

Наконец-то перевели на русский старую и очень известную (на Западе) статью двух классиков эволюционной теории, которая – как ушат холодной воды в сегодняшние времена торжества «однониточных» объяснений, пан-адаптационистских в том числе. Скажем, присутствующая здесь критика социобиологии действенна и по сю пору.
Стивен Лж. Гулд, Ричард Ч. Левонтин
Перевод И. Кузина
Предисловие переводчика
Содержание
Палеонтолога С.Дж. Гулда и генетика Р.Ч. Левонтина относят к крупнейшим эволюционным биологам второй половины XX в. (Koertgе, 200S; Ruse. 1999). Их статья 1979 года «Пазухи собора святого Марка и парадигма Панглосса» входит в различные антологии, посвященные философии биологии (Rosenberg, Arp, 2010; Sober, 2006), и является, вероятно, самой цитируемой статьей в этой области. Изобретательная критика панадаптационизма, предпринятая в данной работе, оказалась важным этапом в осознании сообществом эволюционистов сущности синтетической теории эволюции и стимулировала попытки преодолении заложенных в ней ограничений. В конечном счете Гулд включил панадаптационизм в качестве одного из базовых принципов в структуру дарвинизма, как она была представлена и подвергнута ревизии в его opus magnum «Структура эволюционной теории» (Gould, 2002).
Эта критика имела и определенные политические импликации, так как се мишенью оказалась, в том числе, социобиологии Эдварда О. Уилсона (Wilson, 1975), часто ассоциируемая с биологическим детерминизмом и правыми взглядами (Ruse, 2008). Архитектурный термин «spandrel» с легкой руки Гулда и Левонтина перекочевал в биологию, а богатству риторических приемов, используемых в их статье, была посвящена целая книга (Sеlzcr, 1993). Разнообразие выразительных средств, используемых в данной статье, возможно, повлияло на то, что она до сих пор не была переведена на русский язык.
Литература
Gould S.J. The structure of evolutionary theory. — Cambridge, London: The Belknap Press of Harvard University Press, 2002.
Koertge N. (ed.) New dictionary of scientific biography. — Detroit, MI : Charles Scribner’s Sons/Thomson Gale, 2008.
Rosenberg A., Arp R. (ed.). Philosophy of biology: An anthology. — Wiley- Blackwell, 2010.
Ruse M. Mystery of mysteries: Is evolution a social construction? Cam- bridge, London: Harvard University Press, 1999.
Ruse M. The evolution wars: a guide to the debates. — Grey House Publishing, 2008.
SelzerJ. (ed.). Understanding scientific prose. — Univ. of’ Wisconsin Press, 1993.
Wilson E.O. Sociobiology: The New Synthesis. — 1975.
***
Адаптационистская2 программа доминировала в эволюционной мысли Англии и Соединенных Штатов в течение последних сорока лет. Она основана на вере в силу естественного отбора как оптимизирующего фактора. Она реализуется посредством разбиения организма на единичные «признаки» и выдвижения адаптивного сценария для каждого из них по отдельности. Единственным препятствием к достижению совершенства считается неизбежность компромиссов3; неоптимальность, таким образом, также трактуется как результат адаптации. Мы критикуем этот подход и предпринимаем попытку вновь утвердить альтернативную точку зрения (длительное время бывшую популярной в континентальной Европе), что организм должен рассматриваться как интегрированное целое, план строения которого настолько ограничен филогенетическим прошлым, путями эмбрионального развития и архитектурой строения в целом, что эти ограничения сами по себе оказываются более интересными и более важными в определении путей эволюционного изменения, чем селективные силы, которые могут опосредовать изменение, когда оно происходит.
Мы обвиняем адаптационистскую программу в неспособности различать непосредственную полезность и причины ее возникновения (самцы тираннозавров могли использовать свои миниатюрные передние лапы для щекотания самок, но это не объясняет, почему их передние лапы стали такими маленькими), в нежелании рассматривать альтернативы адаптивным сценариям, в том, что она полагается на одно лишь правдоподобие в качестве критерия для принятия гипотетических сценариев, и в неспособности адекватно учитывать такие альтернативы, как случайную фиксацию аллелей4, возникновение неадаптивных структур из-за онтогенетических корреляций с признаками, по которым идет отбор (за счет аллометрии5, плейотропии6, материальной компенсации7, механически вынужденной корреляции), возможность раэобшения адаптации и естественного отбора, существование множества адаптивных пиков и возникновение непосредственной полезности как эпифеномена неадаптивных структур. Мы поддерживаем плюралистический подход самого Дарвина в установлении эволюционных факторов.
Введение
Мозаика огромного центрального свода кафедрального собора Святого Марка в Венеции представляет собой детальную иконографию основ христианской веры. Вокруг центральной фигуры Христа расположены три круга изображений: ангелов, апостолов и добродетелей. Каждый из кругов поделен на квадранты, хотя свод сам по себе радиально симметричен. Каждый квадрант соприкасается с одной из четырех пазух между арками, подпирающими купол. Пазухи — сужающиеся к концам треугольники, сформированные пересечением двух округлых арок под прямым углом (рис.1), — являются неизбежными побочными продуктами сооружения купола на округлых арках.

Каждая пазуха несет рисунок, замечательно вписанный в сужающийся к концам треугольник. В верхней его части — изображение сидящего евангелиста, окружен носе боков изображениями небесных градов. В нижней части — фигура мужчины, выливающего воду из кувшина в сужающееся пространство под своими ногами, олицетворяющая одну из четырех библейских рек (Тигр, Евфрат, Инд и Нил).
Эта композиция так продумана, гармонична и целесообразна, что мы склонны рассматривать её как исходный пункт анализа, в некотором смысле — как причину окружающей архитектуры. Но в таком случае правильный путь анализа будет извращен. На самом деле система начинается с архитектурного ограничения: с необходимости четырех пазух свода, имеющих форму треугольников ссужающимися концами. Они предоставляют пространства для работы мозаицистов; они же задают четырехчастную симметрию купола над собой.
Примеров подобных архитектурных ограничений множество, и они легки для нашего понимания, потому что мы не проецируем на них биологические предубеждения. У любого веерного свода должен быть ряд свободных пространств вдоль его продольной оси, в местах пересечений веерных перекрытий между колоннами (рис. 2).

Любой, кто попытается утверждать, что эта структура существует, потому что чередование роз и спускных решеток так уместно в часовне Тюдоров, спровоцирует те же насмешки, которыми Вольтер осыпал доктора Панглосса: «…все таково, каким должно быть… все необходимо и создано для наилучшей цели. Вот, заметьте, носы созданы для очков, потому мы и носим очки. Ноги, очевидно, назначены для того, чтобы их обувать, вот мы их и обуваем»8.
Однако эволюционные биологи, с их тенденцией сосредоточиваться исключительно на непосредственной адаптации к локальным условиям, склонны игнорировать архитектурные ограничения и осуществлять именно такую инверсию объяснения.
Вот более близкий пример, фигурирующий в некоторых недавних биологических работах по адаптации: антрополог Майкл Харнер предположил (1977), что человеческие жертвоприношения у ацтеков возникли для решения проблемы хронической нехватки мяса (конечности жертв часто съедались, но только высокопоставленными ацтеками). Э.О. Уилсон (1978) использовал это объяснение в качестве принципиального примера адаптивной, генетической предрасположенности к хищничеству у людей. Харнер и Уилсон предлагают нам рассматривать детально разработанную социальную систему и сложный набор эксплицитных обоснований, включая миф, символ и традицию, как всего лишь эпифеномен, созданный ацтеками в качестве бессознательной рационализации, маскирующей «действительную» причину всего этого: потребность в белке. Однако Салинс (1978) утверждает, что человеческое жертвоприношение представляет лишь одну из частей замысловатой культурной структуры, которая в своей целостности не только является материальным воплощением ацтекской космологии, но и выполняет такие утилитарные функции, как поддержание социальной стратификации и системы даннических отношении между городами.
У нас есть серьезные основания подозревать, что ацтекский каннибализм был такой же «адаптацией» как евангелисты и «реки» в пазухах свода или рельефные украшения в свободных пространствах свода: вторичным эпифеноменом, представляющим плодотворное использование имеющихся частей, а нс причину системы в целом. Грубо говоря, система, созданная для других целей, генерирует новые части, из которых тоже может быть извлечена польза. Зачем выворачивать наизнанку таким странным образом всю систему и считать целую культуру эпифеноменом необычного способа расширения мясного рациона? Пазухи свода не существуют для того, чтобы в них располагались евангелисты.
Более того, как считает Салинс, даже не ясно, было ли вообще человеческое жертвоприношение адаптацией. Человеческие культурные практики могут быть «ортогенетическими»9 и приводить к вымиранию таким путем, который невозможен для дарвиновского процесса, основанного на генетическом отборе. Так как каждый монарх должен был превзойти своего предшественника еще более изощренным и обильным жертвоприношением, то такая практика приводила бы к истощению ресурсов. Это был бы не первый случай, когда человеческая культура погубила саму себя. И, наконец, многие эксперты сомневаются, в первую очередь, в базовой предпосылке, которую использует Харнер (Оrtiz de Montellano, 1978). Они считают, что запас других источников белка не был ограниченным и что практика вознаграждения мясом людей, занимающих привилегированное положение, у которых и так было его достаточно, и которые использовали тела так неэффективно (в ход шли только конечности, и те лишь частично) представляет собой очень плохой способ управления «мясной отраслью».
Мы намеренно расположили эти небиологические примеры в ряд по возрастанию близости к биологии: от архитектуры к антропологии. Мы поступили так, потому что первичность архитектурного ограничения и вторичная природа адаптации не замутнены в этих примерах нашими биологическими предрассудками. Но мы надеемся, что послание к биологам не осталось незамеченным: если бы это были биологические системы, не посчитали ли бы мы, в силу привычки, эпифеноменальную адаптацию первичной и не попытались ли бы выстроить структуру всей системы, исходя из этого пункта?
Адаптационистская программа
Мы надеемся поставить под сомнение глубоко укоренившуюся среди исследователей эволюции мыслительную привычку. Мы называем ее адаптационистской, или панглоссовой, программой. Она основана на представлении, популяризованном А.Р. Уоллесом и А. Вейсманом (но, как мы увидим, не Дарвином) к концу девятнадцатого века: естественный отбор практически всемогущ в изобретении конструкций10 организмов и создании лучшего из миров. Эта программа рассматривает естественный отбор как столь могущественный фактор, а ограничения считает столь немногочисленными, что прямое возникновение адаптации под его действием оказывается главной причиной практически всех органических форм, функций и разновидностей поведения. Конечно, существование ограничений на всепроникающую силу естественного отбора признается (в первую очередь, филогенетической инерции11, хотя непосредственные архитектурные ограничения редко признаются, как было рассмотрено в предыдущем разделе). Но они обычно отвергаются как несущественные или даже, что ещё больше удручает, признаются, но не принимаются всерьез и не используются.
Исследования, проводимые в рамках адаптационистской программы, обычно осуществляются в два этапа:
1. Организм фрагментируется на «признаки» и эти признаки интерпретируются как структуры, оптимально сконструированные естественным отбором для выполнения своих функций. Ввиду недостатка места мы вынуждены опустить подробное обсуждение существенного вопроса «Что такое признак?». Некоторые эволюционисты могут посчитать это тривиальным иди даже исключительно семантической проблемой. Однако это не так. Организмы интегрированы, это не наборы дискретных объектов. Эволюционистов часто сбивает с верного пути неуместная атомизация. как любил указывать Д’Арси Томпсон (1942). Наш любимый пример связан с человеческим подбородком (Could, рр. 281-382; Lewontin. 1978). Если мы рассматриваем подбородок как самостоятельную «вещь», а не как продукт взаимодействия между двумя эмбриональными полями роста (альвеолярным и дентарным). то мы приходим к такой интерпретации его происхождения (рекапитуляционной12:), которая прямо противоположна наиболее популярной на данный момент (неотенической13).
2. После того, как рассмотрение оптимальности признаков по отдельности терпит неудачу, признается, что между признаками имеет место взаимодействие, в следующей формулировке: организм не может оптимизировать одну свою часть, не вызывая издержек в оптимизации другой. Вводится понятие эволюционного компромисса, и организмы интерпретируются как наилучшие компромиссы относительно противоречащих друг другу требований. Таким образом, рассмотрение взаимодействия между частями организма полностью остается в рамках адаптационистской программы. Любая субоптимальность какой-либо части объясняется се вкладом в наилучшую возможную конструкцию целого. Идея, что субоптимальность может указывать на что-либо отличное от непосредственной работы естественного отбора, обычно не одобряется. Как сказал доктор Панглосс, объясняя Кандиду, почему тот страдает от венерического заболевания:
«…это вещь неизбежная в лучшем из миров, необходимая составная часть целого; если бы Колумб не привез с одного из островов Америки болезни, заражающей источник размножения, часто даже мешающей ему и, очевидно, противной великой пели природы, — мы не имели бы ни шоколада, ни кошенили…».
Адаптационистская программа поистине панглоссова. Наш мир. возможно, нехорош с абстрактной точки зрения, но он лучший из возможных. Каждый признак играет свою роль и должен быть таким, какой он есть.
В этот момент некоторые эволюционисты начнут протестовать. что мы изображаем их взгляд на адаптацию в карикатурном виде. В конце концов, разве они не признают дрейф генов, аллометрию и множество других причин неадаптивной эволюции? Да. конечно. Но мы обращаем внимание на другое. В естественной истории возможно всякое. Обычно вы не утверждаете значимость вашего любимого феномена за счет объявления «конкурентов» несуществующими. Скорее, вы признаете существование конкурентов, но сужаете их сферу их действия настолько, что они не могут играть какой-либо важной роли в делах природы.
Затем вы можете похвалить себя за недогматичность и экуменизм. Мы отстаиваем точку зрения, что альтернативы отбору на наилучшую общую конструкцию часто низводятся до полного ничтожества именно при помощи такого способа аргументации. Не все ли мы слышали следующий догмат о дрейфе генов: он может быть важен только в популяциях настолько малых, что они с большой вероятностью вымирают, прежде чем он сыграет какую-либо продолжительную роль в эволюции (однако, см. (Lande, 1976)).
Признание альтернатив само по себе еще не предполагает их серьезного рассмотрения в повседневной практике. Мы декларируем. что не все адаптивно, однако, столкнувшись с организмом, мы склонны разбивать его на части и конструировать адаптивные сценарии так, как будто компромисс между соперничающими, хорошо оптимизированными частями является единственным, что отдаляет каждый признак от совершенства.
Это давняя традиция. Роменс жаловался на А.Р. Уоллеса в 1900 г.:
«Мистер Уоллес не утверждает в явном виде невозможность законов и причин, отличных от полезности и естественного отбора… Тем не менее, так какой нигде не признает какой-либо другой закон или причину, он практически делает вывод, что. с точки зрения индуктивных или эмпирических оснований, нет такого другого закона или при чины. который можно было бы рассматривать».
Адаптационистская программа прослеживается во многих распространенных способах аргументации. Мы приведем лишь несколько примеров: надеемся, что их узнают все.
1. Если один адаптивный аргумент не сработал, попробуйте другой. Зигзагообразный смык между створками раковин двустворчатых моллюсков и плеченогих. когда-то считавшийся приспособлением для укрепления раковины, превратился в сито, не пропускающее частицы, большие некоторого размера (Rudwick, 1964). Наружные образования (рога, клыки), когда-то считавшихся оружием против хищников, становятся символами внутривидовой конкуренции среди самцов (Davitashvili, 1961). Лицо эскимоса, ранее описанное как «сконструированное для защиты от холода» (Сооn et al., 1950). оказываются адаптацией для создания большой жевательной силы и приспособления к ней (Sheа, 1977). Мы не покушаемся на эти новые интерпретации: возможно, с ними все в порядке. Мы. однако, удивлены, почему неудача одного адаптивного объяснения всегда должна вдохновлять на поиск другого объяснения того же класса вместо рассмотрения альтернатив утверждению, что каждая часть сушестнует «для» некоторой специфической притоны.
2. Если один адаптивный аргумент не сработал, предположите, что должен существовать другой — ослабленная версия первого способа аргументации. Коста и Бизол (1978). например, надеялись найти корреляцию между генетическим полиморфизмом в популяциях и стабильностью среды в глубине океана, что им не удалось. Они делают вывод (рр. 132, 133):
«Обнаруженная степень генетического полиморфизма, по-видимому, указывает на отсутствие корреляции с какими-либо факторами внешней среды, характерными для участка, на котором проводили отбор образцов. Эти результаты позволяют предположить, что адаптивные стратегии организмов, принадлежащих к разным типам, различны».
3. В отсутствие хорошего адаптивного аргумента считайте причиной неудачи недостаточное понимание того, где организм живет, и чем он занимается. Это опять-таки старый довод. Рассмотрим. что пишет Уоллес о том. почему все детали окраски и формы наземных улиток должны быть адаптивными, даже если разные виды живут в одинаковой, вроде бы, среде (1899. Р.148):
«Точные пропорции различных видов растений, количества каждого вида насекомых или птиц, особенности, связанные с большей или меньшей подверженностью воздействию солнца или ветра в определенные критические периоды, и другие небольшие различия. которые для нас абсолютно нематериальны и неразличимы. могут иметь огромное значение для этих скромных созданий и быть вполне достаточными для небольшой корректировки размера. формы или цвета, которую вызывает естественный отбор».
4. Подчеркивайте непосредственную полезность и исключайте из рассмотрения другие атрибуты формы. Добрая половина пояснительного текста к стекловолоконной копии тираннозавра в Бостонском музее науки звучит так: «Загадка передних лап: каким образом Тyrannosaurus использовал свои крошечные передние лапы является научной загадкой, они были слишком короткими даже, чтобы дотянуться до головы. Возможно, они помогали животному вставать из лежачего положения». (Мы намеренно выбрали пример, связанный с влиянием науки на общество, чтобы показать. как широко распространяется стиль адаптационистской программы. Мы не используем стеклянных монстров в качестве мальчиков для битья: сходные доводы и соответствующие акценты регулярно встречаются и в профессиональной литературе).
Мы не сомневаемся, что тираннозавр использовал свои уменьшенные передние лапы для чего-либо. Если бы они возникли de novo, то мы бы поддержали поиск непосредственной причины адаптации. Но они являются, в конце концов, лишь результатом редукции гомологичных структур, которые у предков выполняли обычную функцию (длинные конечности аллозавров. например). По существу нам не требуется эксплицитно адаптивное объяснение их уменьшения. Возможно, это онтогенетический коррелят аллометрических полей относительного увеличения размера головы и задних конечностей. Эта неадаптивная гипотеза может быть проверена общепринятыми аллометрически ми методами (Gould, 1974. в общем: Lande, 1978. о редукции конечностей), и кажется нам как более интересной, так и более плодотворной, чем непроверяемые спекуляции, основанные на второстепенной полезности в лучшем из возможных миров.
Не следует путать факт, что структура используется каким-либо образом (обдумайте еще раз пазухи свода, пространства свода и тела ацтеков), с главной эволюционной причиной ее существования и устройства.
Сочинение историй
- Потому что. — говорил я». — если вулкан находится в Лиссабоне, то он и не может быть в другом месте: невозможно. чтобы что-то было не там, где должно быть, ибо все хорошо.(Доктор Панглосс о великом лиссабонском землетрясении 1755 года, в ходе которого расстались с жизнью до 50000 человек)
Мы бы не возражали столь энергично против адаптационистской программы, если бы обращение к ней в каком-либо конкретном случае могло хотя бы в принципе привести к ее отклонению в виду недостатка свидетельств. Тогда мы могли бы рассматривать ее как имеющую ограниченную область применения и возражали бы лишь против ее статуса как «нулевой гипотезы». Если бы ее можно было отвергнуть после неудачного прохождения какой-либо явной проверки, то альтернативы могли бы получить свой шанс. К сожалению. распространенная среди эволюционистов методика не допускает такого явного отвержения по двум причинам. Во-первых, отбрасывание одной адаптивной истории скорее приводит к ее замене на другую, чем к подозрению, что может потребоваться другой способ объяснения.
Так как диапазон адаптивных историй широк настолько, насколько богато наше воображение, всегда могут быть постулированы новые истории. А если такой истории пока нет, то всегда можно апеллировать к временному незнанию и полагаться на то, что она появится в будущем, как делали Коста и Бизол (1978), процитированные выше. Во-вторых, критерии для принятия истории так неопределенны, что многие истории могут быть приняты без надлежащего подтверждения. Часто эволюционисты используют согласованность с гипотезой естественного отбора как единственный критерий и считают свою работу выполненной, если удается состряпать правдоподобную историю. Однако правдоподобные истории можно придумать всегда. Ключ же к историческому исследованию состоит в создании критериев, позволяющих идентифицировать верное объяснение среди значительного количества правдоподобных путей, приводящих к современному состоянию.

Голубая сиалия Sialia currucoides, самец

Она же, самка
По этой причине мы. например, критиковали (Gоuld, 1978) работу Бараша (1976) по агрессии среди голубых сиалий. Бараш устанавливал чучело самца вблизи гнезд двух пар голубых сиалий в то время, когда самцы отсутствовали, добывая пищу. Он проделывал это у тех же самых гнезд три раза с десятидневными интервалами: первый раз—до откладки яиц, два последующих — после. Затем он подсчитывал чисто нападений возвращающегося самца на чучело и на самку. В первый раз агрессия была высокой по отношению к чучелу и низкой по отношению к самкам, но в целом — значительной для обоих гнезд. Агрессия по отношению к чучелу постепенно уменьшалась во второй и в третий раз и упала до нуля по отношению к самкам. Бараш сделал вывод, что в этом есть эволюционный смысл, так как самцы должны быть более чувствительны к вторжению чужаков до откладки яиц, а не после (когда у самцов уже есть некоторая уверенность, что они передали свои гены потомству). Создав правдоподобную историю, он посчитал свою работу завершенной (1976. рр. 1059-1100).
Эти результаты соответствуют ожиданиям эволюционной теории. Таким образом, агрессия по отношению к вторгшемуся самцу (чучелу). очевидно, была бы особенно выгодна в начале сезона размножения, когда обычно и защищают территории и гнезда. Первоначальная агрессивная реакция по отношению к самке из той же пары также адаптивна, так как, учитывая, что в данной ситуации высока вероятность «измены» (а именно, чучело находится рядом с самкой) и в предположении, что доступны другие самки, приобретение нового полового партнера увеличит приспособленность самцов. Уменьшение агрессии самцов по отношению к самкам в период высиживания яиц и на стадии оперения птенцов может быть приписано невозможности измены после того, как яйца были отложены. Эти результаты соответствуют эволюционной интерпретации.
Они действительно соответствуют, но что насчет очевидной альтернативы, отброшенной без проверки Барашем? Самец возвращается во второй и третий раз, приближается к чучелу, немного тестирует его, узнает в нем ту же подделку, которую видел и раньше, и не беспокоит свою самку. Почему хотя бы не провести очевидную проверку этой альтернативы обычной адаптивной истории: предложить чучело самцу в первый раз лишь после откладки яиц?
После нашей критики исследования Бараша Мортон с сотрудниками (1978) повторили его с некоторыми изменениями (включая введение чучела самки), для близкородственного вида — восточной сиалии Sialia sialis. «Мы надеялись подтвердить». — пишут они, — что выводы Бараша указывают на «распространенное эволюционное явление. по крайней мере внутри рода Sialiа. К сожалению. нам это не удалось».

Восточная сиалия Sialia sialis, самец и самка
Они не обнаружили никаких признаков «ревнивого» поведения: самцы никогда не нападали на своих самок после встречи с чучелом, вне зависимости от стадии гнездования. Вместо того самки часто приближались к чучелу самца и в любом случае атаковали чучела самок чаще, чем чучела самцов. «Эта яростная реакция привела к почти полному разрушению чучела самки после двух его предъявлений и к его гибели во время третьего, когда самка скрылась в кустах с головой чучела в самом начале эксперимента» 11978. р. 969). Однако, вместо того чтобы поставить под сомнение сценарий Бараша, исследователи лишь изобрел и свой собственный, интерпретируя результаты обоих экспериментов в адаптационистском ключе. Возможно, предполагают они, «самки на замену» были редкими для их вида и были в изобилии для вида, исследованного Барашем.
Так как в работе Бараша самцы могли заменить потенциально «неверную» самку, они могли позволить себе быть разборчивыми и больше собственниками ми. Самцы восточной сиалии стараются сохранить редких партнеров и должны проявлять к ним уважение. Исследователи заключают: «Хотя мы и не поддерживаем предположение Бараша о том. что самцы голубой сиалии демонстрируют адаптации против «измены». мы предполагаем, что все же в обоих работах «результаты соответствуют ожиданиям эволюционной теории» (Ваrash, 1976, р. 1099). что и должно иметь место, как мы предполагаем, в случае любого тщательного исследования». Но что хорошего в теории, которая не может оказаться ошибочной ни при каком тщательном исследовании (ведь под «эволюционной теорией» они. очевидно, подразумевают действие естественного отбора в конкретных случаях. а не сам факт превращения видов одного в другой)?
Перелистывая Мастера
Так как Дарвин приобрел ореол святости, если не божественности, среди эволюционных биологов, и так как все стороны декларируют правоверность, часто считался, что Дарвин в глубине души был радикальным селекционистом, ссылавшимся на другие механизмы эволюции только в качестве уступки, и лишь из-за свойственного его времени прискорбного незнания механизмов наследственности. Этот взгляд неверен. Хотя Дарвин рассматривал отбор как наиболее важный из эволюционных механизмов (как считаем и мы), ни один из доводов оппонентов не раздражал его более, чем распространенная попытка окарикатурить и тривиализировать его теорию, утверждая, что она основана исключительно на естественном отборе. В последнем падании «Происхождения видов» он писал14 (1872. р. 395):
«Но так как в недавнее время мои вывозы были превратна истолкованы, и утверждали, что я приписываю модифицирование видов исключительно естественному отбору, то мне, может быть, позволено будет заметить, что в первом и последующих изданиях этой книги я поместил на очень видном месте, именно в конце «Введения», следующие слава: «Я убежден, что естественный отбор был главным, но не исключительным фактором модификации». Но это не помогло. Велика сила упорного извращения…
Ромене, чье некогда знаменитое эссе о плюрализме Дарвина в сравнении с панселекционизмом Уоллеса и Вейсмана (1900) заслуживает воскрешения. заметил о приведенном отрывке (1900. р. 5): «Среди всех работ Дарвина не найти пассажа так решительно сформулированного как этот: это единственная нотка горечи среди тысяч страниц, которые он опубликовал». Очевидно, Ромене не знал о письме, которое Дарвин написал в Nature в 1880 году, в котором он осуждает сэра Уайвилла Томсона за карикатурно панселекционистское изображение своей теории (1880. р.32):
«Жалко обнаружить, что сэр Уайвилл Томсон не понимает принцип естественного отбора… Если бы он его понимал, то не мог бы написать следующее предложение во введении к «Плаванию «Челленджера»: «Характер глубоководной фауны ни в малейшей степени не поддерживает теорию, которая рассматривает эволюцию видов вплоть до самых крайних вариантов как направляемую лишь естественным отбором». Эта стандартная критика нередко встречается среди теологов и метафизиков, когда они пишут на научные темы, но когда она исходит от натуралиста, это что-то новое… Может ли сэр Уайвилл Томсон назвать хоть кого-нибудь, кто бы утверждал, что эволюция видов зависит только от естественного отбора? Что касается меня, я полагаю, что никто не представ ил такое количество наблюдений, касающихся использования и неиспользования частей организма, как это сделал я в работе «Изменения животных и растений при одомашнивании»: и эти наблюдения были сделаны именно для этой цели. Я также привел там значительное количество фактов, демонстрирующих прямое действие условий внешней среды не организмы».
Сегодня мы не считаем существенными или хотя бы рабочими все вспомогательные эволюционные механизмы по Дарвину, хотя многие из них, включая непосредственную модификацию15 и корреляции роста, очень важны. Но мы должны с воодушевлением отнестись к его последовательно плюралистической позиции и в отношении объяснения сложности Природы.
Неполная типология альтернатив адаптационистской программе
В духе дарвиновского плюрализма мы представим неполную иерархию альтернатив объяснению формы, функции и поведения через непосредственную адаптацию.
I. Возможно полное отсутствие адаптации и отбора. В настоящий момент мнения популяционных генетиков резко разделились по вопросу, насколько генетический полиморфизм внутри популяций и генетические различия между видами являются, действительно, результатом действия естественного отбора, а не полностью случайных факторов. Размер популяций конечен, и изолированные популяции, которые являются первой стадией процесса видообразования, часто основываются очень маленьким чистом особей. В результате такого ограничения на размер популяции частоты аллелей изменяются за счет дрейфа генов,. чего-то вроде случайной ошибки генетической выборки.
Стохастический процесс изменения частоты гена за счет случайного дрейфа генов, включая ярко выраженный процесс «взятия выборок» в случае формирования новой изолированной популяции из нескольких иммигрантов, приводит к важным последствиям. Во-первых, возникают генетические различия между популяциями и видами, вплоть до фиксации разных аллелей по одному и тому же локусу, при полном отсутствии какого-либо давления естественного отбора.
Во-вторых, аллели могут быть зафиксированы в популяции несмотря на естественный отбор. Лаже если естественный отбор благоприятствует некоторому аллелю, определенная доля популяций. пропорциональная произведению размера популяции N на коэффициент селекции s, станет из-за дрейфа генов гомозиготной по аллелю с меньшей приспособленностью. Если N*s велико, то такая случайная фиксация неблагоприятных аллелей будет редким явлением, но если коэффициент селекции оказывается порядка величины, обратной размеру популяции (N*s=1) или меньше, то фиксация вредных аллелей оказывается обычным явлением.
В том случае, когда количественный признак, связанный, например, с формой, метаболизмом или поведением, определяется действием множества генов, то интенсивность отбора по каждому из локусов будет мала и значение N*s на локус также будет мало. В результате по многим локусам произойдет фиксация неоптимальных аллелей.
В-третьих, для новых мутаций вероятность удержаться в популяции не велика, даже если они поддерживаются отбором. Дрейф генов вызывает немедленную потерю большинства новых мутаций после их появления в популяции. При интенсивности селекции s вероятность удержания новых мутаций — всего лишь 2s. Поэтому нельзя утверждать, что в конечном счете мутация, подходящая для некоторого объяснения через адаптацию, возникнет и распространится в популяции. «В конечном счете» оказывается очень длинным промежутком времени, даже если одна из 1000 или одна из 10000 «подходящих» мутаций, появляющихся в популяции, когда-либо распространится в ней.
II. Дело может быть не в адаптации и не в действии отбора на данную часть организма: форма части может являться следствием корреляции с последствиями отбора, действующего на что-то другое. Под эту важную категорию подходят дарвиновские «загадочные» законы «корреляции роста». Сегодня же мы говорим о плейотропии, аллометрии, «материальной компенсации» (Rensch, 1959. р. 179-187) и механически вынужденных корреляциях в смысле Д’Арси Томсона (1942: Gould, 1971). Здесь мы вплотную сталкиваемся с организмами как интегрированными целыми, фундаментально не разложимыми на независимые и по отдельности оптимизируемые части.
Хотя аллометрические паттерны16 также подвержены отбору, как и сама статическая морфология (Gould, 1966), некоторые закономерности относительного роста, вероятно, не имеют непосредственного отношения к адаптации. Например, мы не ставим под сомнение, что знаменитая межвидовая аллометрия размера мозга во всех главных группах позвоночных с коэффициентом 0,66 представляет собой отобранное «инженерное решение», хотя уловить ее значение по-прежнему не удается (Erlson, 1973). Она встречается слишком часто и в слишком широком таксономическом диапазоне, чтобы указывать на нечто иное, чем на ряды одинаково хорошо сконструированных организмов разных размеров[1]. Но другая распространенная аллометрия, внутривидовая или среди внутривидовых рас, с коэффициентом от 0,2 до 0,4, среди взрослых гомойотермных животных разных размеров, вероятно, не требует селекционистской интерпретации, хотя многие, включая одного из авторов, пытались доказать одну из них (Gould, 1974). Р.Ланде (личное сообщение) использовал эксперименты Фальконера (Falcоner, 1973). чтобы показать, что отбора на изменение размера тела у мышей достаточно для появления аллометрии размера мозга с коэффициентом 0.35.
Более убедительные примеры изобилуют в литературе, посвященной отбору на сдвиг сроков полового созревания (Gould, 1977). По крайней мере три раза в ходе эволюции членистоногих (в случае клещей, мух и жуков) возникала одна и та же сложная адаптация, очевидно, связанная с приспособлением к быстрой смене поколений у организмов, использующих для питания сверхобильные, но эфемерные ресурсы грибного происхождения и подвергающихся сильному давлению отбора в сторону г-стратегии17: самки размножаются на личиночной стадии и выращивают следующее поколение внутри своих тел. Потомство поедает материнскую особь изнутри и вылупляется из ее пустой оболочки лишь для того, чтобы через несколько дней оказаться пожранным собственными отпрысками.
Было бы глупо искать адаптивное значение в педоморфической18 морфологии самой по себе: это в первую очередь побочный результат отбора на быструю смену поколений. В более интересных случаях отбор на уменьшение размера тела (карликовые самцы многих ракообразных) приводит к прогенезу19 (Gould, 1977, рр. 324-336), в результате взрослые потомки обладают смесью ювенильных и дефинитивных признаков своих предков. Многие биологи поддались искушению считать первичным адаптивное значение этого смешения, но оно. вероятно, возникло как побочное следствие усеченного созревай (и. оставившего некоторые признаки из личиночной стадии и в то же время позволившего другим признакам, сильнее корреллирующим с половым созреванием, сохраниться такими же. как у взрослых предков.
III. Отбор и адаптация могут быть разобщены.
(1). Отбор без адаптации. Левонтин (Lewontin, 1979) приводит следующий гипотетический пример:
«Мутация, увеличивающая плодовитость особей в два раза, быстро распространится в популяции. Если при этой не изменится эффективность использования ресурсов, то особи не будут производить потомства больше, чем раньше, а просто будут откладывать в два раза больше яиц, а избыточное потомство будет погибать из-за недостатка ресурсов. В каком смысле особи в популяции будут адаптированы лучше, чем раньше? В самом деле, если хищник, специализирующийся на не- зрелых особях, переключится на этот вид, в котором из-за мутации незрелых особей стало больше, то в результате размер популяции может даже уменьшиться, но естественный отбор в любом случае будет благоприятствовать особям с наибольшей плодовитостью».
(2) Адаптация без отбора. Многие прикрепленные морские организмы, губки и кораллы в особенности, хорошо адаптированы к режиму тока воды, в котором они живут. Разнообразные «хорошие конструкции» такого рода могут иметь исключительно фенотипическое происхождение, будучи индуцированными самим течением. (Мы с уверенностью можем говорить об этом в многочисленных случаях, когда генетически идентичные особи колонии принимают различную форму в разных микросредах). Паттерны географической изменчивости также часто являются адаптивными. нов тоже время исключительно фенотипическими.
Суини и Ванноте (Sweeny, Wannote, 1978), например, показали, что многие водные насекомые с неполным превращением обладают меньшим размером во взрослом состоянии и меньшей плодовитостью, когда их выращивают при температурах ниже или выше оптимальной. В соответствии с этим коррелирующие с климатом паттерны географической изменчивости для этих насекомых, так часто а рriori принимаемые за признак адаптации на генетическом уровне, могут просто отражать их фенотипическую пластичность.
«Адаптация» — хорошая приспособленность организмов к их среде— может достигаться на трех уровнях организации, под действием разных причин. К несчастью, наш язык сосредоточен на общем для этих случаев результате и называет все три явления «адаптацией» — различие процессов маскируется, что часто подталкивает эволюционистов к распространению дарвиновской логики и на два других уровня.
Во-первых, есть то. что называют «адаптацией» физиологи: фенотипическая пластичность, которая позволяет организмам приспосабливать свою форму под разные обстоятельства онтогенеза. «Адаптации» человека к высокогорным условиям попадают в эту категорию (несмотря на то, что другие адаптации человека, такие как устойчивость больных серповидно-клеточной анемией к малярии, могут быть генетическими и дарвиновскими). Физиологические адаптации не наследуются, в то время как способность к их выработке, по умолчанию, наследуется. Во-вторых, есть «наследуемая» форма недарвиновской адаптации у человека (и. в зачаточном виде, у некоторых других высокоорганизованных социальных животных): культурная адаптация, наследуемая благодаря обучению).
Значительная доля путаницы в социобиологии человека возникает из-за неспособности различить этот режим адаптации и дарвиновскую адаптацию, основанную на генетической изменчивости. Наконец, есть адаптация, возникающая в результате обычного дарвиновского механизма отбора, использующего генетическую изменчивость. Хорошей приспособленности организма к среде самой по себе недостаточно для вывода о том. что в данном случае работает естественный отбор.
IV. Если имеет место и адаптация, и отбор, адаптации при этом могут быть не различимы с точки зрения отбора. У близкородственных видов или у популяций внутри видов часто возникают разные адаптации для решения одной и той же проблемы. Когда занято несколько «адаптивных пиков», у нас обычно нет оснований предполагать, что одно из решений лучше другого. Какое решение будет выбрано в какой-либо ситуации, определяется исторически: тем, что первые шаги были в определенном направлении, хотя другие направления могли бы привести к не меньшему процветаний.
Каждый натуралист может привести свой любимый пример. Например, у наземной улитки Сеrion c островов Вест-Индии популяции, живущие на скалистых и обдуваемых ветром берегах по понятным причинам, связанным с адаптацией, почти всегда образуют белые, толстостенные и относительно «приземистые» раковины. В онтогенезе улиток рода Сеrion можно различить по крайней мере два разных способа приобретения белой окраски за счет линьки ранних завитков, два способа утолщения стенок раковин и три вида аллометрии, приводящие к «приземистости». Все двенадцать возможных комбинаций встречаются в популяциях Багамских островов, но плодотворно ли будет рассматривать вопрос, почему Сеriоп с расположенного восточнее острова Лонг-Айленд эволюционировали в сторону одного решения, а Сеrion с острова Аклинс — в другую, с точки зрения оптимального дизайна, а не с точки зрения исторической контингентноcти:
«.Если имеет место и адаптация, и отбор, адаптация при этом может представлять вторичную утилизацию частей организма, присутствующих в нем по архитектурным, онтогенетическим или историческим причинам. Мы уже обсудили эту игнорируемую тему в первом разделе, на примерах пазух свода, свободных пространств свода и каннибализма. Если румянец оказывается адаптацией, возникшей у человека под действием полового отбора, это не помогает нам понять, почему кровь является красной. Непосредственная полезность органической структуры часто ничего не говорит о причинах существования этой структуры.

Cerion chrysalis
Ещё один оклеветанный подход к изучению эволюции
Для эволюционистов из континентальной Европы никогда особенно не была характерна англо-американская склонность «атомизировать» организм на части и пытаться объяснить каждую как непосредственную адаптацию. Их главный альтернативный подход существует в сильной и в слабой формулировке. В сильной формулировке, которую отстаивают такие ведущие теоретики. как Шиндевольф (Schindewolf, 1950). Ремане (Remane, 1971) и Грассе (Grasse, 1977), естественный отбор может объяснить лишь поверхностные модификации плана строения (Вaирlaп). которые приспосабливают строение организма к особенностям среды: почему, например, кроты слепые, у жирафов длинная шея, а у уток перепончатые лапы. Но важные этапы эволюции, такие как возникновение самого плана строения и переход между разными планами строения, должны происходить с участием какого-то другого неизвестного и. возможно, «внутреннего» механизма. Мы полагаем, что английские биологи правильно отвергают эту сильную формулировку, граничащую с мистицизмом.
Но у данного аргумента есть и слабая, парадоксальным образом оказывающаяся мощной, формулировка, которая не была оценена по достоинству, но заслуживает этого. Она так же признает, что обычный естественный отбор приводит к поверхностным модификациям плана строения. И она так же отрицает, что адаптационистская программа (атомизация плюс оптимизация отдельных частей под действием отбора) способна на многое в объяснении планов строения и переходов между ними.
Но она не обращается вследствие этого к фундаментально неизвестному процессу. Вместо того она утверждает, что базовые планы строения организмов так интегрированы и так насыщены ограничениями на возможность адаптации (категории 2 и 5 в нашей типологии), что традиционные аргументы от естественного отбора мало что могут объяснить относительно них. Слабая формулировка не отрицает, что изменение. когда оно происходит, может быть опосредовано естественным отбором, но она утверждает, что налагаемые связи настолько ограничивают пути и способы изменения, что эти ограничения сами по себе оказываются гораздо более интересным аспектом эволюции.
Руперт Ридль. австрийский зоолог, попытавшийся развить этот тезис для англоязычной аудитории (Riedle, 1977. 1975. 1978), пишет:
“Так оказалась, что живой мир полон универсальных схем организации, которые, совершенно очевидно, не могут быть напрямую объяснены условиями окружающей среды или адаптивной радиацией20, но существуют в первую очередь из-за универсальных системных требований к сложной организации как таковой… Это не самоочевидно, так как весь глубокий и обширный идейный запас, накопленный морфо- логией от Гете до Ремане, буквально отрезан от современной биологии. Его не изучают в большинстве американских университетов. Исчезли даже преподаватели, которые могли бы его преподавать».
Ограничения на возможность эволюционных изменений могут быть подразделены по крайней мере на две категории. Все эволюционисты знакомы с филогенетическими21 ограничениями, которые можно проиллюстрировать проведенным Грегори классическим различением между внешним обликом и наследством, доставшимся от предков (Gregory, 1936). Мы допускаем что-то вроде филогенетической инерции, когда признаем, например, что люди не идеально приспособлены для прямохождения, потому что многое в нашем плане строения возникло для жизни на четырех конечностях. Мы также привлекаем филогенетические ограничены для объяснения, почему нет летающих моллюсков или почему не бывает насекомых размером со слона.
Онтогенетические22 ограничения — разновидность филогенетических ограничений — возможно, наиболее могущественное из всего, что ограничивает пути эволюции. В сложных организмах ранние стадии онтогенеза замечательно устойчивы к эволюционным изменениям — предположительно из-за того, что дифференциация систем органов и их интеграция в функционирующий организм — очень хрупкий процесс, легко «сходящий с рельсов» в случае ранних ошибок. эффекты которых затем накапливаются.
Фундаментальные эмбриологические законы фон Бэра (von Ваег. 1828) означают немногим большее, чем признание того, что ранние стадии развития одновременно и очень консервативны, и сильно ограничивают возможности дальнейшего развития. Биогенетический закон Геккеля, главная тема эволюционной биологии в конце XIX века, основана на неправильной интерпретации этих же данных (Gould, 1977). Если развитие осуществляется интегрированными модулями и не может разбираться по кусочкам в ходе эволюции, то адаптационистская программа не может объяснить перестройку программ развития, лежащую в основе почти всех изменений плана строения.
Немецкий палеонтолог А. Зейлахер, чья работа заслуживает гораздо больше внимания, чем она получила, подчеркивает то, что он называет «строительно-техническими23, или архитектурными, ограничениями» (Sellacher, 1970). Они возникают не из бывших адаптаций, сохранившихся в новом экологическом окружении (как обычно понимают филогенетические ограничения), но как архитектурные ограничения, которые никогда небыли адаптациями, но, скорее, были необходимыми следствиями выбора материалов и конструкционных решений при создании базового плана строения. Мы посвятили первый раздел этой работы небиологическим примерам из этой категории. Пазухи свода собора возникают с необходимостью, как только чертеж предписывает, что свод должен покоиться на округлых арках. Архитектурные ограничения могут оказывать далеко идущее влияние и на организмы. Эта тема скрывает множество возможных прозрений, так как даже её существование редко признается.
Зейлахер рассматривает очаровательный пример (Sellacher, 1972). Диварикатная скульптура24 раковины (рис.3) возникает снова и снова во всех группах моллюсков и у плеченогих.
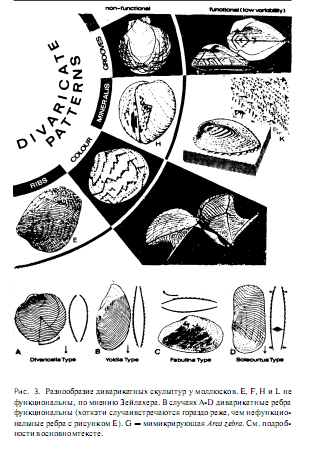
Эта базовая форма может проявляться в разнообразных структурах: рельефных ребрах (не совпадающих со следами остановки роста, так как они никогда не согласуются с краем мантии), характере окраски, внутренних структурах в минерализованном кальцитовом слое раковины и диварикатных желобках на поверхности раковины. Зейлахер не знает, за счет чего возникает диварикатный паттерн, но он убежден, что традиционная концентрация внимания почти исключительно на адаптивном значении каждого его проявления отвлекает от вопроса о возникновении этих проявлений в процессе роста и также препятствует признанию универсальности этого паттерна. Этот феномен должен возникать из-за некоторой характерной негомогенности в растущей мантии, вероятно, из-за интерференции между регулярно расположенными центрами роста: простая компьютерная симуляция может привести к возникновению той же формы таким способом (Waddington, Cowe, 1969). Диварикатный паттерн в целом может вовсе не являться непосредственной адаптацией.
Зеллахер затем утверждает, что большинство проявлений данного паттерна, вероятно, неадаптивны. Его доводы разнообразны, но нам кажутся в целом разумными. Некоторые из них основаны на данных полевых наблюдений: характер окраски оказывается невидимым, так как двустворчатые моллюски, обладающие им. либо живут, зарывшись в грунте, либо их раковины покрыты наружным роговым слоем такой толщины, что цвета под ним неразличимы. Другие основаны на более общих принципах: некоторые проявления встречаются только у необычных и патологических особей, или встречаются редко и в виде аномалий развития, или демонстрируют избыточную изменчивость по сравнению с гораздо меньшей изменчивостью в тех случаях, когда в целом та же самая структура принимает форму, выглядящую функциональной с инженерной точки зрения.
Для каждой из четырех перечисленных категорий диварикатная скульптура раковины оказывается функциональной в явном меньшинстве случаев (рис.3). Диварикатные ребра могут служить в качестве ковшей и якорей при закапывании (Stanley, 1970), но у большинства двустворчатых моллюсков они не расположены подходящим образом. Окрашенные шевроны выполняют функцию мимикрии у одного вида (Pteria zebra), который живет на ветвях колоний гидроидных полипов, в этом случае их изменчивость сильно уменьшена. Минерализованные шевроны адаптивны, вероятно. только у одного замечательного создания, необычного двустворчатого моллюска Сorculum cardissa (у других видов они либо встречаются у особей с отклонения ми. либо возникают в результате посмертной эрозии раковины). Для этого моллюска характерно уникальное уплощение в передне-заднем направлении. Он лежит на субстрате, задним краем вверх. Вокруг заднего конца располагаются минерализованные диварикатные треугольники. Они полупрозрачны, в то время как оставшаяся часть раковины непрозрачна. Под этими «окошками» живут эндосимбиотические водоросли!

Pteria zebra

Сorculum cardissa
Все предыдущие работы по диварикатной скульптуре сосредоточивались на ее адаптивном значении (и в большинстве случаев не могли ее обнаружить). Однако Зейлахер, вероятно, прав, расcматривая этот случай так же, как мы рассматривали пазухи свода, свободные места на потолке и человеческие жертвоприношения в первом разделе. Диварикатная скульптура раковины является фундаментальным архитектурным ограничением. Время от времени, раз уж она присутствует, она может быть с выгодой использована. Но мы не сможем понять эту структуру или ее эволюционный смысл, если будем рассматривать эти нечастые и вторичные адаптации как причину самой структуры.
Гальтон вы разил разницу между адаптационистской программой и установкой на изучение ограничений и режимов развития, сославшись на выразительную историю об отпечатках пальцев Герберта Спенсера (Galton, 1909. р. 257):
«Многое было написано, хотя последнее слово пока еще и не сказано, о разумном объяснении этих любопытных папиллярных линий: почему у одних людей и на одних пальцах они образуют спирали, а на других — петли. В связи с этим я могу упомянуть характерную историю, связанную с Гербертом Спенсером. Он попросил показать ему мою лабораторию и взять отпечатки его пальцев, что я и сделал. Затем я заговорил о неспособности выяснить происхождение этих рисунков, потом, как препарируют пальцы еще не рожденных детей, чтобы прояснить ранние стадии их возникновения, и так далее. Спенсер заметил, что этот подход неверен изначально, и что я должен рассмотреть цель, которой служат эти линии, и проделать работу в обратном направлении. В данном случае, сказал он, очевидно, что нежные устья потовых желез требуют зашиты, которую им и предоставляют кожные гребешки по обе стороны от них, и исходя из этого он а больших подробностях развил непротиворечивую и хитроумную гипотезу. Я ответил, что его доводы прекрасны и заслуживают быть истинными, но так сложилась, что устья протоков располагаются не а бороздках между гребешками, а на вершине самих гребешков».
Мы убеждены, что потенциальные выгоды от отказа сосредоточиваться исключительно на адаптационистской программе действительно велики. Это не жест отчаяния, в чем нас обвиняют адаптационисты. так как «неадаптивный» не значит «непостижимый». Мы приветствуем то богатство, которое может предоставить плюралистический подход, столь близкий духу Дарвина. В период господства адаптационистской программы великие исторические темы морфологии развития были почти полностью заброшены, так как если отбор может разрушать любую корреляцию и оптимизировать части по отдельности, тогда интегрированность организма немногого стоит. Слишком часто адаптаиионистская программа предлагает нам эволюционную биологию частей организма и генов, но не организмов. Считается, что все переходы могут осуществляться пошагово и игнорируется важность интегрированных модулей, составляющих формирующийся организм, и всюду проникающих исторических и архитектурных ограничений. Плюралистический взгляд мог бы вернуть организмы со всей их непокорной, но постижимой сложностью назад в эволюционную теорию.
Литература



Примечания переводчика
1. Высказывались соображения, что вместо термина «spandrels» (пазухи свода или сводов) в названы и статьи следовало использовать «pendentives» (паруса свода или сводов), на что Гулд указывает, что «spandrels» является более обoим понятием, которое легче экстраполировать на биологические примеры. См. Gould S.J. The exaptive excellence of spandrels as term and prototype// PNAS. 1997. V.94. №.20. С.10750-10755). Также заметим, что а случае данной статьи верно переводить «spandrels» как «пазухи свода», а не «пазухи сводов», та к как речь идет не о всех сводах собора Святого Марка, а лишь о центральном.
2. Гулд и Левонтин в качестве синонимов используют термины «адаптационистский», «панадаптационистский» и «селекционистский».
3. В оригинале «trade-off». Данный термин обозначает невозможность одновременной оптимизации по нескольким параметрам. Общепринятого варианта перевода нет, здесь и далее термин, переводится как «компромисс» или «эволюционный компромисс», в «биологической литературе встречается также «трейд-офф» или «трейдофф».
4. Фиксация аллеля (варианта гена) — достижение им в популяции частоты 100%. Случайная фиксация аллелей происходит под действием дрейфа генов.
5. Аллометрия — неравномерный рост частей тела, часто описывается степенным законом и в таком случае характеризуется степенным коэффициентом.
6. Плейотропия — множественное действие гена.
7. Материальная компенсация — отрицательная корреляция между частями организма а связи с конкуренцией между ними за ресурсы при их одновременном или взаимосвязанном росте. См. West-Eberhard M.J. Developmental plasticity and evolution. Oxford Univ. Press., 2003.
8. Здесь и далее цитирование по изданию Вольтер Ф.-М. А. Избранные сочинения. Пер. с фр. М.. «РИПОЛ КЛАССИК». 1997.
9. Ортогенез — концепция, предполагающая обусловленность эволюции внутренними причинами, направляющими ее по определенному маршруту. В рамках данной концепции допускается «инадаптивная эволюция», при которой заданное направление эволюции противоречит требованиям адаптации, что в итоге приводит к вымиранию.
10. В оригинале — «design». По отношению к организмам далее в тексте переводится также как «дизайн», а в биологической литературе встречаются и другие варианты, например «замысел» («intelligent design» — «разумный замысел»).
11. Филогенетическая инерция — паттерны (см. примечание 16) эволюции, указывающие на ее направленный характер. Филогенетическую инерцию трактовали по-разному: как свидетельство ортогенеза, как результат естественного отбора или как фундаментальные свойства популяций или организмов, влияющие на степень и скорость эволюционных изменений под действием естественного отбора. См. Shanahan T. Phylogenetic inertia and Darwin’s higher low// Studies in History & Philosophy of Science. Part C: Studies in History & Philosophy of Biological and Biomedical Science. V.42. №1. Р.60-68.
12. Рекапитуляция — повторение в индивидуальном развитии признаков, свойственных более ранней стадии эволюции.
13. Неотения — достижение половозрелости и окончание онтогенеза на более ранних стадиях индивидуального развития, чем у предкоаых организмов.
14. Цитата приведена по изданию: Дарвин Ч. Происхождение видов путем естественного отбора. СПб.. Наука. 2001.
15. Подразумевается наследование приобретенных признаков «по Ламарку».
16. «Pattern»— распространенный термин, не переводимый однозначно на русский язык и означающий закономерную регулярность в данных, повторяющийся шаблон, образец. В данном тексте используются варианты перевода «паттерн» и «схема».
17. r-стратегия — экологическая стратегия размножения, состояния в стремлении к максимально возможной скорости роста численности популяции и приводящая к гибели значительной доли потомства до достижения им зрелого возраста.
18. Педоморфоз — путь и механизм эволюционных преобразований, при котором во взрослой морфологии потомков в качестве дефинитивных («взрослых») сохраняются ювенильные или личиночные признаки предковой морфологии.
19. Различают две формы педоморфоза: неотению (замедление соматического развития) и прогенез (ускорение полового развития).
20. Адаптивная радиация — образование разнообразных форм организмов в пределах вида или группы родственных видов в результате приобретения организмами новых адаптаций и проникновения в новые адаптивные зоны.
21. В оригинале — «phyletic».
22. В оригинале— «developmental».
23. В оригинале — «bautechnischer» (нем.).
24. Скульптура раковины моллюска — паттерн неровностей наружной поверхности раковины, представленных килями, ребрами, ребрышками, гребнями и пластиками. Диварикаткая скульптура — скульптура в виде косой или зигзагообразной струйчатосги или ребристости элементов внутреннего строения раковины.
Примечание
[1]тогда как отклонение от него в пользу значимо больших показателей степени в филогенетической линии приматов показывает эволюционный тренд на ускоренное развитие мозга, а в пользу значимо меньших коэффициентов в линии ушастых тюленей – на «отставание» увеличения мозга от размеров тела, связанного, видимо, с конкуренцией между самцами на лежбищах.
Рекомендуем прочесть
 Долговые тюрьмы убивают бедных американцев
Долговые тюрьмы убивают бедных американцев Правозащитникам
Правозащитникам Норвегия. Результаты 5-летней работы законодательства, криминализующего клиента проституток.
Норвегия. Результаты 5-летней работы законодательства, криминализующего клиента проституток. Коллективизм и здоровье: эффект Розетто
Коллективизм и здоровье: эффект Розетто Про акмэ идеи «естественности неравенства полов»
Про акмэ идеи «естественности неравенства полов» Кризис гегемонии ПСР
Кризис гегемонии ПСР Нейроны фон Экономо
Нейроны фон Экономо Предшественники «русских фантастов»
Предшественники «русских фантастов»
