
Я когда-то писал, чем не нравится выражение «гены шизофрении» (диабета, алкоголизма и других состояний или форм поведения, в которых есть значимая наследственная компонента). Гены в онтогенезе в отношении к признакам, продуцируемым в ходе этого онтогенеза – это как рабочие на конвейере, собирающие машины, заменимые, контролируемые и подгоняемые, но никак не часть этой машины, и не нечто вроде средневековых ремесленников, делающих своё изделие самостоятельно. См. про диабет в книге Грега Гибсона «Во всём виноват геном»
Эпидемия генетического заболевания
Содержание
В случае диабета второго типа ситуация совершенно другая. Это эпидемическое заболевание. За 30 лет заболеваемость выросла от нескольких процентов до десяти процентов и более и продолжает расти с угрожающей скоростью. Одни только гены не могут вызвать эпидемию, здесь обязательно должен быть какой-то внешний фактор. И каждый из нас знает, что этот фактор — переход на систему быстрого питания и сидячий образ жизни. Гены здесь лишь невольные соучастники. В этой опасной игре они являются невинными жертвами тех изменений, которые произвел сам человек, нечаянно вовлеченными в болезнь, к которой не имеют никакого отношения.
Мы сколь угодно долго можем говорить о гипергликемии или устойчивости к инсулину, но основная проблема при диабете второго типа заключается в нарушении регуляции процессов метаболизма. Постоянно высокое содержание сахара в нашем рационе питания приводит к увеличению выработки инсулина выше того уровня, который организм может нормально переносить. В конечном итоге организм не выдерживает и прекращает реагировать на гормон. Современный стиль жизни довел тонко организованный механизм проверок и равновесий до границ его буферной способности. Те люди, кому не повезло и кто ввиду своих генетических особенностей обладает более слабой буферной системой, имеет более высокий риск развития диабета.
Итак, гены участвуют в развитии диабета, но скорее в качестве пособников, чем действующей силы. Если мы хотим понять их роль в этом процессе, мы должны рассмотреть три вопроса. Во-первых, почему некоторые из нас более склонны к перееданию, чем другие? Во-вторых, почему переедание у одних людей ведет к полноте, а у других нет? В-третьих, какова связь между увеличением массы тела и диабетом второго типа, и существуют ли гены, участвующие в развитии заболевания независимо от массы тела человека?
Связать нарушение метаболизма с увеличением массы тела несложно. Со временем происходит дисбаланс между потреблением калорий и их расходованием. Сорокалетний мужчина с избытком массы тела в 20 кг, что сегодня встречается достаточно часто, с момента окончания школы прибавлял в весе в среднем по полкило в год. В неделю это составляет примерно 10 г. Вопрос: сколько граммов сахара содержится в одном стакане кока-колы? Ответ: 39. Теоретически откажите себе в одном лишнем пиве или пончике на еженедельной вечеринке — и проблема решена.
Каждый из нас, погружая руку в кружку с леденцами, стоящую на столе у коллеги по работе, думает, что эти калории он израсходует на обратном пути в собственный офис. Европейцы, по крайней мере, каждое утро идут пешком до трамвая, а не до гаража. Если съеденные калории напрямую связаны с объемом талии, кажется вполне разумным, что полтора часа физических упражнений в неделю могут помочь слегка сбавить вес.
Действительно, в развитии диабета важную роль играют некоторые другие факторы. Основной из них — переход молодых людей к неподвижному образу жизни, в частности, это связано с тем, что современная работа больше связана с сидением за компьютером, чем с физическим перемещением. Даже если большинство из нас стараются питаться более или менее правильно, многие не могут найти времени для физических упражнений.
Следующий фактор имеет социально-экономическую природу. Совершенно очевидно, что ожирение связано с материальным положением людей. Еще сто лет назад все было наоборот, а сейчас недостаток питания в развивающихся странах встречается гораздо реже, чем излишняя полнота. Виновата в этом низкая самооценка и распространившаяся система быстрого питания. Гораздо труднее похудеть, если вы не уверены в себе, если вы видите, что успех приходит к более привлекательным и энергичным людям, и тут замыкается порочный круг. Диета никогда не дает результата мгновенно, а физическая нагрузка, помимо этого, вначале довольно сильно утомляет. И нужно найти в себе силы, чтобы продолжать.
С другой стороны, сети ресторанов МакДоналдc и подобные им заведения обещают дешевую и вкусную еду, а заплатишь чуточку больше — получишь порцию огромного размера. Доллар за калорию — и получите высококалорийные гамбургеры и газировку (все это в некотором роде переработанная кукуруза), и все это в три раза дешевле, чем здоровая еда. Несколько долларов за утреннюю булку с сосиской и кофе. Стоимость еды для семьи из четырех человек в одном из таких ресторанов редко превышает 30 долларов, а вот сбалансированная и свежеприготовленная пища, рекламируемая в журнале «Стиль жизни» Марты Стюарт[1], обойдется вам на 50 долларов дороже. Важную роль играет также недостаток времени, связанный с дополнительной работой, воспитанием детей и просто с желанием расслабиться перед телевизором. [см. «10 системных причин ожирения в США»]
На самом деле вся наша культура еды сводится к быстрому потреблению. В своей замечательной книге «Дилемма всеядных» Майкл Поллан[2] исследует историю процесса производства пищевых продуктов в Америке, начиная с индустриального внедрения кукурузы. Он прямо обвиняет Эрла Батца, секретаря Ричарда Никсона по сельскому хозяйству, политика которого привела к повсеместному увеличению производства кукурузы, направляющейся на комбикормовые заводы Канзаса и мельницы Среднего Запада, которые превращают зерна кукурузы в тысячи наименований сахара в бутылках.
Конечным продуктом вложения десятков миллиардов долларов государственных субсидий в океаны кукурузы на полях Америки в прямом смысле слова являются большие животы и забитые артерии жителей современных американских предместий. Причем сильнее всего страдают те, кто находится внизу социально-экономической лестницы. В следующий раз, когда будете пользоваться системой быстрого питания, мысленно удвойте цену и сравните разницу с теми сотнями долларов, которые вы платите за медицинскую страховку, чтобы покрыть миллиарды долларов, расходуемые на лечение болезней сердца.
И все же дело не только в манере питания. Наши гены играют определенную роль в том, как каждый из нас реагирует на избыточное потребление калорий. Все мы знаем полных людей, которые едят не больше, чем котята, и с завистью смотрят на тех, кто может есть сколько угодно, не увеличиваясь в размере. Все знают голливудскую карикатуру на толстого мужчину, заглатывающего одну за другой индюшачьи ножки, однако болезненное ожирение является в равной степени результатом как «неудачного» набора генов, так и неправильного питания.
Генетика ожирения
Вопрос о роли генов в развитии ожирения можно рассмотреть на примере мутантной мыши, страдающей ожирением. Такая линия в остальном совершенно нормальных мышей появилась в 1950-х годах в лабораторных условиях в результате спонтанной мутации. Животные, наследующие обе копии мутантного гена, действительно очень большие (в четыре или пять раз больше однопометных животных с одной копией мутантного гена), так что в складки их тела можно спрятать всех братьев и сестер. Дело в том, что эти животные не могут контролировать свой аппетит и все время едят.
В середине 1990-х гг. выяснилось, что ответственная за ожирение мутация выключает ген, кодирующий пептидный гормон лептин. Лептин служит одним из основных сигналов, с помощью которого наш мозг заставляет нас перестать есть, когда понимает, что мы уже достаточно насытились. Один из важнейших отделов мозга, называемый гипоталамусом, среди многих других функций отвечает также за ощущение насыщения. Другими словами, мы прекращаем есть не только потому, что на тарелке больше нет еды, но скорее по той причине, что наши чувствительные системы активно информируют нас о том, что пора заканчивать. Если нарушить работу этих датчиков, мы будем есть постоянно, и ожирение нам обеспечено.
Представьте себе, с каким ликованием гиганты фармацевтического бизнеса восприняли бы это известие. Ясное дело, что лептин в качестве лекарственного препарат стал бы панацеей и занял бы место, освободившееся после трагической истории с фенфеном[3]. Но дело в том, что на самом деле у тучных людей уровень циркулирующего в крови лептина обычно выше, чем у людей с нормальной массой тела. Это означает, что у людей с ожирением вырабатывается устойчивость к действию этого гормона. Лишь в небольшом числе случаев в семьях с ожирением имеет место делеция гена лептина, как у упоминавшейся выше мыши, однако в развитии ожирения в общей популяции людей ген лептина играет очень небольшую роль.
Для контроля массы тела применяют три основных класса лекарственных препаратов: «гасители» аппетита, блокаторы углеводов и сжигатели жиров. Фенфен предназначен для снижения аппетита и, как и некоторые другие препараты, ослабляет проведение нервного сигнала в гипоталамусе с помощью серотонина. Не удивительно, что такие препараты вызывают множество побочных эффектов, включая нарушение работы сердца, поэтому их прием связан с высоким риском для здоровья.
Чем больше специалисты занимаются вопросами регуляции аппетита, тем сложнее кажется эта проблема. Сложная регуляторная система поддерживает необходимый баланс, предотвращая экстремальные варианты питания, приводящие к ожирению или анорексии. Спектр гормонов, регулирующих гомеостаз энергии и глюкозы, широк, как рога оленя Санта-Клауса: здесь лептин и висфатин, адипонектин и оментин, грелин и резистин, окситомодулин и амилин, не забудем также пептид УУ, глюкозозависимый инсулинотропный полипептид и глюкагоноподобные пептиды. Действие всех этих сигнальных молекул должно быть скоординировано в зависимости от суточного ритма (ночью аппетит снижается) и нашего самочувствия.
Эта система взаимодействий почти не изменилась за десятки миллионов лет эволюции приматов, но внезапно попала под двойной удар, вызванный значительным увеличением размеров человеческого тела и современными изменениями в характере питания. Действие системы не рассчитано на потребление большого количества сладкой пищи, поэтому естественный баланс нарушается. Геном вышел из равновесия с внешним миром.
Даже из сделанного краткого обзора следует, что с увеличением массы тела могут быть связаны около 20 различных генов. Варианты каждого из этих генов могут вносить свой вклад в эпидемию ожирения. Мы еще не начали говорить о пищеварении, отложении жиров, расходовании энергии или основном метаболизме, но если обратимся к рассмотрению этих процессов, придется упомянуть не менее сотни генов.
Проведенные за последнее время исследования выявили свыше 250 участков в геноме человека, которые могут быть связаны с развитием ожирения, однако конкретные «виновники» не найдены. Каждое из этих исследований в отдельности вряд ли стоит той бумаги, на которой они изложены, и большинство из них не приводит к какому-либо результату. Но если рассмотреть их в комплексе, то следует банальный вывод, что с ожирением связан весь геном человека.
Примерно тот же тезис прозвучал в главе «Геном подростка»: компании часто прогорают не из-за некомпетентности генерального директора, а в результате сочетания множества естественных причин. В каждой компании сотрудники разводятся, пытаются справиться со своими пятнадцатилетними детьми, решают за счет компании свои личные дела или борются с последними версиями компьютерного обеспечения. Слегка измените условия, и сравнительно успешная группа людей может стать нефункциональной. При развитии ожирения происходит нечто подобное.
К каким же результатам привел поиск связи между сотнями тысяч отдельных участков генома и ожирением? Один интересный результат заключается в том, что о склонности конкретного человека к ожирению в большинстве популяций многое может сказать ген FТО. Шестнадцать процентов взрослых, имеющих две копии определенной аллели гена FТО, страдают от избыточного веса приблизительно в полтора раза чаще остальных людей. Напротив, 36% взрослых с двумя копиями другой аллели имеют в два раза больший шанс избежать ожирения, и их масса тела в среднем на 5 фунтов меньше. Эти выводы сделаны на основании изучения 40000 человек в 13 независимых исследованиях, поэтому они кажутся достаточно достоверными. Удивительно, что действие этих генов начинает проявляться в семилетием возрасте, т. е. до того, как ребенок может сам отвечать за свои привычки в еде и физической активности. Это не означает, что во взрослом возрасте такой человек не сумеет избавиться от ожирения, но уже с самого рождения он находится в несколько невыгодном положении.
К сожалению, мы пока почти ничего не знаем о том, в чем же состоит функция гена FТО и почему разные аллели оказывают столь разное действие. Безусловно, вскоре ученые смогут ответить на эти вопросы, но пока это один из поводов для разочарования в современной генетике: как с ценами на бензин, часто мы видим, в чем заключается проблема, но сделать с этим ничего не можем.
Еще одним примером является ген INSIG2. Моим первым впечатлением от статьи, посвященной открытию этого гена, было то, что данную группу исследователей отличает странное чувство юмора и отсутствие страха перед агентствами, финансирующими научные исследования. Такое множество значимых связей между генами и болезнями оказалось ложным, что называть свой ген «ничтожным[4]» означает просто искать неприятностей. На самом деле, оказалось, что INSIG2 — это просто сокращение от определения «индуцируемый инсулином ген 2» (insulin-induced gene 2). По-видимому, кодируемый этим геном белок участвует в синтезе жирных кислот и холестерина, что вполне понятно в контексте проблемы ожирения.
Еще только предстоит установить конкретные изменения в ДНК, влияющие на функцию гена, однако три разных исследования, проведенные в группах из нескольких тысяч белых людей по обе стороны Атлантики и в группе афроамериканцев, показали, что 10% людей, имеющих две копии менее распространенной аллели, с наибольшей вероятностью столкнутся с проблемой ожирения. Этот вариант риска встречается повсеместно приблизительно с одинаковой частотой, и на его долю приходится около 1/3 всех вариантов аллелей. Удивительно, но он не в равной степени способствует развитию ожирения в разных группах населения. По всей видимости, эта аллель является наиболее древней; она присутствовала в геноме, еще когда люди не начали перемещаться по всему земному шару и заниматься сельским хозяйством. Защитный вариант аллели более новый, и это означает, что постепенно возникающий новый генотип должен в меньшей степени способствовать ожирению.
Следующий ген, ЕNРP1, также известный как РС1, выделяется среди других генов, которые могут быть связаны с развитием диабета. Он кодирует белок, который связывается с рецептором инсулина и ослабляет его действие. Если рецептор инсулина — это регулятор на передней панели автомобиля, позволяющий включить кондиционер, то ЕNPP1 — это ручка, отвечающая за подачу воздуха. В клеточной биологии известно множество подобных механизмов, без которых можно обойтись, но которые значительно облегчают жизнь.
Отличие от обычной формы ЕNPP1 человека заключается лишь в одной аминокислоте, и эта замена влияет на связывание белка с рецептором инсулина. Существуют и другие аллели, различия между которыми выражаются в количестве белка, производимого в поджелудочной железе, печени и жировых клетках. Легко себе представить, что такие разные формы влияют на развитие устойчивости к инсулину и, следовательно, способствуют повышенной подверженности диабету. Кроме того, они также являются предрасполагающим фактором в развитии ожирения, не сильным, но заметным, особенно среди представителей европейской расы. А вот в Японии подобного различия обнаружено не было. Такое часто случается при выявлении генетических основ заболевания: действие аллели гена может быть различным в разных группах населения.
Как и в случае рака, лишь небольшая доля всех случаев ожирения действительно объясняется наследственным фактором, который можно связать с известной мутацией. Один из таких случаев — мутации гена рецептора меланокортина-4 (МСR4), ответственного за ощущение сытости. Ген меланокортина относится к тому удивительному разряду генов, которые участвуют во множестве различных процессов. Вариации гена МСR4 ответственны за белые пятна на шкурах наших меховых друзей, вариации гена другого рецептора меланокортина вызывают нарушение эрекции, но и сезонные аффективные расстройства тоже связаны с этим геном. Многочисленные работы, в которых было исследовано несколько десятков тысяч пациентов, позволили выявить слабую связь между общим вариантом МСR4 и развитием ожирения. Я жду того момента, когда в аннотации к лекарственному препарату, изменяющему наш генетический аппарат, будет написано, что если в результате приема препарата для снижения веса у вас возникает эрекция, длящаяся более четырех часов, вам следует обратиться к врачу.
Диабет второго типа
Диабет имеет такую же связь с ожирением, как преступность с безработицей. Одно без другого может существовать, но второе, безусловно, способствует появлению первого. В конечном итоге карта генов, связанных с ожирением, практически полностью перекрывается с картой генов, связанных с диабетом, хотя теперь мы знаем, что эта связь неоднозначная. Многие распространенные варианты полиморфизма вносят свой вклад в развитие диабета, но не играют важной роли в контроле массы тела.
В организме выработался тонкий механизм, основанный на действии инсулина, который позволяет поддерживать определенный уровень глюкозы. В норме избыток глюкозы превращается в запасные жиры. Но если система перегружена слишком долгое время, она попадает в состояние стресса и в конечном итоге сдается. И тогда в результате двойного нарушения равновесия возникает диабет. Первое равновесие — это равновесие между генами и окружающей средой, второе — в действии гормонов, которые в норме призваны обеспечивать запасание энергии в определенных рамках.
Почему организм теряет чувствительность к жизненно необходимому гормону? Почему эволюция допускает накопление генетических вариантов, столь сильно нарушающих передачу сигнала с помощью инсулина? Дело здесь не в плохих генах, а скорее в хороших генах, которые в аномальной ситуации вынуждены совершать неправильные поступки.
После приема пищи уровень инсулина в организме возрастает, поскольку гормон указывает всему организму, что нужно делать с вновь поступившей глюкозой, но высокий уровень гормона не должен держаться постоянно. Хотя производится инсулин только в одном месте, его использование контролируется в каждой конкретной ткани — так получается эффективнее.
Зачем нужен идентификатор телефонных номеров? Телефон — прекрасная вещь, когда он помогает нам не забыть купить помидоры по дороге домой или связаться с любимым человеком, находящимся от нас на расстоянии тысячи километров. Но когда мы только что сели ужинать, а нам по телефону начинают предлагать ненужные нам банковские услуги, это становится неприятной проблемой. Тут может пригодиться идентификатор номеров. Он несколько усложняет сравнительно простое устройство, но позволяет лучше контролировать его работу.
Одна из удивительных особенностей механизма действия инсулина заключается в том, что чувствительность к нему корректируется в соответствии с возрастом. Например, в пубертатном периоде или при беременности наш физиологический ответ на потребление пищи изменяется, и так же изменяется метаболизм глюкозы — частично за счет повышения выработки инсулина. Аналогичным образом обычно нормальный уровень инсулина может оказаться не совсем нормальным, если мы начали набирать вес, занимаемся физическими упражнениями, больны или просто стареем. Поэтому организм постоянно корректирует уровень устойчивости к инсулину в соответствии с уровнем его образования. Повышается выработка инсулина — повышается и устойчивость к нему. Такая система обратной связи позволяет поддерживать в организме равновесие.
У полных людей с постоянно повышенным уровнем так называемых неэтерифицированных (свободных) жирных кислот и толстым слоем абдоминального жира высокий уровень устойчивости к инсулину поддерживается на протяжении длительного времени, отчего организму становится нехорошо. Клетки, которые в норме должны постоянно получать сигнал инсулина, в конечном итоге перестают на него реагировать. На этом этапе возникшая устойчивость приводит к нарушению синтеза инсулина поджелудочной железой в результате гибели β-клеток. Отрицательная обратная связь, которая в норме поддерживает необходимый уровень глюкозы, со временем или в результате стресса становится источником болезни. Гены ни в чем не виноваты, они пытаются все делать правильно, но у них ничего не получается, поскольку они вышли из равновесия с внешними условиями.
Среди 50 генов, которые считаются «виновными» в развитии диабета по данной схеме, три охарактеризованы особенно полно, так что даже самый большой скептик не сможет отрицать их участия в развитии заболевания.
Кальпаин 10 является идеальным примером того, как следует искать гены, связанные с комплексными заболеваниями, а также того, насколько различным может быть действие одного гена в разных группах населения. Среди финнов и мексиканских американцев этот ген отвечает за 10% случаев заболевания, а вот у жителей Японии, Полинезии и Африки такой связи с диабетом не обнаружено.
Полное название гена РРАRG настолько длинное, что утомляет даже биохимиков, — гамма-рецептор, активируемый пролифератором пероксисомы (реroxisome proliferatoe-activated receptor-gamma). Этот ген кодирует рецептор фактора роста, который участвует в регуляции развития адипоцитов (жировых клеток), тогда как кальпаин 10 кодирует белок, расщепляющий другие белки и влияющий на функцию поджелудочной железы. Необычность гена РРАRG состоит в том, что способствующая подверженности диабету аллель является наиболее распространенной аллелью в геноме человека. Свыше 75% людей имеют то, что упрощенно можно назвать «плохим геном», но действие его настолько слабое, что у большинства из нас он не приводит к развитию диабета. С другой стороны, большинство диабетиков имеют эту версию гена, так что в конечном итоге он до какой-то степени объясняет подверженность заболеванию.
Третий ген был выявлен в результате серий сканирования всего генома в конце 2006 г. Это настолько «плохой ген», что имя ему дали совершенно ужасное: ТСF7L2. Можете называть его Лексом Лютором[5], поскольку он кодирует транскрипционный фактор, функция которого состоит в контроле других генов. Как и в случае двух других генов, вариации ТСF7L2 ответственны за появление заболевания у 20% африканцев и европейцев. Что интересно, более старая «аллель риска» почти исчезла у представителей стран Азии, где соответствующие случаи диабета встречаются сравнительно редко.
Развенчание гипотезы об «экономных» генах
Обратимся к вопросу о том, почему аллели риска встречаются в геноме человека с достаточно высокой частотой. Здесь есть три возможности. Во-первых, это результат дрейфа генов, который является настолько несущественным для естественного отбора, что эволюция просто «не обращает на него внимания». Во-вторых, возможно, дело в определенном равновесии между преимуществами и недостатками таких аллелей. В-третьих, некоторые из этих аллелей могут быть действительно полезными, но они еще настолько молоды, что просто не смогли вытеснить более старые варианты. В соответствии с накапливающимися данными определенную роль могут играть все три механизма.
Но давайте начнем нашу беседу с обсуждения популярной теории о так называемых экономных генах. Быть экономным означает мудро расходовать имеющиеся ресурсы, не быть расточительным и думать о будущем. Следовательно, экономные гены должны были бы позволять нам сохранять на черный день определенные пищевые резервы, или запас зерна, или добытое мясо, если речь идет о древнем человеке.
Именно это имел в виду генетик Джим Нил из университета Мичигана, когда в 1962 г. предложил гипотезу об «экономных» генах для объяснения роста численности людей с избыточной массой тела. Те люди, которые лучше умели превращать избыток калорий в запасные жиры, легче переживали периоды голода, достаточно часто возникавшие в истории вида. А вот при столкновении с современной системой питания, в которой жир и сахар присутствуют в избытке, люди с таким генетическим строением склонны к образованию избыточного жирового запаса.
Профессор Нил был одним из ведущих мировых генетиков второй половины XX века, но страдал косноязычием, и в конце своей знаменитой статьи он не сумел четко сформулировать свою мысль. В разделе, озаглавленном «Некоторые евгенические размышления», он сделал следующее ценное замечание:
«Если… растущее давление численности населения означает ухудшение условий жизни с возвращением к сезонным флуктуациям доступной еды во многих частях света, в интересах человечества имеет смысл сохранить диабетический генотип в период нашего временного благоденствия».
Далее он заключил: «Это яркая иллюстрация необходимости осторожного обращения с тем, что на первый взгляд кажется “очевидным” евгеническим суждением». Я бы сказал, что это яркая иллюстрация необходимости выдающимся ученым как следует подумать, прежде чем разглагольствовать о генетических методах лечения во имя будущих поколений.
Ему следовало бы быть более осведомленным. Недостаток пищи создает гораздо больше проблем для человечества, чем сохранение генетических вариантов, как показывают события в Судане или Руанде. Беспокоит также то, как профессор представляет себе западный мир, создающий это преобладание экономных генов у населения развивающихся стран. Однако отнесемся к нему снисходительно. Возможно, он писал свою статью в конце мая, когда серая зима в Анн-Арборе только что прекратила свое восьмимесячное господство, а такой стресс трудно перенести!
[Снисходительность к нему лично – почему бы и нет, но вот понимать, откуда берутся эти прорывы людоедства (евгенического, мальтузианского или расистского) у вроде бы респектабельных и интеллигентных господ, совершенно необходимо. Они органичны для их социального статуса; что ещё выдумает умный и хорошо образованный человек в оправдание собственной привилегированной позиции относительно того самого «человечества», незаслуженность которой он, как профессиональный исследователь, чувствует лучше всего. Исключение здесь – учёные и, шире, интеллигенты антисистемных взглядов – социалистических, коммунистических, которые понимают, как это устроено на самом деле и что «устройство» надо менять. См. «Манифест генетиков 1939 г.»]
Однако евгеника не смогла обесценить теорию экономных генов, которая процветает до сих пор. Людей просто притягивают простые идеи, которые, как им кажется, объясняют все даже без достаточного количества веских доказательств. Отдельные элементы теории кажутся осмысленными. Здесь есть все три положения теории естественного отбора: вариации среди индивидов, некий наследуемый признак и различие в выживаемости, связанное с этим признаком. Соедините все это вместе, и вы придете к выводу, что генетические варианты, предрасполагающие к ожирению, должны подвергаться положительному отбору в современной истории человечества.
Однако более внимательное рассмотрение вопроса позволяет обнаружить три признака, которые являются общими для многих аргументов эволюционной медицины: гиперадаптационизм, ошибка наследования и небрежные количественные расчеты.
Понятие гиперадаптационизма подразумевает следующее: если у организма есть какой-то признак, то он служит для определенной цели. Но теперь мы знаем, что часто признаки появляются как побочное явление какого-то другого процесса. Вместо того чтобы принять адаптацию как данность, эволюционные генетики пытаются найти в ней смысл.
Ошибка наследования — это идея, заключающаяся в том, что если существуют генетические вариации какого-то признака и частота встречаемости признака варьирует в разных группах населения, то генетики обязаны объяснить это различие. Это старая расистская игра в обсуждении IQ[6] американцев, которая приводится в качестве доказательства того, что высокий процент полных людей среди жителей Тихоокеанских островов связан с естественным отбором. Не кто иной, как Джаред Даймонд[7], автор двух блестящих книг о подъеме и падении цивилизаций («Пушки, микробы и сталь» и «Коллапс»), зашел настолько далеко, что предположил, что прародители жителей островов Полинезии имели особенно много экономных генов, иначе они не смогли бы пересечь Тихий океан. Однако капитан Блай[8] и команда его корабля «Баунти» были худосочными представителями Британского флота и тем не менее смогли пережить 49-дневное плавание после бунта на корабле. Существует множество более современных примеров, когда люди переживали и гораздо более долгие мытарства, так что подобное заключение о строгом отборе следует рассматривать достаточно скептически.
Тщательный количественный расчет необходим в каждом случае, когда речь идет об адаптации, но при ближайшем рассмотрении при формулировке гипотезы об экономных генах такой подход применен не был. Можно рассчитать, какое преимущество должен приносить генетический вариант, чтобы, возникнув у одного человека, через несколько тысяч лет охватить одну пятую всего населения. Это значение составляет около 5%; то есть люди с благоприятной аллелью имеют на 5% больший шанс иметь потомство, чем люди без такой аллели.
Однако критик теории экономных генов Джон Спикман (John Speakman) из Абердина (Шотландия) продемонстрировал, что отбор под давлением голода вряд ли был настолько строгим. Обычно от голода в каждом поколении погибают не более нескольких процентов населения, причем обычно самые молодые и самые старые, так что этот процесс практически не влияет на передачу наследственного материала. В любом случае люди обычно умирают от инфекционных заболеваний, а не от голода. Поэтому следует весьма скептически относиться к заявлениям, что диабет возник оттого, что гены, благоприятствующие увеличению массы тела, хороши для кочевников и скотоводов, но не для жителей современных городов.
Есть один ген, который соответствует критериям экономного гена, но он отвечает за толерантность к лактозе, а не за увеличение массы тела. Пока разведение коз и крупного рогатого скота не стало повсеместным, младенцам после отлучения от груди не давали никакого другого молока, люди не ели сыра, йогурта или каких-то других молочных продуктов. Многие из нас способны расщеплять лактозу, поскольку детский вариант гена превратился во взрослый. Фермент лактазо-флоризин гидролаза в тонком кишечнике ребенка превращает лактозу в глюкозу и другие сахара. За последние 10000 лет произошли как минимум две мутации гена этого фермента (одно из этих событий произошло в Европе, другое — в Восточной Африке), которые позволили ему продолжать работать во взрослом организме. Популяционная генетика позволяет рассчитать, что обладание этой мутацией дало людям эпохи раннего скотоводства приблизительно 5%-ное преимущество, и вскоре это привело к тому, что молоко стало одним из основных продуктов питания.
Напротив, наиболее убедительное доказательство того, что подверженность диабету была заложена в геноме несколько десятков тысяч лет назад, а не возникла относительно недавно, на самом деле следует из самого генома. Почти в каждом случае та аллель, которая роднит нас с шимпанзе и другими приматами, как раз и является фактором риска. Другими словами, защитные варианты аллелей начали появляться с возрастающей частотой только в последние тысячелетия. Без этого ожирение и диабет распространились бы еще сильнее, чем это происходит сейчас. Откровенно говоря, пока мы не знаем, какие силы, если такие существуют, благоприятствуют этому процессу, но мы можем поблагодарить небо за то, что это так.
Нарушение равновесия и метаболический синдром
Возможным объяснением распространения факторов устойчивости является наличие отбора против возникновения диабета. Однако это кажется маловероятным, поскольку до недавнего времени эта болезнь была распространена не до такой степени, чтобы влиять на репродукцию. Возможно, существует отбор, основанный на более тонких аспектах питания, а распространение диабета — лишь побочный эффект этого процесса. Возможно и другое: регуляция метаболизма очень сложна, и в ней участвуют сотни генов, каждый из которых имеет различные аллели, так что неизбежно при изменении некоторых компонентов системы у определенных людей вся система выходит из строя.
Одной из центральных идей данной книги является идея о том, что современные люди находятся за пределами нормальной буферной зоны. Вместо того чтобы думать об оптимальной массе тела или уровне глюкозы, нам нужно задуматься о хорошо сбалансированной системе взаимодействий, поддерживающих массу тела и энергетический обмен в пределах определенных рамок при постоянно изменяющихся внешних условиях. С этой точки зрения очень важно существование генетических вариаций, способных противостоять атакам со стороны окружающей среды: сезонному изменению характера питания, засухе, голоду, болезням, беременности и старению. Все эти явления оказывают давление на наш организм, и за гибкий ответ на них ему приходится платить.
На протяжении миллионов лет приматы находились в условиях почти постоянного давления, и у них выработалась гормональная система на основе инсулина, лептина и некоторых других веществ, которая работала достаточно эффективно. Затем люди начали мигрировать по всему земному шару, получили необычный для других приматов месячный менструальный цикл и стали жить дольше, чем когда-либо. В различные моменты времени люди превращались из травоядных в плотоядных и всеядных, из охотников и собирателей — в скотоводов. Наша система метаболизма сбита с толку и находится в состоянии стресса.
Нарушение равновесия. Десинхронизация. Нестабильность. Ошибки. Называйте как хотите, но основная проблема состоит в том, что наш современный образ жизни не соответствует нашему генетическому статусу, формировавшемуся на протяжении миллионов лет. Наша всеядность подвергает нас действию гораздо более широкого спектра токсинов и патогенов, чем тот, с которым приходится иметь дело большинству других видов, и оказывает при этом влияние на работу системы цитокинов и других сигнальных молекул, ответственных за регуляцию метаболизма жиров и сахаров.
Наше стремление к перемещению подвергает нас воздействию различных климатических условий и источников питания, к которым метаболизм вынужден адаптироваться. Все эти и другие способы давления выводят систему генов, связанных с метаболизмом, из состояния равновесия, сложившегося в ходе эволюции млекопитающих. Более трети людей, проживающих в развитых странах мира, находятся под влиянием факторов риска метаболического синдрома, который включает в себя диабет и болезни сердца. Только через десятки, если не сотни, тысяч поколений сможет установиться новое равновесие, которое преодолеет влияние тысячелетий глубоких пертурбаций.
Кроме того, нарушено равновесие между высококалорийным питанием западных людей и их генетической конституцией, однако это нарушение возникло в результате эволюции человека. Если выталкивать устойчивую систему за границы буферной зоны, система разрушится. Мы ежедневно сдвигаем биохимию и физиологию гомеостаза глюкозы, при этом какие-то сочетания генов неизбежно оказываются лучше приспособленными к подобным переменам, чем другие.
Что же нам делать? Западный подход состоит в том, чтобы лечить симптомы. Еще лекарства, пожалуйста! В результате, возможно, все общество будет призывать к применению евгенического подхода, который освободил бы наш общий набор генов от этого бича. И только тогда мы осознаем, что гораздо проще и намного гуманнее изменить свои привычки и свой образ жизни. Изменить то, как мы едим и что мы едим, а также изменить нашу практику воспитания детей, которая стимулирует формирование привычек, угрожающих их будущему благополучию».
Грег Гибсон. Во всём виноват геном. М.: Эксмо, 2010. Серия «Открытия, которые отрясли мир». С.215-256

Несмотря на попсовое название, книга хорошая, очень понятно описано, как «повреждения» в сети межгенных взаимодействий, взаимодействуя с образом жизни и пр. факторами среды, дают или не дают болезнь + рассказано, в чём прокалывается гипотеза «экономных фенотипов», описанная в статье С.А.Боринской.
То есть гены, влияющие на нашу уязвимость к диабету, не «делают» диабет, а лишь обеспечивают «тонкое место», которое порвётся или нет, зависит от среды (в основном) и/или работы других генов (немного), совместно работающих в рамках генной сети, поддерживающей данный участок метаболизма, и вместе контролируемых организмом.
Генные сети: структура, функция, эволюция
«Генные сети – это функциональные группы координированно экспрессирующихся генов, своего рода «морфструктуры на уровне генотипа» (в том числе потому, что иерархически организованы, и управляются сверху, от организма как целого).
«Любая генная сеть (ГС) имеет:
1) группу генов («ядро»), обеспечивающую выполнение ее функций;
2) центральные регуляторы – транскрипционные факторы, организующие гены «ядра» в координированно экспрессирующиеся кассеты генов при взаимодействии с общими сайтами связывания их регуляторных районов;
3) рецепторы, запускающие работу ГС в ответ на внешние сигналы; 4) пути передачи сигналов с рецепторов ГС на ее центральные регуляторы.
Часть продуктов генов «ядра» ГС также играют роль сигналов, взаимодействуя с рецепторами как собственной ГС, так и других генных сетей и формируя регуляторные контуры с положительными и отрицательными обратными связями» (Колчанов и др., 2000).
Режим функционирования ГС определяют: (а) наборы молекул-регуляторов и (б) набор генов «ядра» сети, (в) граф сети – структура связей между ее компонентами, (г) состояние критических параметров внешней и внутренней среды и (д) начальное состояние переменных генной сети. Таким образом, в генных сетях можно выделить три иерархических информационных уровня: функциональный (а и б), чьи закономерности связаны с конкретными молекулярными механизмами взаимодействия молекул; структурный (в); и параметрический (г и д) (Колчанов и др., 2000; Лихошвай, Колчанов, 2008).
Морфологические признаки, представляя законченный результат онтогенетического процесса, хорошо визуализуются. Визуализация генных сетей менее очевидна – в разное время один и тот же ген может участвовать в разных кассетах или в разных генных сетях, изменения начальных параметров меняют режим функционирования генных сетей. Таким образом, фенотипическим гомологическим рядам логичнее сопоставлять не сами генные сети, а их состояния в тот или иной момент онтогенеза.
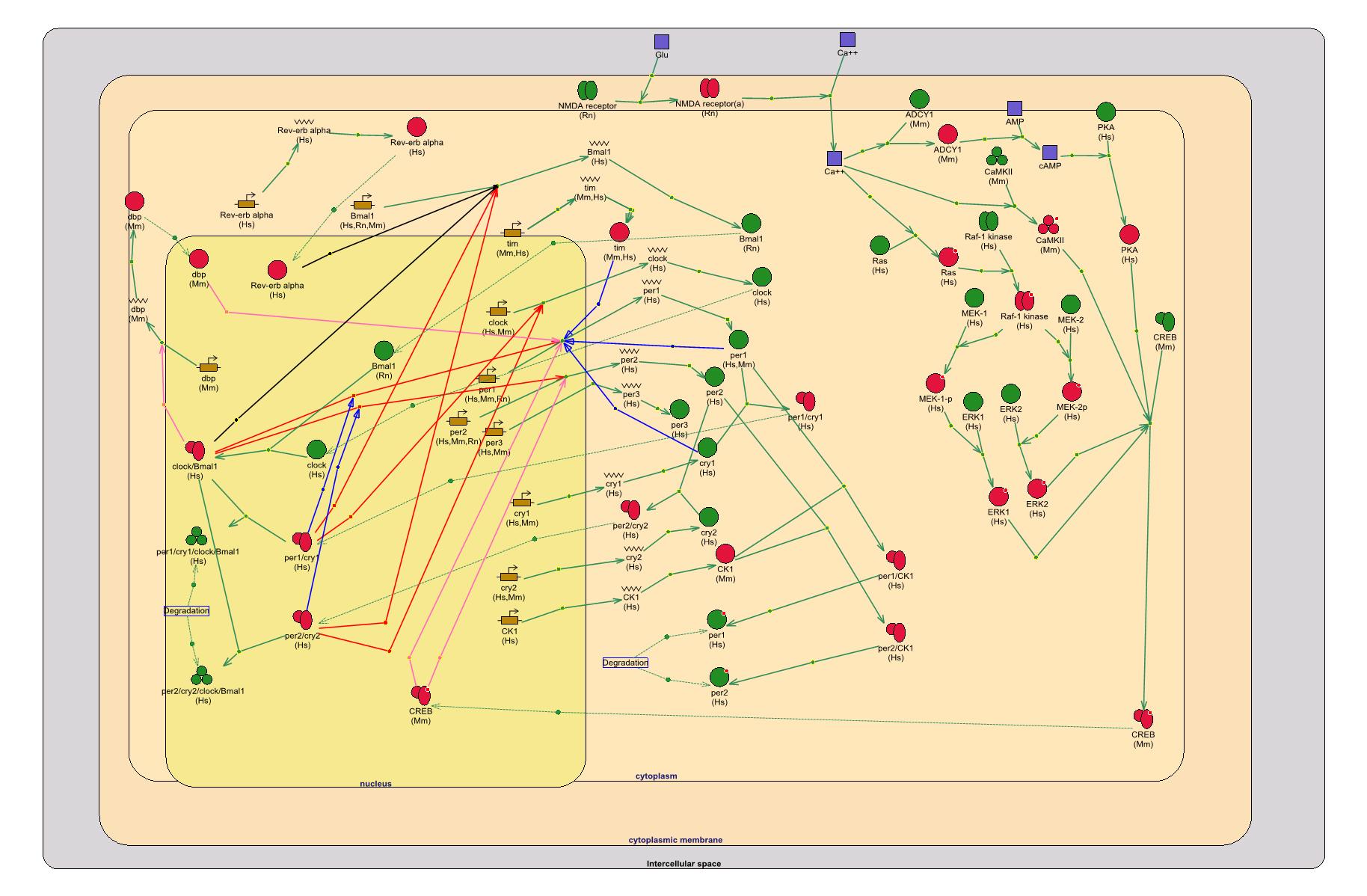
Состояние генной сети – своего рода ее моментальный снимок – можно оценить по микрочиповым профилям коэкспрессии. Паттерны коэкспрессии, консервативные в пределах цветковых растений, образуют, например, гены, входящие в генную сеть формирования цветка (рис. 4, а). В пределах таксона Metazoa консервативно коэкспрессируются гены, входящие в генные сети циркадного ритма (рис. 4, б) или Hh-каскада (рис. 4, в). У прокариот консервативно коэкспресссируются гены базовых генетических процессов и основных метаболических путей (Teichmann, Babu, 2002, 2004).
Для консервативных модулей коэкспрессии был предложен термин «регулог» (regulog). Регулоги были обнаружены как у про-, так и у эукариот. См. рисунок 4.
Рисунок 4. Примеры трех генных сетей, гены которых формируют консервативные паттерны коэкспрессии у разных таксонов – регулоги: а – генная сеть Hh-каскада (по: Nybakken, Perrimon, 2002); б – генная сеть циркадного ритма (по: Badiu, 2003); в – генная сеть морфогенеза цветка (по: Honma, Goto, 2001; Theissen, Saedler, 2001).
У прокариот гены, входящие в регулог, часто сохраняют свое взаиморасположение и положение на кольцевой хромосоме, что связано с оперонной структурой генома прокариот (Teichmann, Babu, 2002; Захаров-Гезехус, 2008). У эукариот в ряде случаев отмечена связь регулогов и определенных районов хромосом (Lindsay et al., 1999; Paterson et al., 2000; Taddei et al., 2001; Teichmann, Babu, 2002; Pauli et al., 2006).
…Паттерны коэкспрессии можно разделить на три типа:
- Генным сетям базовых процессов «домашнего хозяйства» соответствуют паттерны с высококонсервативным составом и коэкспрессией ортологов, а также с высокосвязным графом ГС;
- Паттерны, в которых состав ортологов варьировал при переходе от дрожжей к многоклеточным (сети регуляции транскрипции, межклеточной коммуникации, а также сети сложных белковых комплексов – рибосомы, протеасомы), связаны с появлением новых функций;
- Паттерны, в которых консервативность состава ортологов не соответствует консервативности коэкспрессии (нейрогенез), связаны с дивергенцией функций в эволюции (Chervitz et al., 1998; Peregrin-Alvarez et al., 2003; Stuart et al., 2003; Bergmann et al., 2004).
Три типа паттернов коэкспрессии заставляют вспомнить вавиловский радикал [в законе гомологических рядов Н,И.Вавилова]: тип 1 соответствует левой части, тип 3 – правой части с неустойчивым (варьирующим) фенотипическим проявлением, тип 2 – признакам, которые невозможно включить в радикал на данном таксономическом уровне. Замечательно, что для генов с неизвестной функцией ассоциация с такими паттернами позволяет предсказывать функцию (Stuart et al., 2003; Huynen et al., 2004), что взаимодополнительно к закону гомологических рядов.
Для многоклеточных эукариот функциональный паттерн коэкспрессии распадается на регулоги отдельных клеточных типов и стадий дифференцировки. Так, в общем регулоге, ответственном за сперматогенез, регулог, объединяющий гены, экспрессирующиеся в клетках Сертолли и митотических сперматогониях, выявляется также в различных соматических тканях (в частности, в него входят выше упоминавшиеся гены липидного транспорта). Отдельные частично перекрывающиеся регулоги можно выявить для разных мейотических и постмейотических стадий созревания сперматозойда (Eddy, 2002; Сhalmel et al., 2007).
Нужно отметить, что у мейотических и постмейотических регулогов также отмечено минорное перекрывание с соматическими регулогами (в частности мозг) и более широкое – с малигнизированными клетками (Schlecht, Primig, 2003; Сhalmel et al., 2007; Jacobs et al., 2007; Jacobs et al., 2008).
В регулогах можно выделить «ядро», содержащее гомологичные гены (как ортологи, так и паралоги) и «шлейф» негомологичных видоспецифичных генов. Размеры «ядра» могут сильно варьировать по составу генов и/или по степени коэкспрессии (Bergmann et аl., 2004).
Так, «ядро» небольшой генной сети циркадного ритма у Metazoa (рис. 4, б) консервативно (Badiu, 2003) – мутации в гене казеинкиназы эпсилон I укорачивают циркадный ритм у дрозофилы, грызунов и человека, снижая уровень фосфорилирования белка PER (Vanselow et al., 2006). Мутации по другим казеинкиназам также сохраняют гомологичность, хотя направленность нарушений (удлинение или укорочение циркадного ритма) в разных видах может не совпадать (Xu et al., 2005).
Размер «ядра» связан с эволюционной историей регулога. Так, регулог пролиферации клеток в глобальных генных сетях человека и дрозофилы имеет хорошо выраженное консервативное ядро.
Напротив, регулог дифференциации клеток распадается на перекрывающиеся стадиеспецифичные регулоги, варьирующие как по составу генов, так и по графу взаимодействия между ними, что свидетельствует об их быстрой эволюции (Sharan et al., 2005; Xia et al., 2006). Анализ глобальных генных сетей показал, что наиболее консервативными являются регулоги, содержащие гены, продукты которых функционируют в составе мультимерных белковых комплексов (Sharan et al., 2005).
Менее консервативными оказались регулоги генных сетей онтогенеза и дифференцировки. Напротив, генные сети метаболических путей оказались слабоконсервативными, так как содержали очень небольшое консервативное «ядро» с генами собственно метаболизма и большой варьирующий шлейф регуляторной компоненты (Teichmann, Babu, 2002; Bergmann et al., 2004).
Анализ регулогов показал, что среди транскрипционных факторов сравнительно немного как строго тканеспецифичных, так и функционирующих на всех стадиях онтогенеза. Большинство транскрипционных факторов участвуют в нескольких процессах и экспрессируются на нескольких стадиях онтогенеза (рис. 6) (Teichmann, Babu, 2002; Luscombe et al., 2004; Сhalmel et al., 2007; Jacobs et al., 2008).
Устойчивость регуляторных модулей в эволюции
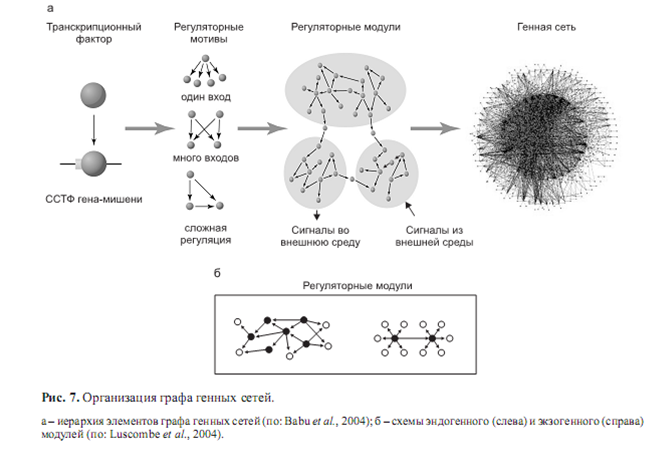
Регулоги имеют иерархическое строение, собираясь из небольших блоков (рис. 7, а) в ходе выполнения той или иной функции.
Анализ таких блоков позволил выявить типовые регуляторные мотивы (Babu et al., 2004; Bergmann et al., 2004; Luscombe et al., 2004) (рис. 7, а). Рассматривая иерархическую организациию генных сетей с позиций регуляции транскрипции, заметим, что низшей ступенью иерархии является функциональная связка «транскрипционный фактор–сайт его связывания в ДНК» (Babu et al., 2004). В результате комбинаторики таких пар и образуются регулоги, комбинаторика которых в свою очередь дает регуляторные мотивы ГС. Мотив может быть представлен (рис. 7, а) транскрипционным фактором, взаимодействующим с кассетой сайтов; триггероподобной структурой, основанной на взаимодействии двух идентичных транскрипционных факторов и сайтов их связывания; композиционным элементом, состоящим из пары взаимодействующих сайтов связывания (Babu et al., 2004) и др.
Комбинаторика мотивов, в свою очередь, образует регуляторные модули, которые, объединяясь за счет генерируемых ими сигналов, формируют регуляторную машину глобальной генной сети организма (Babu et al., 2004; Wittkopp, 2007). Возможность такой комбинаторики обеспечивается общим пулом регуляторных молекул, а ограничение на комбинаторику в эволюции определяет степень перекрывания регулогов, проявляемая в фенотипе как плейотропные эффекты мутаций.
Модули, выполняющие эндогенные (гены домашнего хозяйства и базовых метаболических функций, гены онтогенеза) и экзогенные (рецепция и передача сигнала, стрессовый ответ, регуляция метаболизма) функции, обогащены различными регуляторными мотивами. Экзогенные модули обогащены генными кассетами, регулируемыми единственным центральным регулятором. Эндогенные модули обогащены замкнутыми регуляторными контурами и генными кассетами, включающими множество коактиваторов центрального регулятора (рис. 7, б) (Luscombe et al., 2004).
 Snel и соавторы (Snel et al., 2004) провели попарный внутри- и межвидовой анализ коэкспрессирующихся корегулируемых генов. Критерием корегуляции генов считался высокий коэффициент их коэкспрессии (r > 0,6) при наличии в их регуляторных районах общего сайта связывания транскрипционного фактора. У S. cerevisiae было выявлено 975 таких пар. В выборке коэкспрессирующихся генов C. elegance ортологи таких пар оказались в правом хвосте распределения по величине коэффициента коэкспрессии (рис. 8).
Snel и соавторы (Snel et al., 2004) провели попарный внутри- и межвидовой анализ коэкспрессирующихся корегулируемых генов. Критерием корегуляции генов считался высокий коэффициент их коэкспрессии (r > 0,6) при наличии в их регуляторных районах общего сайта связывания транскрипционного фактора. У S. cerevisiae было выявлено 975 таких пар. В выборке коэкспрессирующихся генов C. elegance ортологи таких пар оказались в правом хвосте распределения по величине коэффициента коэкспрессии (рис. 8).
Высокая степень коэкспрессии ортологов корегулируемых генов S. cerevisiae позволяет предположить, что они сохранили корегуляцию и в геноме C. elegans. Наиболее впечатляющие свидетельства эволюционной устойчивости небольших регуляторных модулей получены при изучении паттернов коэкспрессии у насекомых и хордовых. Так, сходный набор ортологов регулирует формирование фасеточных глаз насекомых и сложных камеральных глаз хордовых (Gehring, 2005). Фасеточные глаза фиксируются палеонтологами не позднее кембрия (~ 570–550 млн лет назад), но согласно палеонтологическим и эмбриологическим данным, предки хордовых никогда не обладали такими глазами. Развитые камеральные глаза впервые появляются в ордовике (500 млн лет назад) независимо у головоногих и хордовых (Михайлова, 1997).
Следовательно, консервативные модули коэкспрессии, ответственные за формирование глаз, возникли еще раньше, возможно, в венде (~ 650 млн лет назад), а потом независимо рекрутировались в ходе эволюции.
Интересно, что для эктопического развития функционирующих фасеточных глаз или (обратный эксперимент) – тканей камерального глаза (Gehring, 2005) оказалось достаточно трансгенеза единственным транскрипционным фактором Pax-6. Попав в различные ткани, Pax-6 – один из центральных регуляторов ГС развития глаза – рекрутировал из разнообразного генетического окружения гены, участвующие в формировании глаз, по сути, восстановив генную сеть.
В морфогенез крыльев насекомых и парных конечностей хордовых также вовлечен общий набор генов – центральных регуляторов, сохранивших сходный паттерн экспрессии. Такой неожиданный результат можно объяснить, лишь предположив, что для формирования крыльев и ног в эволюции был независимо вовлечен древний консервативный регулог, предназначенный для формирования билатерально-симметричных придатков тела. Он образовался, судя по совокупности палеонтологических (появление билатерально-симметричных складок, а затем преобразование их в конечности (Михайлова и др., 1997)) и молекулярно-биологических (филогения отдельных генов (Шаталкин, 2003)) данных, незадолго до кембрийского взрыва (~ 550 млн лет назад) у общего предка Metazoa.
По сравнению с первичноротыми дорзо-вентральная ось хордовых инвертирована, поэтому ноги и крылья соответствуют придаткам одной и той же стороны, иннервировавшимся одной и той же нервной системой (Reichert, Simeone, 2001).
Комбинаторика регуляторных модулей в генных сетях позволяет объяснить быстрое и неоднократное формирование новых органов в эволюции. Так, исследование профилей коэкспрессирующихся генов в плаценте Eutheria показало практически полное отсутствие регуляторных генов с плацентоспецифическим профилем экспрессии.
Дифференцирующаяся плацента оказалась обогащена регулогами с древними генами, ортологичными для эукариот и/или хордовых в целом и ответственными за базовые клеточные процессы. Зрелая плацента обогащена регулогами, включающими таксон-специфичные (от отряда и ниже) гены-паралоги, ответственные за негативную регуляцию ростовых и метаболических процессов. Образование паралогов – результат дупликаций, сопровождавших дивергенцию отрядов и семейств плацентарных (Cross et al., 2003; Knox, Baker, 2008).
Таким образом, в ходе эволюции плацента Eutheria была сформирована блочно-модульным способом из уже готовых генных сетей или их фрагментов[9]. В дальнейшем в ходе эволюционного становления характерной для каждого таксона репродуктивной стратегии (многоплодие или малоплодие; тип потомства «птенцовый» – рождение недоразвитых детенышей – или «выводковый» – детеныши способны следовать за матерью и др.) в ряде таксонов произошло параллельное становление идентичных типов плацент, видимо, тем же блочно-модульным способом.
Так, человекообразные рекрутировали в плаценту ретровирусный блок, связанный с формированием синцитиотрофобласта (Knerr et al., 2004). Эпителиохориальная плацента параллельно развилась у китов, парнокопытных, непарнокопытных, панголинов и лемуров (Сarter, Enders, 2004). Такая плацента обеспечивает низкотравматичные для самки роды малоплодного «выводкового» потомства (адаптивно для видов, испытывающих пресс хищников, либо агрессивной внешней среды).
Древние генные сети, вовлеченные в становление живорождения у млекопитающих, продолжают существовать у остальных хордовых.
Таким образом, предположение Л.С. Берга (1977) о параллельном возникновении живорождения в неродственных таксонах вновь обретает актуальность[10] (с поправкой на роль отбора) и хорошо согласуется с концепцией маммализации (Татаринов, 1976) – параллельного развития маммального комплекса признаков в нескольких линиях терапсид.
Регуляторные модули могут не только иметь транскрипционную природу, но и формироваться на базе связок «миРНК–сайт ее связывания в мРНК». Например, miR319 регулирует экспрессию генов семейства TCP у столь отдаленных растений, как томат и арабидопсис (Palatnik et al., 2003). При повышении интенсивности взаимодействия миРНК с мРНК-мишенью формируется лист с изрезанным краем (норма для томата, мутация для арабидопсиса), а при низкой интенсивности – цельнокрайний лист (мутация для томата, норма для арабидопсиса). Важно, что фенотипические проявления мутационного нарушения такой регуляции сходны, хотя молекулярные механизмы совершенно различны: у томата (рис. 9, а) снижение интенсивности взаимодействия получено за счет мутации, меняющей сродство к миРНК сайта-мишени мРНК гена LA (семейство TCP), а у арабидопсиса (рис. 9, б) интенсивность взаимодействия повышена вследствие усиления экспрессии miR319 (мутация jaw)».
В.В.Суслов, Н.А.Колчанов. Дарвиновская революция и регуляторные генетические системы// Вестник ВОГИС, 2009. №2.
Источник wolf_kitses
Примечания
[1]Марта Стюарт — американская миллионерша, известная телеведущая и автор более 30 книг по ведению домашнего хозяйства.
[2] Майкл Поллан — американский журналист, выступающий на тему здорового питания.
[3]Фенфен — лекарственный препарат против ожирения, состоящий из фенфлурамина и фентермина; препарат был снят с производства из-за тяжелейших побочных эффектов, таких как легочная гипертензия и нарушение работы сердца.
[4]Insig — первые буквы английского слова insignificant, которое переводится как «ничтожный» или «ничего не значащий».
[5]Лекc Лютор — заклятый враг Супермена из одноименного фильма.
[6] «как следует реагировать на заявление, что учёные нашли генетический вариант, который с погрешностью 0,00001 связан с IQ и который встречается у 70% белых людей, но лишь у незначительного количества чёрных? Во-первых, эти цифры ничего не говорят о силе эффекта. Мы уже не раз говорили о генетических вариантах, вероятность появления которых значительно превышает названное значение, но которые при этом объясняют лишь небольшую долю риска развития заболевания. Данные результаты необходимо проверить на десятках тысяч людей, и только тогда они станут в достаточной степени убедительными. Обычно при проведении подобного исследования оцениваемая величина эффекта становится всё менее и менее значительной…
Во-вторых, давайте поинтересуемся, а был ли оставлен обратный эксперимент? Был ли проведён анализ связи с IQ в других расах с последующим выявлением частоты встречаемости таких аллелей среди белых людей? Существует значительный элемент необъективности, заключающийся в том, что генетический анализ в основном осуществляется на жителях Северной Европы и богатых американцах. Можно найти десятки признаков, с которыми связаны аллели, преобладающие у данной группы населения, но это ничего не говорит о том, какие варианты могут присутствовать в других расовых группах и отсутствовать у белых людей, подобный тип исследований должен быть сбалансирован. Когда это условие выполняется, выясняется, что африканцы в целом значительно сильнее других групп различаются по своему генетическому строению и уже один этот факт приводит к выводу, что в ходе миграции по земному шару определённая доля генетического потенциала была утеряна»
Грег Гибсон. Во всём виноват геном. М.: Эксмо, 2010. Серия «Открытия, которые отрясли мир». С.215-256
[7] Джаред Даймонд — современный американский эволюционный биолог и физиолог, автор нескольких научно-популярных работ, объединяющих идеи антропологии, генетики и истории.
[8] Уильям Блай (капитан Блай) — вице-адмирал королевского флота Великобритании; в ходе плавания на «Баунти» в 1789 г. был низложен командой и вместе с некоторыми членами экипажа на баркасе совершил длительный морской переход.
[9] Возможно, еще более глубокая перекомпоновка, объединившая восстановленные (?) эволюционно древние и молодые генные сети, сопровождала возникновение уникального эмбриогенеза млекопитающих, с его комбинацией примитивных (дробление, сходное с таковым у кишечнополостных) и продвинутых стадий (формирование амниотического эмбриона). Кроме того, возникновение трофобласта могло быть связано с адаптивным рекрутированием генных сетей онкогенеза (Eddy et al., 2002; Hiden et al., 2007; Jacobs et al., 2007; Old, 2007), связанных с регуляцией клеточного цикла и межклеточных взаимодействий у Metazoa. Еще один случай адаптивного рекрутирования таких генных сетей описан Н.Н. Воронцовым (1967): в семействе слепышевых (Spalacidae) ворсинчатый эпителий желудка, заселяемый симбиотической микрофлорой и способствующий мацерации растительных тканей, – норма у цокора (Myospalax), землекопов (Cryptomys) и африканских бамбуковых крыс (Tachyoryctes) и онкологическое заболевание у прочих грызунов.
[10]Имеются косвенные данные о параллельном формировании живорождения у сумчатых и плацентарных (Freyer et al., 2003) и частичной гомологии между плацентами разных таксонов живородящих рептилий (Flemming, Blackburn, 2003; Stewart, Thompson, 2003).
Рекомендуем прочесть
 «Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве
«Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве COVID-19 вскрывает правду
COVID-19 вскрывает правду Ревень Rheum palaestinum, самоорошающееся растение пустынь Израиля
Ревень Rheum palaestinum, самоорошающееся растение пустынь Израиля Анатомия кредитного кризиса. Взлет и падение цен на фермерские земли в США в 1920-е годы
Анатомия кредитного кризиса. Взлет и падение цен на фермерские земли в США в 1920-е годы Восприятие шумов
Восприятие шумов Культ книги в РККА: как красноармеец стал главным пропагандистом грамоты в стране
Культ книги в РККА: как красноармеец стал главным пропагандистом грамоты в стране Вибрационные сигналы в коммуникации животных
Вибрационные сигналы в коммуникации животных Марихуановое наступление продолжается
Марихуановое наступление продолжается
