживаясь отбором, они так и не создают адаптации].
Фактически соловьи, столкнувшиеся с укорочением периода, наиболее удобного для размножения, вынужденные выращивать птенцов под «ударами» растущей засушливости, пробуют выиграть что-то для себя здесь и сейчас сокращением инвестиций в воспроизводство [авторы называют это выигрышем, однако именно в эволюционном смысле это не выигрыш, т. е. не адаптация. Эволюция идёт в популяциях, а жизнеспособность местных популяций данное направление изменений подрывает не меньше, чем рост смертности на миграциях, следующий из укорочения крыльев]. Так или иначе, в сложившейся обстановке здесь и сейчас отбор поддерживает уменьшение кладок, птицы с обычным прежде размером кладок чем дальше, тем больше оказываются в невыгодном положении (Boyce and Perrins 1987).
Согласно концепции пакета генов-регуляторов миграции, поддержанное отбором кладки ведёт к уменьшению длины крыла вследствие корреляции соответствующих генов (Van Noordwijk et al., 2006, Merilä, 2012). Фактически, сравнение популяций воробьиных-мигрантов, различающихся по темпу жизни [одни виды более «быстрые», другие — более » медленные»], убедительно доказывает что более медленный темп жизни коррелирует с более короткими крыльями относительно тела и большими PC1-2 (Gwinner et al. 1995, Starck et al. 1995, Pérez-Tris and Tellería 2002, Wikelski et al. 2003, Baldwin et al. 2010). [Однако это видно в межвидовых сравнениях, можно ли результат перенести на внутривидовую изменчивость? В общем случае это неясно, каждый раз надо смотреть отдельно].
Примечательно, что недавнее исследование 52 видов североамериканских перелетных птиц выявило общее уменьшение длины предплюсны, но увеличение длины крыльев параллельно с потеплением климата (Weeks et al., 2020). Однако у южных соловьев морфология меняется в противоположную сторону и, видимо, это инадаптивно! С другой стороны, эти данные подтверждают концепцию миграционного синдрома «от противного» — селективные изменения одного признака из состава синдрома вызывают изменения другого, с ним скоррелированного, итог же обоих подрывает жизнеспособность популяции, а не усиливает её. Обычно считается, что подобные изменения адаптивны (Weeks et al., 2020), но могут и скорей навредить, чем помочь, как в случае укорочения крылья соловьев (Radchuk et al., 2019).
Не имея данных о репродуктивном успехе, авторы для проверки этого предположения используют разные линии доказательств. Более поздние кладки, сопряженные с поздним прилетом, уменьшаются (Rowe et al., 1994, Smith and Moore, 2005). Это адаптивно, если уменьшение репродуктивного усилия ускорит размножение соловьев. Птицы, дающие меньшие кладки (или меньше тратящиеся на выращивание потомства), после прилета могут начать кладку раньше, чем соловьи с большим репродуктивным усилием (Rowe et al., 1994). Действительно, из молодых весной раньше вернулись более короткокрылые птицы, более длиннокрылые позже. У раньше вылупляющихся более короткокрылых птиц есть ещё одно преимущество: активный рост не совпадёт с пиком засушливости — главным стрессором (Perez-Tris et al., 2000), у них есть время улучшить физическое состояние перед миграцией (Gill et al., 2014).
Согласно этой модели, выигрыш более короткокрылых птиц размножении уменьшит со временем длину крыла в популяции. Хотя авторская интерпретация отдает предпочтение преимуществам размножения, морфология крыльев также может меняться при корреляции с признаками, благоприятствующими [достижению] мест зимовки, либо напрямую, либо через различные последствия для индивидуальной приспособленности (Imlay et al., 2019). Район Сахеля сегодня — яркий пример опустынивания, поддерживаемого глобальным потеплением (Huang et al., 2016). Если в этих условиях предпочтительны особи с более низким уровнем метаболизма в состоянии покоя, это способствует выбору медленного темпа жизни.
В большинстве регионов Европы более теплые вёсны способствуют более ранним прилетам перелетных видов (Usui et al., 2017, Mayor et al. 2017), хотя реакция дальних мигрантов выражена слабей, чем у ближних (Rubolini et al., 2010). Южные соловьи в Центральной Испании, наоборот, за прошедшие 20 лет прилетали все позже и позже, что согласуется со все более поздними вëснами, хотя явно расходится и с более ранним появлением насекомых, так и более интенсивной летней сушью.
Фактически они попадают между молотом и наковальней, приспособиться к этим «ножницам» очень и очень трудно, происходящие изменения скорей загоняют вид в «эволюционную ловушку«.
Два этих фактора прямо влияют на успех размножения, и должны способствовать более раннему прилету (Saino et al., 2011). Однако эти же наблюдения показывают много большую ограниченность диапазона возможных дат прибытия у дальних мигрантов. С одной стороны, ему лучше соответствовать максимуму продуктивности среды обитания [доступности и обилию насекомых и других ресурсов, важных для репродукции], с другой — досягаемость мест размножения за известное время для птиц, «выдвигающихся» с мест зимовок, зависит от их морфологии (Møller et al., 2017). Более ранний прилет предполагает большую скорость движения, а она — удлинение крыльев (Hahn et al., 2016). Действительно, одно из селективных давлений, под прессом которых живут обе соловьиные популяции — на удлинение крыльев; чем короче они в популяции, тем он сильнее и лучше выживаемость более длиннокрылых птиц.
Однако неблагоприятные условия размножения более длиннокрылых птиц могут препятствовать вроде бы адаптивному удлинению крыльев у соловьев. Выявив связь между определённой морфологией летательного аппарата и уменьшением численности соловьев, авторы поставили под сомнение идею о том, что тренды морфологических изменений у перелетных птиц имеют всегда адаптивный смысл и объясняются биомеханически (Radchuk et al., 2019). Это «готовит почву» для объяснений, отличающегося от доминирующего сейчас панадаптационизма [см.его критику 1, 2, 3, 4], например, эволюцией неадаптивных признаков по корреляциям [в данном случае генетическим: влияющие на них гены находятся в генетическом тексте «неподалёку» от генов, попадающих под отбор)]. Это значит, что фиксируемые тренды имеют генетическую основу (Gienapp et al. 2008, Hoffmann and Sgrò 2011, Teplitsky and Millien 2014).
Подтверждая эту идею, авторы опровергли предположение, что отмеченные изменения формы крыла созданы фенотипической пластичностью. Кроме того, MFA молодняка все годы наблюдений устойчиво коррелировал с MFA размножающихся, подтверждая наследуемость данного признака. Наследуемость длины крыльев показана для многих видов птиц (Merilä and Sheldon 2000, Teplitsky et al. 2009), и соловьи, вероятно, не исключение (Kipper et al., 2006).
Учёных сейчас беспокоит вопрос — смогут ли дальние мигранты приспособиться к современному изменению климата [, поскольку риски для них максимально повышены сравнительно не только с оседлыми видами, но и с ближними мигрантами] (Radchuk et al., 2019). В ряде работ изучали последствия глобального потепления для их демографии (Sæther et al. 2004). Влияя на фенологию, потепление климатаможет менять разные аспекты жизненного цикла вида, что требует всестороннего изучения множественных последствий (Stenseth and Mysterud 2002, Winkler et al. 2002).
Отбор в пользу более «медленного« темпа жизни может быть объяснением общего неблагополучия тех видов и популяций, у которых даты прилёта не сместились вперёд (Møller et al. 2008, Saino et al. 2011). Он же возникнет при усилении расхождения сроков прилёта мигрирующего вида со сроками весеннего развития растительности (Mayor et al. 2017) [и тоже ведёт к «замедлению» жизненного цикла]. Однако эволюция в эту сторону для дальних мигрантов опасна — хотя приспособленность индивидов формально и возрастает, одновременно растёт риск гибели популяции в целом [из-за подъёма катастрофической, неселективной смертности в годы климатических аномалий в местах зимовок, или остановок на пролёте, а их частоту потепление увеличивает] (Siliceo and Díaz 2010, Møller et al. 2017, Wegge and Rolstad 2017).
Число и величина наших пробелов в знаниях [о (микро)эволюционных ответах на изменения климата] пропорциональны сложности возможных реакций популяции. Это подчеркивает важность долговременного мониторинга отдельных контролируемых популяций, фиксации сдвигов в морфологии, жизненном цикле и пр. (Clutton-Brock
and Sheldon 2010, Tellería et al. 2013, Radchuk et al. 2019), увязывая их, по возможности, с количественной же оценкой средовых изменений и определением генетической детерминации меняющихся признаков. Вместе все 3 составляющие дают инструмент понимания адаптивных реакций птиц на глобальные изменения.
Рекомендуем прочесть
 Эль-Ниньо и Ла-Нинья: почему мальчик и девочка, меняющие климат, стали приходить чаще
Эль-Ниньо и Ла-Нинья: почему мальчик и девочка, меняющие климат, стали приходить чаще Как изменение климата способствует быстрой эволюции
Как изменение климата способствует быстрой эволюции В эпоху глобального экологического кризиса города — не только «проблема», но и “решение”
В эпоху глобального экологического кризиса города — не только «проблема», но и “решение” Механизм фракционирования изотопов углерода при фотосинтезе и углекислотная составляющая парникового эффекта
Механизм фракционирования изотопов углерода при фотосинтезе и углекислотная составляющая парникового эффекта В Старом и Новом Свете птицы сходно реагируют на глобальное потепление
В Старом и Новом Свете птицы сходно реагируют на глобальное потепление Глобальное потепление, опустынивание/деградация почв и засухи в аридных регионах
Глобальное потепление, опустынивание/деградация почв и засухи в аридных регионах Глобальное потепление изменяет леса лишь по нарушениям лесного полога
Глобальное потепление изменяет леса лишь по нарушениям лесного полога Глобальное потепление бьёт по клювам
Глобальное потепление бьёт по клювам
