

У людей референциальные жесты намеренно привлекают внимание партнёра к объектам их общего интереса, почему рассматриваются как важный элемент в развитии языка (созданная на основе обмена этими телодвижениями и жестами непроизвольная кооперация и совместное внимание у участников некой деятельности формирует среду, по которой потом «прорастает» обмен языковыми знаками и без которой он бы не мог бы поддерживаться или не был бы эффективен). Помимо людей, подобные жесты типичны только для антропоидов и лишь недавно были найдены у воронов Corvus corax, что интерпретировалось как дальнейшее доказательство сравнимости когнитивных способностей у приматов и крановых.
А вот недавно такой же сигнал обнаружился у хищных рыб коралловых рифов — групера Plectropomus pessuliferus marisrubri и коралловой форели Plectropomus leopardus. Оба вида используют их при коллективной охоте как «указания» на скрытых (в убежище) жертв для их партнёров по охоте, в числе коих гигантская мурена Gymnothorax javanicus, губан-наполеон Chelinus undulatus и осьминог Octopus cyanea. Авторы доказывают, что эти сигналы характеризуются пятью атрибутами, позволяющими заключить, что это именно референтные жесты — они направлены на объект совместной деятельности, но неэффективны как собственно действия, получатель сам выбирает, отвечать на них, или нет, и как именно, их отличает намеренность издавания. Ранее ничего подобного не описывалось у позвоночных с большим (и лучшим) мозгом. Дальше я перевожу статью из Nature Communications с комментариями
«Введение
Содержание
С детства люди общаются, используя жесты (Ж.) в сложных случаях. Ж. определяются как намеренный сигнал, адресуемый реципиенту с целью определённым образом повлиять на его поведение (причём это влияние не «механическое», т. е. жест — знак, а не действие). Дети используют жесты вроде указания, показывания или изображения (руками) для того чтобы обратить внимание партнёра на нечто специфическое в их общей среде.
Такие сигналы определяются как «референциальные» на основе их общей функции. Они могут использоваться [как посредники в кооперативном взаимодействии у детей и антропоидов], чтобы разделить с партнёром их общий интерес или получить для себя нечто осязаемое с его помощью (если сам не можешь). Указание — наиболее распространённый человеческий жест такого типа; развитие соответствующей жестикуляции [как показал Томазелло в «Истоках человеческого общения»] — лучший предиктор языкового развития ребёнка и представляется ключевым компонентом развития языка.
По контрасту с широким использованием этих жестов людьми, находки их у иных видов редки. Большинство примеров — жестикуляция обыкновенных шимпанзе, направленная на людей-экспериментаторов. В природе же они показывают [собрату по грумингу], какую часть тела им хотелось бы чтобы им почесали, и бонобо указывают другим на потенциальную опасность [недавние работы, однако, показывают, что референтные жесты у бонобо и больших шимпанзе распространены не менее, чем у детей. Обыкновенные шимпанзе намеренно используют жестикуляцию при «приглашении» самками подчинённых самцов к спариванию (и наоборот) так, чтобы не видел доминант, пытающийся самок монополизировать, и адекватно модицифируют Ж., чтобы преодолеть возникающие здесь трудности.
То же самое было показано для шимпанзе, обученных языкам-посредникам (language-trained), при наведении на спрятанную пищу людей, не знающих о её местонахождении.Бонобо намеренно жестикулируют при приглашении потенциальных партнёров к сексу, в том числе однополому; то и другое, играет важную роль в снятии стрессов и росте социального статуса обезьян, и они также адекватно модифицируют жесты, чтобы их видел — и ответил — именно нужный партнёр, а все прочие не мешали. Жестовый репертуар больших шимпанзе включает 65 «ручных» единиц и 55 телодвижений, причём в первых характерные признаки референциальности выражены лучше, чем во вторых].
У не-приматов показано использование референциальной «жестикуляции» у воронов, которые для указания показывают и предагают мелкие предметы, функционально для них бесполезные. Кроме собственно указания, это имеет побочный эффект в виде укрепления брачных связей в парах, члены которых= «обмениваются указаниями». Авторы это интерпретировали как ещё одно доказательство сходного уровня когнитивного развития врановых и приматов (итак разделяющих друг с другом множество «интеллектуальных умений», отсутствующих у других форм). Однако экологически обусловленная концепция когнитивных способностей говорит что каждая из них развивается в ответ на конкретную экологическую потребность, а тут она не ясна [концепция в целом скорее неверная — интеллект животных совершенствуется как целое, а не мозаика частных способностей, каждая из которых возникает отдельно в ответ на конкретную «нужду»; особенно это верно для птиц и млекопитающих. Однако же в данном случае следование этой идее «навело» на интересную вещь в коммуникации рыб].
Примером такого отбора на совершенствование именно референтной «жестикуляции», обусловленного необходимостью «указания» на скрывающуюся жертву партнёром по совместной охоте, будут исследованные автором странствующие коралловые груперы Plectropomus pessuliferus marisrubri (далее — «групер»)
Они регулярно охотятся на прочие виды рыб в «сотрудничестве» с гигантской муреной Gymnothorax javanicus (далее «мурена») и губаном-наполеоном Cheilinus undulatus (далее «губан»). Все участники выигрывают от совместной охоты в силу комплементарности их видовых тактик. Груперы — спринтеры, способные догнать жертву в открытой воде, мурены и губаны могут достать жертв в расщелинах и трещинах рифов (куда те прячутся если уйдут от групера) или выпугнуть их оттуда. Мурена может прямо проникать туда благодаря ножевидному телу, губан благодаря сильным выдвижным челюстям (см. название) может втягивая воду, «высосать» жертв из убежища или сокрушить коралловый матрикс вокруг них. Когда эти хищники применяют столь комплементарные тактики одновременно, успех охоты существенно возрастает.

Губан-наполеон
Показано, что груперы для координации совместной охоты используют 2 чётко различимых сигнала. Первый и наиболее часто используемый — «танец шимми» (вибрация всего тела с высокой частотой, выполняемый в горизонтальном положении перед укрывшейся муреной). Он побуждает её присоединиться к груперу и совместно высматривать потенциальных жертв (см. видео S1 здесь). Мурена может и лодырничать — не присоединиться в ответ на первый такой сигнал или слишком рано прервать охоту, юркнув в какую-нибудь трещину кораллов. В этом случае групер часто повторяет сигнал, пока мурена — таки не начнёт двигаться, прерывая телотрясение короткими перерывами, в ходе которых он смотрит — начала она движение или нет?
[Я думаю, он не сигнализирует мурене в том же смысле, как птицы сигнализируют друг другу — и другим видам — о корме или опасности, а стимулирует её органы боковой линии волной воды, распространяющейся при вибрации с «нужной частотой». Так сказать, вырабатывает у неё условный рефлекс и/или включает выработавшийся ранее. Но тут надо смотреть, пойдут ли мурены, никогда не видевшие груперов, за таким сигналом или нет? И наоборот — будет ли групер, выращенный в изоляции, так трястись?]

Гигантская мурена
«Шимми» соответствует определению жеста по Hobaiter & Burne — дискретное телодвижение, бесполезное как собственно действие, нацеленное на конкретного реципиента и вызывающее у него определённое поведение, направленно используемое в конкретной ситуации (что показывает намерение исполнителя вызвать конкретный результат, в данном случае присоединение к совместной охоте). Однако этот жест не референтный — он не направляет внимание мурены на конкретную цель во внешнем мире, потенциальных жертв оба участника ищут оппортунистически, независимо друг от друга. [Впрочем, если это не сигнализация, а стимуляция (т. е. эффективное действие), этот вовсе не жест — хотя намеренность такого воздействия и «понимание цели» у групера уже интересно].
Напротив, второй сигнал груперов прямо показывает на объект; в этом время рыба сама ориентирована вертикально, головой вниз (над скрывшейся жертвой) и совершает чёткие дёргания головой, с перерывами для осмотра вокруг (см. видео S3 здесь). Хотя этот сигнал виден нечасто, он появляется только над местонахождением скрывшейся жертвы при неуспешности первой (преследовательской) фазы охоты груперов. Он как бы показывает её компаньонам — муренам с губанами, обращает внимание их на щель, куда скрылась жертва.
К этой форме сигнализации относятся все 5 признаков, характеризующих референтные жесты по Пика и Буньяру (в работе про воронов). Он направляется прямо на референт, неэффективен как действие (годен лишь для «указания»), ответ на него произволен, а демонстрация — намеренна [хотя также может быть стимуляцией — неслучайно и здесь используется вибрация, а не статичная поза или одиночный звук, как при издавании сигналов-символов у птиц и т. д. позвоночных].
С учётом последнего критерия, очень многие виды активности использовались предварительно как при знак намеренной сигнальной активности. Наблюдаемое данными авторами не вполне сходно с писанным Пика и Буньяром, но также рассматривается большинством авторов как признак намеренности.
Проверка издавания и действия сигнала «стойки на голове» груперов на их партнёров по охоте показывает, что он обладает всеми признаками референциальных жестов. Кроме того, ранее сообщалось авторами, что близкий вид груперов (Plectropomidae) — корраловый P.leopardus охотится в сходной манере на Большом барьерном рифе и использует сходным образом «стойку на голове», только партнёром его оказывается осьминог Octopus cyanea.

Коралловая форель

Осьминог Octopus cyanea
Результаты
Результаты исследований показывают, что:
1) «стойки на голове» прямо указывают на искомый объект
Всего зафиксировано 34 «стойки» (табл.1) в 29 эпизодах «срыва» совместной охоты, всегда в случаях ускользания жертвы в щель (в 4-х случаях было >1 предъявления «стойки»). Их предъявляли минимум 9 различных особей, за общее время 187.5 ч наблюдений за дикими рыбами в природе. «Стойки» появлялись прямо над местом конца неудачной охоты (о чём говорил предшествующий взрывной рывок рыбы на высокой скорости, обрывающийся внезапной остановкой). Из-за сложного рельефа рифового матрикса было трудно проверить, осталась ли жертва в месте «указания» или переместилась в другое незаметно для наблюдателя и возможно для групера.
Т.е. непонятно, «указывает» он на место бегства жертвы или на местонахождение самой беглянки, однако он стимулирует партнёров к их охоте. В 5 случаях после этого скрывшаяся жертва была поймана: в 2 случаях муреной, в 2 — групером, в 1 — губаном.
2) «стойка на голове» не годится как действие
Возможный реципиент находится в зоне видимости групера (дальше «вблизи», см. Методы) во всех 34 случаях исполнения «стоек»: в 14 случаях мурена, в 9 губан, в 10 — мурена и губан вместе и в одном — рыба «жёлтый император» Lethrinus xanthochilus (табл.2).

«Жёлтый император»
Они определены как возможные реципиенты, поскольку неизменно отвечали на «стойку» подходом к соответствующему месту и потенциально способны извлечь беглянку. «Возможные» они потому что при наличии рядом рыб более чем одного вида мы не можем определить, кому адресован сигнал — как считают авторы, до момента, пока один из них не откликнется (на деле и после этого понять адресата нельзя, ибо среагировать может «не тот кто нужен»).
В 7 из 24 случаях, когда рядом с групером, делающим «стойку», рядом была мурена, последняя в 7 случаях перед этим двигалась вместе с групером, в 17 — «тыкалась» головой в щели в радиусе <10 м от движущегося групера (табл.3). В 5 случаях из 19, когда рядом с групером, выполняющим «стойку», был губан, они ранее охотились вместе. 4 из этих 5 «стоек» совершились после того, как губан безуспешно обследовал указанное место в ответ на «стойку», отплывал прочь. В остальных 14 случаях губан, бывший поблизости в момент начала «стойки», обследовал «указанную» щель в течении <1 после этого. какую-то щель (обычно быстрее, но временнЫе детали здесь не очень важны, реакция через <1 мин авторы оценивали как «непосредственно»). Исходя из скорости плавания, вряд ли губан может достичь «указанной» щели и начать инспектировать, «бросившись» к данному месту независимо от сигнала из зоны вне видимости «стоящего на голове» групера. В 5 случаях губан ждал 2, 2, 3, 9, 10, 12 и 25 мин над местонахождением жертвы (не начиная «стойки»), после того как перед этим упустил её (табл.4).
В 2 из 7 этих случаев мурена была в 10 м от групера (хотя, может быть, была ему не видна), но 5 потенциальных реципиентов вообще не было — и групер тянул с началом «стойки».
Как только являлся реципиент и начинал обследовать расселины в этом районе, в 6 случаях из 7 последовала «стойка», почему авторы думают, что групер ждал пока подплывёт поближе тот, кто должным образом откликнется на его сигнал.
Хотя потенциальный реципиент присутствовал при всех 34 «стойках», надо определить, действительно ли это сочетание неслучайно. Для этого использовали данные о плотности видов — партнёров по совместной охоте в основном месте исследований — Mersa Bareika. Плотность мурены там — 1 / 100 м., и в радиусе до 10 м от групера случайным образом она может оказаться с вероятностью 20%. Однако мурены отмечены рядом с груперами совершающими «стойки» в 19 из 29 случаев, значимо более чем случайно (биноминальный критерий, n=29, P<0.001). Там же в среднем приходится 1 губан на 350 м и максимальная дистанция, с которой он может за минуту «скакнуть» к «указывающему» груперу и начать охотиться независимо от его сигналов — 26.4 м., что даёт вероятность случайной ассоциации этих видов 15%. Реально губаны в этих пределах отмечены в 16 случаях «стоек» из 29, т. е. они и до этого охотились вместе с групером или независимо от него подошли обследовать «указанное» место в ответ на «стойку» значимо чаще чем случайно (биноминальный критерий, n=29, P<0.001).
3) Ответ на сигналы групера намеренный
Мурены непосредственно начинают активность в ответ на сигнал. Они начинали движение и обследование в 14 из 29 случаев, когда находящийся рядом групер делал «стойку», что значимо чаще, чем в 11 из 38 случаев, когда мурена начинала движение вблизи не демонстрирующих груперов (Fisher’s exact test: n=62, P=0.033).
Губаны привлекаются «стойками» ещё сильней, чем мурены. Во всех 9 случаях, когда они были единственными реципиентами, они немедленно начинали обследование расселин. Вместе со случаями, когда мурена уже присутствовала, и групер начинал «стойки» после неудачной охоты, в ответ на них появлялся губан, чтобы обследовать расселины (не более чем чере 5 мин в 12 случаях из 23). Эта пропорция 12/23 значимо выше частоты присоединения губанов к недемонстрирующим груперам за период <5 мин после неудачной охоты (6/74; Fisher’s exact test: n=97, P<0.001). См. рис.1
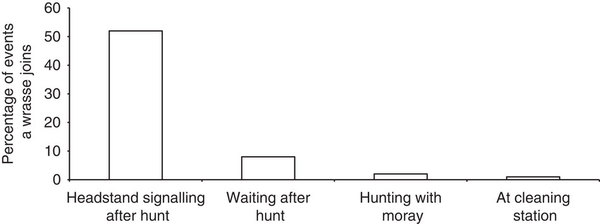
- Привлекательность груперов для присоединения губана. Показан % случаев появление губана в течении первых 5 мин. Рядом с: групером, делающим «стойку на голове», ожидающим без «стоек» после неудачной охоты, просто групера вместе с муреной, на ровном месте (слева направо).
4) Сигнализация групера характеризуется намеренностью
Авторы доказывают, что «стойки на голове» груперов содержат 3 ключевых индикатора намеренности продуцирования: а) они продолжаются неизменно, пока цель не будет достигнута; б) коммуникация ими усиливается, если вдруг попервоначалу не достигнет цели и в) наблюдается дисассоциация между с сигнальными средствами и целями поведения, одна и та же цель достижима использование разных средств и наоборот (что говорит о том, что средства — произвольные знаки, не действия).
В доказательство а) груперы продолжают демонстрировать пока партнёр по охоте не обследует «указанную» рассели ну (27 случаев), или пока не обнаружилось въявь, что сигнал не достиг цели и пришёл альтернативный сигнал (6 случаев), или потенциальный реципиент не ответил (1 случай). В тех 27 случаях, когда потенциальный реципиент откликался и начинал обследование, сигнал удерживался от нескольких секунд до 7 мин. В 4 случаях, когда привлечённый сигналом губан отплыл прочь, групер вновь делал «стойку», что побуждало губана вернуться, и так 3 раза подряд.
Доказательством б) и в) служит реакция групера на неадекватное поведение мурен в ответ на сигнал. Мурены достаточно часто не движутся к месту, «указанному» стойкой: они не пошевелились в 7 случаях и двинулись не в том направлении в 4-х. Это объясняется тем, что мурены движутся, следуя внешнему рельефу, а его складки могут и не направлять их к месту, «указываемому» групером. В 9 из 11 таких охот, когда мурена не обследовала щель, «указываемую» групером (и в 3-х случаях после губана, обследовавшего указанную расщелину, после чего покинул её) подплывал к ней ближе (на расстояние до 20 см) и вновь демонстрировал первый сигнал горизонтальную тряску — «шимми», побуждающую присоединиться к охоте, после чего немедленно плыл на прежнее место и вновь показывал на него «стойкой на голове». В 4-х таких случаях групер заходил сбоку мурены и пытался рылом толкать её в направлении искомой щели (без каких-либо «жестов»).
Подобная «доработка» сигнала-«стойки» (в виде движения к мурене и «горизонтальной трясучки») наблюдалась значимо чаще в случае, когда «стойка» не действовала — в ответ на неё мурена плыла прочь (4 из 4 охот), чем когда «стойка» действовала должным образом, и мурена двигалась к груперу (0 из 4 охот) (Fisher’s exact test: n=14, P=0.001). В целом, груперы редко «заставляют» мурен достичь щели, куда скрылась жертва, при помощи такой «доработки» сигналов (1 из 9 охот).
Коралловый групер также использует «стойки»
Взаимодействия на охоте коралловых груперов и осьминогов на Б.Барьерном рифе столь же комплементарны, как у данного вида груперов с муренами и губанами. Первый преследует жертв в угон в открытой воде; второй имеет длинные щупальца для инспектирования расщелин.
«Стойки на голове» отмечены здесь 5 раз из 62 прослеживаний особей данного вида, в среднем 50 мин хронометража каждое. Только в 3-х из них наблюдатель прибыл раньше, чем сигнализация началась, и видел всё полностью. Во всех 3-х случаях груперы уже были тесно ассоированы с осьминогами перед началом «стоек», производимых после очевидно неудачного броска рыбы, «принявшей» жертву от осьминога.
«Ж.» производились лишь когда осьминог был поблизости (<10 м), и никогда во время охотничьих поисков и бросков, когда его рядом с групером не было. Сигнал значимо чаще появляется в первом случае (5 из 21 протоколов хронометража), чем во втором (0 из 41 протоколов; Fisher’s exact test: n=62, P=0.003).
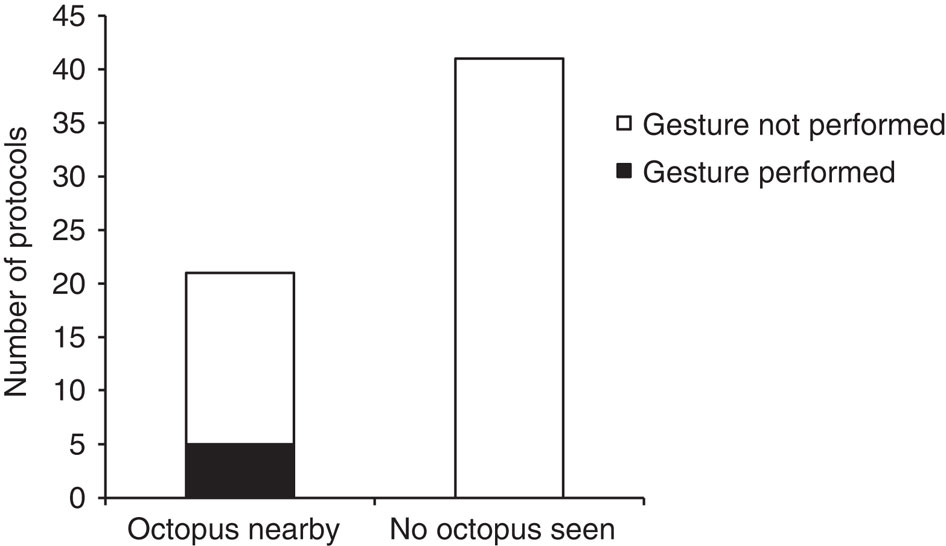
См. рис.2. Вероятность появления «стоек» коралолового групера в присутствие и отсутствие осьминогов поблизости
Лишь в одном случае осьминоги двинулись к месту, «указанному» «стойкой», но они значимо чаще приближали к демонстрирующему её груперу (4 из 5 протоколов), чем к недемонстрирующему (1 из 16 протоколов, Fisher’s exact test: n=21, P=0.004). См. рис.3.
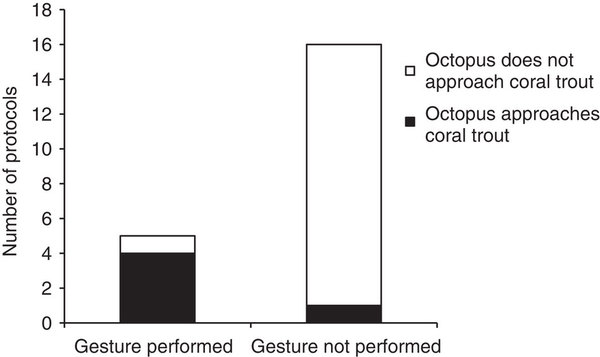
- Привлекательность демонстрирующих и недемонстрирующих груперов для осьминогов
Обсуждение
Авторы считают, что полученные данные дают строгие доказательство использования референтных Ж. двумя видами рыб, групером и коралловой форелью, при совместной охоте с муреной, губаном и осьминогом соответственно [у меня есть сомнения, я бы оценил доказанность этого процентов на 50, тем более что «не признаки определяют род, а род определяют признаки», и искать у рыб материальное соответствие понятию, установленному для людей, по синдрому признаков довольно опасно, можно нарваться на обманчивую конвергенцию]. Сигнал «стойки» «обращает внимание» партнёров групера на объект, «интересный» им обоим, и побуждает к определённым действиям в его отношении и, вообще, соответствует 5 критериям референциальных Ж., предложенным Буньяром и Пикой.
Таким образом, их использование не ограничивается антропоидами и врановыми. Авторы предполагают, что способность к подобному поведению не так редка, как раньше думали, и использование референциальных жестов может быть не только одним из следствий крупного и сложного мозга, но конкретным эволюционным ответом на экологическую необходимость, не связанную с увеличением «общего интеллекта» и не следующим из него. Так, дальнейшие исследования могут показать широкое использование референциальных Ж. у многих позвоночных, включая рыб.
В обсуждении данных по воронам Пика и Буньяр предположили, что в согласии с идеей Л.С.Выготского эволюционно новые когнитивные способности «внутри» особей происходят из «внешней» коммуникации, начинающей управляющей более кооперативными мотивами. Использование референтных Ж. при совместной охоте дополнительно подкрепляет эту мысль; видимо, у других видов подобное поведение следует искать также в кооперативных взаимодействиях. Такая кооперация и используемые в ней сигналы (социальные подсказки, не обладающие всеми признаками референтных Ж.) описана у домашних собак и дельфинах в неволе при коммуникации с человеком относительно общеинтересных для них объектов. В природе такое возможно у ряда видов птиц, вроде шалашников, зеленоспинных овсянок Arremonops chloronotus, аистов марабу и австралийских астрильдов рода Poephila. Все они предъявляют непищевые объекты потенциальному половому партнёру во время ухаживания во время образования пар и/или при последующем токовании в образолвавшейся паре.

Зеленоспинная овсянка Arremonops chloronotus
Австралийские сороки (которые совсем не сороки, а флейтовые птицы) при появлении хищника демонстрируют нечто подобное «указанию», и эти действия вызывают страх у других особей; хотя нужны дополнительные исследования чтобы исключить альтернативные объяснения. Другой возможный пример — медоуказчики, чьё поведение направляет людей (и медоедов — хищников сем.куньих) к скрытому улью. Они «указывают» на цель короткими перелётами в сторону улья, или совершают характерные полёты в том же направлении.
Использование референциальных Ж. у воронов интерпретируется как «значительное когнитивное достижение», а у шимпанзе говорит о наличии ментальных состояний, позволяющих сделать вывод о наличии у них «модели сознания» других особей [не само по себе, но в сочетании с другими признаками, о чём авторы не пишут. Впрочем, «модель сознания» других особей есть не только у обезьян но, и например, у соек — а значит, у воронов есть точно.].
Данная работа показывает, что использование референциальных Ж. само по себе — не повод для утверждений о значительных когнитивных способностях. Отбор вырабатывает функцию скорее чем механизм, различные внутренние (в смысле когнитивные) процессы могут поддерживать сходный паттерн поведения у разных таксонов, но это должно быть установлено экспериментально [в смысле, что нельзя утверждать их единство «на глаз». Я вообще думаю, что в сравнении «жестов» груперов с жестами воронов или шимпанзе видим ещё один случай изоморфизма инстинкта и интеллекта — первый «делает» то же, что и второй, но без «участия» индивидуальности действующего животного, почему не свидетельствует о развитии индивидуального ума, а лишь об экологической необходимости, как в недавнем примере с «понятиями» в коммуникации, использующей сигналы-символы — да и в приводимом авторами примере с «танцем пчёл».].
Авторы пишут, что их случай вносит хороший вклад в поддержку экологического подхода к познавательным способностям у животных, акцентирующего тот момент, что отмеченные когнитивные решения специфических проблем тесно связаны с «узкими местами» биологии вида. Использование орудий в охоте на термитов клопами-хищнецами, планирование путей обхода пауками-скакунчиками или символический «язык танца» у пчёл показывают нам другие случаи, требующие осторожности в приписывании сложных ментальных процессов исходя только из наблюдаемого поведения.
У груперов и коралловых форелей когнитивные процессы, лежащие в основе использования Ж., увы, неизвестны. Однако, хотя рыб традиционно считают «глуповатыми», сейчас растёт число доказательств, что в плане когнитивных способностей они более развиты, чем раньше считали. В данной работе показана способность групера ждать у невидимой жертвы до 25 мин, а это «обезьяний» уровень в задачах на память, используемых для оценки когнитивных способностей. В целом, рыбы способны использовать сложные стратегии поведения в контексте совместной охоты, социального обучения и взаимодействия рыб-чистильщиков с рыбами-объектами «чистки», показывают более сложные когнитивные процессы, такие как транзитивный перенос и способность к генерализации [в первом случае, правда, скорей всего, это следствие инстинктов, реализуемых в агонистической коммуникации самцов астатотиляпий, а не «умозаключений» самих рыб — в отличие от аналогичной «оценки ранга» в стае сосновых соек]. На нейрофизиологическом уровне есть отдельные данные, что система вознаграждения у рыб в мозгу устроена сходно с млекопитающими — скажем, тактильная стимуляция снижает стресс по той же схеме, что груминг приматов».
Т.е. в этом абзаце автор пытается отрицать сказанное в предыдущем, что есть либо ошибка логики, либо предложение натуралистам проверить обе возможности как примерно равно вероятные (надеюсь, второе).
Оригинал работы в Nature Communications
Рекомендуем прочесть
 Ещё про коммуникативную роль жестов у антропоидов
Ещё про коммуникативную роль жестов у антропоидов «Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве
«Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве Как Европа открыла для себя Московскую Русь, а потом закрыла
Как Европа открыла для себя Московскую Русь, а потом закрыла COVID-19 вскрывает правду
COVID-19 вскрывает правду Как соотносятся прямое приспособление и эволюционные изменения?
Как соотносятся прямое приспособление и эволюционные изменения? Малоизвестная победа короля Фейсала над коммунизмом и СССР
Малоизвестная победа короля Фейсала над коммунизмом и СССР Синдром темпа жизни (pace-of-life syndrome, POLS): эволюция концепции
Синдром темпа жизни (pace-of-life syndrome, POLS): эволюция концепции Быстрая перестройка фенологии в новых местах гнездования ласточек
Быстрая перестройка фенологии в новых местах гнездования ласточек
