: базовые принципы и основные направления эволюции")

Крупная стая шилохвостей Anas acuta
Иваницкий В. В.
Введение
Интерес к общественному устройству и социальному поведению гусеобразных имеет длительную историю, отмеченную именами многих вы-дающихся исследователей, в ряду которых нельзя не упомянуть Оскара Хейнрота и Конрада Лоренца. В дальнейшем развитие научных представлений на эту тему происходило под влиянием работ таких известных орнитологов, как сэр Питер Скотт и его дочь Дафила Скотт, Пол Джонсгард, Дуглас МакКинни и многие другие. На современном этапе изучение социальной организации и механизмов коммуникации гусеобразных проводится широким фронтом, в результате чего накоплен значительный объем данных, позволяющий составить довольно полное представление о разнообразии социальных систем в границах всего отряда.
Предлагаемая работа представляет собой попытку анализа этого разнообразия и реконструкции возможных путей его происхождения с позиций филогенетического подхода к анализу эволюции социальных систем животных (Панов, 1983). Обычно принято считать, что темпы эволюции поведения слишком велики для того, чтобы этологические признаки можно было бы использовать для суждений о высоких уровнях дивергенции (Veselovsky, 1975). Ранее мы показали, однако, что этологический анализ достаточно продуктивен при рассмотрении родов в границах одного семейства и даже группы родственных семейств (Иваницкий, 1997). В этой связи одна из задач, которые мы ставили перед собой в рамках предлагаемой статьи, состояла в том, чтобы проанализировать возможности этологического метода на более высоких уровнях дивергенции.
Присутствие в настоящем сборнике обзоров по проблемам классификации, филогении и происхождения гусеобразных освобождает меня от необходимости информировать читателя о составе и таксономической структуре этого отряда. Считаю, однако, не лишним отметить впечатляющее многообразие представленных в нем жизненных форм. Прежде всего, бросается в глаза значительная изменчивость размеров тела – от 16 кг у лебедей до 200–300 г у мелких уток. Затем обращает на себя внимание присутствие в составе отряда как преимущественно сухопутных, шагающих птиц (роды Anchima, Chauna, Anser, Branta, Cereopsis, Chloephaga), так и почти исключительно водных, плавающих и ныряющих видов (роды Cygnus, Oxyura, Aythya).
Пасущиеся на суше гуси и казарки преодолевают сопротивление срываемой растительности, для чего требуются крепкие клювы с утолщением на кончике и зазубренными краями, а также, как можно думать, немалая физическая сила. Та же задача стоит у лебедей, срывающих на плаву подводную растительность, а также у уток, потребляющих двустворчатых моллюсков, прочно прирастающих к донному субстрату (роды Tachyeres и Somateria). Не случайно лебеди, гуси, утки-пароходы и гаги входят в число самых крупных представителей отряда. Что же касается фильтрации, то она не требует больших физических усилий и вполне доступна самым мелким видам при наличии у них соответствующих морфологических приспособлений. Известно, что цедильный аппарат достигает наивысшей степени развития у речных уток (род Anas), составляющих, по всей видимости, одну из наиболее специализированных и прогрессивных ветвей гусеобразных, к которой принадлежат самые мелкие представители отряда.
Стая как ключевое звено социальной организации гусеобразных
Содержание
Когда впервые подступаешься к материалу, относительно которого можно предполагать его системную организацию, всегда ощущаешь желание отыскать некую универсальную зацепку, способную увлечь за собой возможно более густой пучок причинно-следственных связей. Применительно к гусеобразным одной из таких зацепок, как нам кажется, может стать феномен стайности, на важность которого в сфере изучения социальной организации птиц обращали внимание многие авторы (Литвин, 1985; Симкин, 1988; Иваницкий, 1997).
Во-первых, стайность присуща абсолютному большинству видов гусеобразных. Исключение составляют лишь немногие оседлые обитатели тропиков. Во-вторых, стая представляет собой тот тип сообщества, где эти птицы проводят, в сущности, всю жизнь. Период внестайной жизни всегда очень короток, у многих видов его не бывает вовсе. Через процесс объединения выводков восстановление стай исподволь начинается уже вскоре после вылупления птенцов. Но даже в разгар размножения значительная часть популяции гусеобразных представлена негнездящимися особями, которые также держатся стаями. Селезни вновь сбиваются в стаи в разгар насиживания, а иногда еще раньше. В-третьих, нельзя не заметить, что по части размаха стайности гусеобразные стоят на одном из первых мест среди всех птиц. Их стаи бывают огромные – многосотенные и многотысячные, при этом плотность размещения особей в пространстве, как правило, очень высока и на земле, и на воде, и в воздухе. Например, во время осеннего пролета на сжатом поле ячменя в одном месте могут собраться свыше полумиллиона шилохвостей Anas acuta, а зимние стаи австралийского черного лебедя Cygnus atratus, принадлежащего к числу самых крупных представителей отряда, насчитывают до 50 000 особей (Perrins, 1990). Таким образом, в масштабе всего отряда гусеобразных стая представляет собой тот тип сообщества, который уже в первом приближении выглядит как самый универсальный, регулярный и устойчивый.
Настоящим прорывом в области изучения социальной организации стай гусеобразных стала серия публикаций о малых лебедях Сygnus bewickii, зимующих в Англии. Их авторами были сэр Питер Скотт, спаситель гавайской казарки и создатель Треста водоплавающих птиц в Слимбридже, на прудах которого и проходили исследования, а также его дочь Дафила Скотт. Именно П. Скотт в середине 1960-х годов впервые привлек внимание орнитологов к возможности долгосрочного индивидуального распознавания малых лебедей по окраске надклювья. С помощью этого метода были получены свидетельства поразительного постоянства супружеских пар малого лебедя, которого, без всякого преувеличения, называют «птицей, не знающей, что такое развод». Этот метод взяла на вооружение Д. Скотт, и с его помощью ей удалось составить одно из самых первых, строго документированных и развернутых описаний многолетних связей родителей и потомства у гусеобразных и подвергнуть эти связи глубокому функциональному анализу.
Внимание Д. Скотт (Scott, 1980) было сосредоточено на тех причинах, которые побуждают молодежь оставаться на зиму в родной семье – вместе друг с другом и с родителями, в ходе осенней миграции и зимовки. Наблюдая за зимующими стаями лебедей, она выяснила, что чем ближе к родителям находятся птенцы-первогодки, тем выше вероятность их победы в конфликте, и тем больше времени тратят они на беспрепятственную кормежку. С увеличением плотности стаи сплочение семейных групп становится более тесным. Дочери, как более робкие, обычно держатся поближе к родителям, сыновья, обуреваемые жаждой приключений, напротив, чаще пребывают на периферии семейной группы. Исход стычек со сверстниками у первогодков зависит от социального ранга их семей – чем выше ранг, тем выше вероятность победы в конфликте – как в одиночку, так и при поддержке родителей. [См. однотипное явление у обезьян]
Сплоченность семьи, по крайней мере, в первую половину зимы обеспечивается взаимной привязанностью первогодков и их родителей. Но нередко тяготение молодых к своим родителям отчетливо выражено во вторую и даже третью зиму. Но к этому времени отношения родителей со старшими детьми, как правило, становятся асимметричными. Повзрослевшие дети отчетливо стремятся быть поближе к родной семье, черпая в этой близости психологическую уверенность, помогающую им побеждать в конфликтах. Родители, напротив, выказывают равнодушие и всегда прогоняют своих старших отпрысков, если они подплывают слишком близко.
Отношения детей и родителей и поныне остаются одной из центральных тем современного «гусеведения». Самый впечатляющий пример устойчивости семей дают, пожалуй, белолобые гуси Anser albifrons. Например, С. Уоррен и другие авторы (Warren, et al., 1993), изучавшие гренландскую популяцию этого вида на зимовках в Шотландии, обнаружили, что даже в четвертую после рождения зиму 12 % гусей держатся вместе с обоими своими родителями и 35 % – по крайней мере, с одним из них. По данным К. Или (Ely, 1993), белолобые гуси, зимующие в Калифорнии, тоже демонстрируют высокую стабильность семейных групп. В первую зиму с родителями держатся 69 % гусят, во вторую – 39 %, в третью – 38 %. Соответственно, 74, 50 и 39 % сибсов поддерживают друг с другом отчетливые социальные связи. Сплоченность семьи сохраняется вопреки интенсивной охоте и массовым отловам в местах пролета и зимовок. После поимки и кольцевания гуси, выпущенные на свободу порознь, быстро находят друг друга, и семьи воссоединяются.
Многие наблюдатели отмечают, что в стаях гусеобразных царит дух соперничества, создающий в высшей степени конкурентную среду. При этом сразу вспоминается, что среди всех птиц именно водоплавающие располагают наиболее опасным оружием борьбы. Во-первых, как уже отмечалось, многие виды гусеобразных – это крупные и сильные птицы, вооруженные крепкими клювами, самой природой приспособленными к эффективному захвату и отрыванию объектов. Обращая клюв в оружие, крупные гусеобразные способны наносить чувствительные повреждения мягким тканям противника. Сверх того, некоторые виды вооружены костными и роговыми выростами на кистевых сгибах крыльев. Например, у южноамериканских паламедей острые шпоры достигают 4 см, а у африканских шпорцевых гусей Plectropterus gambensis – 2,5 см. Утки-пароходы (Tachyeres spp.), ручьевые (шпорцевые) утки Merganetta armata, новозеландские синие утки Hymenolaimus malacorhynchos обладают массивными кастетами, а у гусей рода Anser сгибы крыльев вооружены твердыми роговыми выростами. Известно также, что удары мощных крыльев лебедей чреваты весьма серьезными ушибами.

Синие утки Hymenolaimus malacorhynchos
Таким образом, сама физическая конституция крупных гусеобразных, в том числе и те ее черты, которые обусловлены особенностями кормодобывания, подразумевает большую вероятность травм при стычках между ними.
О причинах, порождающих соперничество, можно спорить, но несомненно то, что коль скоро оно существует, соперничество это не может выступать в роли самостоятельного фактора, регламентирующего социальные отношения. В этой связи можно предполагать, что разные формы семейной кооперации у гусеобразных, включая перманентные супружеские отношения в рамках долгосрочной моногамии, стали эволюционным ответом на вызовы социальной среды. Твердо установлено также, что долгосрочная родительская опека и даже сам факт присутствия родителей в стае оказывает самое благоприятное влияние на судьбу молодых птиц, как первогодков, так и более старших.
Тема семьи и стаи у гусеобразных затронута в работах очень многих авторов, начиная с Конрада Лоренца. Современный облик этой темы сложился, прежде всего, благодаря исследованиям орнитологов Слимбриджа – уже упомянутой Дафилы Скотт по малому лебедю, а также Джеффри Блэка по белощекой казарке и Денниса Равелинга по канадской казарке. Именно их работами я позволил себе воспользоваться для того, чтобы составить обоб-щенную схему социальной организации многосемейных стай гусеобразных (рис. 1).
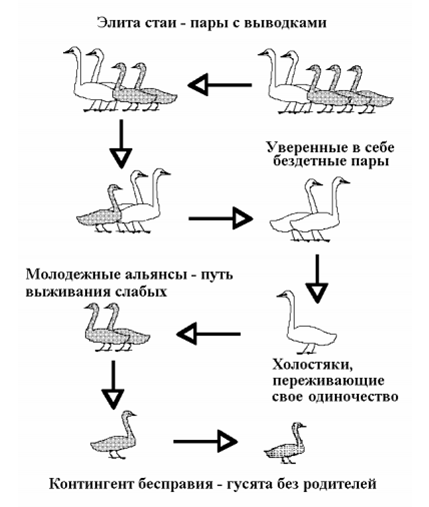
Рис. 1. Векторы доминирования в многосемейных стаях гусеобразных.
Положение особи в стае определяется двумя главными факторами: во-первых, ее принадлежностью к определенному контингенту (поло-возрастной категории или социальной группировке – семье или альянсу) и, во-вторых, ее сугубо индивидуальными качествами (физическими возможностями, агрессивностью и пр.). Элиту стаи составляют семейные пары с потомством. Это особый контингент, представители которого неизменно доминируют в конфликтах с выходцами из всех прочих контингентов, принадлежащих к той же самой половозрастной категории. Указанные на рисунке векторы и уровни доминирования к настоящему времени твердо установлены для многих видов лебедей, гусей и казарок.
Интересный поворот этой темы состоит в том, в какой мере само присутствие молодых или же их количество влияет на конкурентоспособность и иерархическое положение семьи в стае? У всех изученных на этот счет видов, таких как белощекая Branta leucopsis и канадская B. canadensis казарки, белолобые и индийские Anser indicus гуси, такое влияние было найдено. Например, по свидетельству Д. Блэка и М. Оуэна (Black, Owen, 1987; 1989), многочисленные семьи белощеких казарок на зимовках в Шотландии доминируют над малочисленными семьями и бездетными парами. Эти казарки выводят птенцов на Шпицбергене. Когда они в октябре прилетают в Шотландию, в семье бывает в среднем 2,5 молодых, ко времени отлета в марте – 2,1, на промежуточной остановке в Норвегии в мае – за неделю до отлета на Шпицберген – остается 1,8. Первогодки из малочисленных семей кормятся меньше и больше времени тратят на отслеживание социальной обстановки с тем, чтобы избежать агрессии со стороны доминирующих членов стаи. Таким образом, поддержание тесных отношений с потомством на зимовках приносит родителям вполне определенные дивиденды. Некоторые результаты работ Д. Блэка представлены на рис. 2. Самый важный из них состоит в том, что под опекой родителей гусята дольше кормятся и отдыхают по сравнению со своими сверстниками без родителей. В то же время семейные гусята тратят много времени на то, чтобы поддерживать высокий ранг в иерархии стаи. По мнению Д. Блэка, эти усилия не пропадают даром, потому что высокий ранг семьи обеспечивает ее членам право первоочередного доступа к наиболее привлекательным участкам пастбища. Таким образом, вся обстановка в гусиной стае способствует сплочению родичей и объединению их усилий ради удержания высокого общественно-го положения семьи.

Ручьевые (шпорцевые) утки Merganetta armata
Территориальность и колониальность у гусеобразных
Ключевое значение стай в общественном устройстве гусеобразных в значительной мере состоит в том, что именно здесь составляются новые супружеские союзы. При образовании пар абсолютное большинство видов гусеобразных обходятся без специфических брачных посредников, в роли которых у большинства птиц выступают охраняемые и рекламируемые самцами стационарные территории (см. Иваницкий, 1997). Пары у лебедей, гусей и уток образуются непосредственно в стаях – иногда уже осенью, чаще на зимовках, но у некоторых уток, зимующих и мигрирующих преимущественно разнополыми стаями, – на заключительных этапах пролета или даже по прибытии к местам гнездования. Выбор гнездового участка по большей части осуществляет самка, но степень его монополизации, т. е. размеры и конфигурацию гнездовой территории, контролирует самец. Если самке участок не понравился, самец безропотно перемещается вслед за ней на новое место, даже если уже несколько дней охранял прежнюю территорию. Холостые самцы территории не занимают и не рекламируют, если самка погибает, самец тут же оставляет территорию.
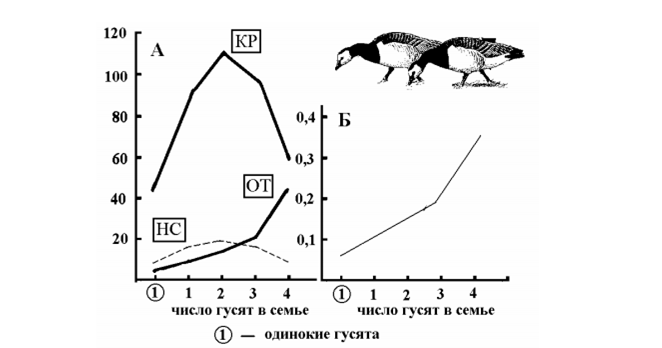
Рис. 2. Зависимости поведения гусят белощекой казарки от их семейного положения в период зимовки (по Black, Owen, 1987, 1989; с изменениями). А – продолжительность в секундах непрерывных сессий активности (КР – кормежка; ОТ – отдых; НС – настороженность); Б – частота конфликтов за минуту наблюдений.
Таким образом, территориальность гусеобразных всегда вторична по отношению к процессу образования и консолидации пары, причем даже у тех из них, у которых территориальное поведение в гнездовой период (лебеди) или даже на протяжении всего года (утки-пароходы) выражено чрезвычайно ярко. В этом мы видим важное отличие гусеобразных от многих других птиц, у которых момент встречи брачных партнеров, вне зависимости от продолжительности их последующих отношений, жестко обусловлен наличием у одного из них территории, на которой происходит только спаривание или протекает весь гнездовой цикл.

Шпорцевый гусь Plectropterus gambensis
Автором одной из наиболее известных классификаций форм территориального поведения уток является Ж. Готье (Gauthier, 1988). На одном полюсе спектра изменчивости он помещает виды, для которых, по его мнению, территориальность свойственна в минимальной степени, если не отсутствует вовсе. К ним он относит, в частности, шилохвость и чирка-свистунка, а также крохалей. Редкие проявления антагонизма у этих видов, отличающихся миролюбием, обусловлены в основном защитой самцами своих самок. Противоположный полюс занимают виды, которым свойственна регулярная и весьма бескомпромиссная агрессия не только при защите самок, но, главное, при защите стационарных гнездовых территорий. Таковы гоголи и пеганки, а квинтэссенцию этого типа составляют крупные южноамериканские утки-пароходы (Tachyeres spp.), известные гипертрофированной агрессивностью при охране территорий. В промежутке между полюсами находятся все прочие виды уток, разделяемые на два класса. Среди известных персонажей относительно более мягкий тип территориальности иллюстрируют кряквы, свиязи и чернети, более жесткий – широконоска, чирок-трескунок, морянка и каменушка.
Классификация выглядит простой и удобной, даже если отвлечься от нечеткости в разграничении двух промежуточных классов. При этом, однако, вне поля зрения автора остались некоторые весьма важные, на мой взгляд, аспекты территориальности, о которых я позволю себе вкратце упомянуть.
Например, американская черная утка (Anas rubriceps) и синекрылый чирок (A. discors) отнесены Ж. Готье к третьему типу. Между тем, после того, как я ознакомился с литературой по этим видам, целесообразность их объединения утратила для меня очевидность. Средние размеры территорий и плотность популяций в период гнездования у них действительно довольно сходные. Но манера охраны территорий совершенно различна.

Чёрная утка Anas rubripes селезень
У черных уток основной способ защиты территории – характерный полет-преследование, известный всем, кому весной довелось наблюдать за утками. Абсолютное большинство территориальных инцидентов у черных уток – это нападения самца-резидента на пары, подыскивающие гнездовые территории, переходящие в довольно длительные погони. Важно, что реакция самца-резидента всегда направлена строго на чужую самку (при этом нередки попытки насильственной копуляции), ее партнер игнорируется. На холостых самцов, попадающих на территорию, самцы-резиденты тоже обычно не обращают внимания, сверх того, в их присутствии хозяин территории может устремиться за пролетающей парой, оставив собственную супругу в более чем сомнительной компании (Seymour, Titman, 1978).

Cинекрылый чирок Anas discors
Что же касается синекрылых чирков, то у них взаимный антагонизм самцов выражен очень ярко. Границы территорий соблюдаются строго. При появлении нарушителя самец принимает угрожающую позу и направляется навстречу ему по воде. Чаще всего нарушитель улетает, иногда взаимодействие переходит в полет-преследование. Объектом атаки в половине случаев служат именно самцы-нарушители, еще половину провоцируют пары, пролетающие поблизости или присаживающиеся на территорию (Stewart, Titman, 1980).
Таким образом, оформление территориальности у разных видов уток может быть очень разным вопреки сходству размеров их территорий. Привлекают к себе внимание и некоторые исключения из общей схемы территориального поведения гусеобразных. Можно упомянуть, например, круглогодичную жесткую территориальность и многолетнюю привязанность к постоянным территориям у оседлых уток-пароходов, шпорцевых уток, новозеландских синих уток, составляющую резкий контраст с краткосрочной сезонной территориальностью большинства гусеобразных. Еще более ярко выделяются на общем фоне, в том числе и в ряду близких видов, африканская савка Oxyura maccoa, австралийская утка Biziura lobata и мускусная утка Cairina moschata.
Это те немногие представители гусеобразных, социо-сексуальные отношения которых строятся на принципах промискуитета. При этом у африканской савки и австралийской утки самцы в одиночестве и по собственной инициативе занимают, охраняют и рекламируют обширные территории, привлекая на них самок для спаривания. Самцы мускусных уток, напротив, территорий не защищают, а пытаются спариваться со всеми самками подряд, причем никакого предварительного ухаживания не бывает и, на взгляд постороннего наблюдателя, копуляции у этого вида мало чем отличаются от изнасилования. Еще один путь специализации в сфере территориального поведения иллюстрируют гоголи (Bucephala spp.), самки которых, в отличие от большинства других уток, после вылупления птенцов защищают хорошо выраженные территории, на которых держатся выводки, часто совсем в другом месте по сравнению с гнездовой территорией, защищавшейся ранее самцом.
Одну из характерных сторон повседневной жизни стай гусеобразных составляет высокая сексуальная агрессивность самцов, что приводит к широкому распространению практики насильственных копуляций (Jonsgard, 1965; Barash, 1978; Afton, 1985; Welsh, Sedinger, 1990; Баранюк, 1999). Возможно поэтому в разнополых стаях уток самки стремятся как можно быстрее заполучить собственного самца и переложить на него значительную часть забот о собственной безопасности. Известно, например, что в утиных стаях свадьбы иногда стартуют с осени, и к середине зимы почти все самки уже держатся в парах. Пары часто распадаются и формируются в новом составе. Но каким бы динамичным не был процесс, принцип парного существования незыблем, и происходит лишь смена партнеров. Утки при этом ведут себя чрезвычайно активно. Оставшись в одиночестве, утка испытывает явный дискомфорт, немедленно пускает в ход средства обольщения и через короткое время заполучает нового защитника, тем более что селезни в стаях, как правило, преобладают над утками. Надо отметить, что утки обладают чрезвычайно эффективными средствами для привлечения селезней, о чем знает всякий, кому случалось охотиться с хорошими подсадными. Кроме того, еще со времен классических исследований К. Лоренца известно, что утки обладают уникальным набором поведенческих реакций, предназначенных к тому, чтобы стимулировать агрессивность опекающих их селезней и направлять ее на соперников [т. н. «натравливание»].
Разумеется, это самая общая картина, которая находится под влиянием ряда причин, таких как степень пространственной разобщенности самцов и самок, уровень развития их репродуктивной системы, соотношения полов в стаях. Даже самый энергичный селезень способен обеспечить своей подруге сносное существование в стае лишь до того момента, пока частота сексуальных покушений на нее не превышает определенного предела. В свою очередь, эта частота зависит от общего количества самцов в стае, их сексуальной активности, пропорции холостых самцов, ведущих себя в особенности навязчиво. Чтобы избавиться от докучливых ухажеров, в какой-то момент самка в сопровождении самца просто покидает стаю. Ключевой момент здесь – это видовые особенности устойчивости самок к социальной стимуляции. Одна самка одновременное ухаживание нескольких самцов расценивает как дискомфортную ситуацию и стремится скрыться, для другой коллективное ухаживание – это норма, не содержащая никаких элементов дискомфорта.
Еще один важный момент в понимании социорепродуктивной специфики гусеобразных состоит, на мой взгляд, в том, какие факторы контролируют созревание яиц. Известно, что у мигрирующих гусей и уток оно начинается еще на пролете под влиянием усиленного питания самок на длительных остановках в промежуточных пунктах с обильными пищевыми ресурсами. Созревание яиц начинается задолго до прибытия в гнездовой район и расселения по территориям, еще в стаях, и, как можно думать, находится под влиянием мощной социальной стимуляции. Обычно пары гусей и уток приступают к поиску гнездового участка и постройке гнезда за несколько дней до снесения первого яйца, но многие самки несут яйца, не имея собственных гнезд. Это неизбежно ведет к массовому отходу яиц и широкому распространению внутривидового и межвидового гнездового паразитизма. В работах Е. В. Сыроечковского (1979) и В. В. Баранюка (1999), посвященных колониальному гнездованию и размножению белых гусей (Chen caerulescens) на о. Врангеля, можно найти весьма впечатляющие примеры на эту тему. В холодные вёсны с поздним таянием снега гуси оказываются перед лицом жесткого дефицита пригодного для гнездования пространства.
В результате огромное количество гусынь оказываются без собственных гнёзд и несут яйца в чужие гнёзда или в случайных местах. Зависимость масштабов гнездового паразитизма у гусеобразных от наличия мест, пригодных для гнездования, хорошо известна. Тем больший интерес, на мой взгляд, представляют случаи массового распространения этого явления в тех популяциях, где подобного рода дефицит вряд ли имеет место. Сошлемся на данные А. В. Кречмара (1995) по гнездованию шилохвости в долине р.Анадыря. Средняя плотность популяции составляла здесь всего лишь 1 гнездо на км2, но это не мешало самкам регулярно подкладывать яйца как в чужие гнезда своего вида, так и в гнезда свиязи (A. penelope).

Cвиязь Anas penelope
К началу гнездового сезона развитие социорепродуктивных отношений в стаях гусеобразных достигает такого уровня, что возникновение колоний воспринимается как нечто закономерное, специального объяснения не требующее. Столь же естественным выглядит и широкое распространение колониальности, наиболее продвинутые варианты которой представлены у лебедей, гусей, казарок, гаг, пеганок, речных и нырковых уток. Тем не менее, взгляд на колонию, как на осевшую на гнездование стаю, по крайней мере применительно к гусеобразным, выглядит скорее метафорой, нежели отражением реального хода событий. Многие колонии являются вторичными агломерациями и формируются в результате сближения пар, достигших к этому моменту значительной степени пространственной разобщенности по сравнению с исходными стаями. В колонии птицы привязаны к гнёздам и лишены свободы маневра, поэтому социальная среда здесь становится ещё более конфликтной, чем в стае.
В то же время трудно спорить с тем, что по общей картине социальных взаимоотношений колония гусей или казарок сильно напоминает стаю, тем более что механизмы пространственного разобщения семей в колонии весьма несовершенны. Например, по наблюдениям Е.В.Сыроечковского (1976), в колонии белых гусей неприкосновенность территории надежно обеспечивается только постоянным присутствием на ней гусака. Стоит ему улететь на кормежку, как самка сейчас же подвергается нападению соседних самцов. По свидетельству С.П.Харитонова (1988), отношения соседних пар белых гусей в колонии часто асимметричны и строятся не на равных, а по принципу субординации. Это значит, что территория низкоранговой пары открыта для посещений соседями, занимающими более высокий иерархический статус, в то время как ответные визиты возможны лишь в отсутствие хозяев. Таким образом, в колонии гусей, как и в стае, достаточно отчетливо проявляется ранжирование семейных групп.
Причины, побуждающие гусеобразных селиться колониями, вызывают много споров. Переуплотнение, несомненно, может навязываться простыми внешними обстоятельствами, такими, как дефицит самых привлекательных участков, например, мелких островов или колоний чаек или крачек, составляющих излюбленное место гнездования многих пластинчатоклювых. В то же время плотные поселения могут возникать вследствие взаимного тяготения особей, однако доказать существование такого стремления бывает трудно. В поисках объяснения происхождения колониальности гусеобразных, мне кажется, полезно иметь в виду, во-первых, ее чрезвычайно широкое распространение в пределах подотряда и, во-вторых, ту высокую свободу взаимного размещения семей в пространстве, которую мы находим в популяциях большинства видов (Сыроечковский, 2000). Контрастные типы пространственного размещения размножающихся пар обнаруживаются даже в ряду близкородственных видов, например, среди лебедей (малый лебедь – строгая территориальность, черный лебедь – высокая степень колониальности, лебедь-шипун – в разных популяциях можно наблюдать оба упомянутых типа пространственной структуры). Мне кажется, что истоки такой свободы, по крайней мере, отчасти, лежат в отказе подавляющего большинства гусеобразных от использования гнездовой территории в роли брачного посредника, а также и в роли ключевого резервуара энергетических ресурсов, обеспечивающих формирование яиц у самок (по крайней мере, у перелетных видов). С этой точки зрения самым «экзотическим» проявлением социального поведения гусеобразных выглядят вовсе не колонии, а наиболее продвинутые и специализированные формы территориальности, подобные тем, которые наблюдаются у малых лебедей (Литвин и др., 1999) или уток-пароходов (Livezey, Humphrey, 1985; Nuechterlein, Storer, 1985).
Основные типы социальных систем гусеобразных
Мои представления о типологии социальных систем гусеобразных отображены на рис. 3, где приведены три основные типа организации сообщества – многосемейные стаи, пространственная сегрегация полов и территориальная изоляция семей. В целом эволюционный веер социальных систем гусеобразных выглядит довольно узким и состоит из вариаций, принадлежность которых к общему типу или, как говорят морфологи, к единому плану строения не вызывает сомнений. Края этого веера образованы социальными системами, которые условно можно назвать «гусиной» (многосемейные стаи) и «утиной» (пространственная сегрегация полов), имея в виду, разумеется, что речь идет о жизненных формах, а не о таксонах. Сделав эту оговорку, мы можем утверждать, что главный водораздел между «утками» и «гусями» проходит в зависимости от сроков физического и социального возмужания потомства и от соотношения конкурентоспособности молодых и взрослых птиц. У «гусей» первогодки сравниваются по весу со взрослыми птицами только к окончанию первой зимовки и уступают им во всех конфликтах вплоть до собственного бракосочетания. У самых крупных лебедей и гусей это происходит в возрасте 3–5 лет. Кроме того, процесс социализации у гусей протекает на фоне высокой и практически перманентной агрессивности членов стай, прежде всего, самцов. Сильные, агрессивные и сексуально озабоченные самцы поддерживают в стае и в колонии атмосферу насилия, в которой слабые самки и молодые без помощи самцов выжить не могут. Иными словами, самцы должны присутствовать в стае для того, что-бы защищать свои семьи от других самцов.
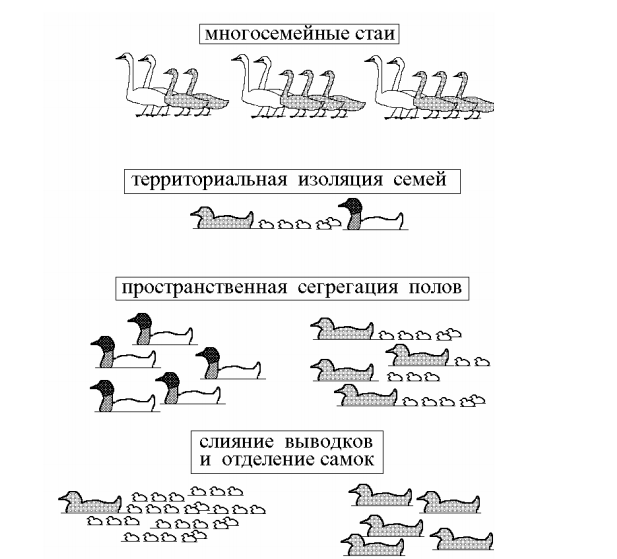
Рис. 3. Основные типы сообществ гусеобразных на ранних стадиях социализации потомства.
Утки в эту ловушку не попали, и тот эволюционный маневр, который они для этого проделали, весьма впечатляет своей простотой и изяществом. Все проблемы были решены путем временного разделения полов на период насиживания кладок и вождения выводков. Тем самым самки и молодежь избавились от всякой конкуренции и беспокойства со стороны самцов. Утята взрослеют быстро и к осени по весу сравниваются со взрослыми. Их социальная среда вплоть до начала зимовки состоит из равных по силам сверстников и относительно миролюбивых самок. Когда начинается образование пар, самцы-первогодки вполне успешно конкурируют со старшими самцами и весной, как правило, благополучно приступают к размножению.
Быстрой модификацией как «гусиного», так и «утиного» типов социальной организации (многосемейные стаи) можно считать социальные системы некоторых оседлых видов, построенные на многолетней круглогодичной привязанности супругов как друг к другу, так и к постоянным, как правило, хорошо обособленным друг от друга и активно защищаемым участкам обитания. Вскоре после подъема на крыло молодые покидают семьи и сбиваются в стаи, где и проживают вплоть до наступления половозрелости и бракосочетания, после чего новобрачные приступают к поиску свободной территории. Так выглядит, например, структура оседлых популяций лебедя-шипуна в Великобритании и черного лебедя в Австралии. Среди наиболее ярких представителей этого типа вновь упомяну уток-пароходов, по склонности к экстремальной агрессии вполне заслуживающих названия «утки-убийцы». Это весьма внушительные создания весом от 4 до 6 кг, вооруженные крепкими клювами и массивными костяными бляшками на сгибах крыльев. К этому же типу «территориальных экстремалов» принадлежит и новозеландская синяя утка. Самцы и самки этого вида круглый год живут парами на обособленных территориях, активно защищаемых в репродуктивный сезон. Патрулирование территорий сопровождается жестокими драками самцов, в ходе которых они стараются нанести удар хорошо заметными на их крыльях кистевыми выступами. Выводок опекают оба партнера. Молодые остаются вместе с родителями около 5–6 месяцев – пока не приобретут взрослый наряд, после чего покидают родную территорию (Kear, Steel, 1971).
Вариативность в рамках этого типа зависит главным образом от площади и конфигурации территорий. Так, компактные территории уток-пароходов располагаются на морской литорали, в местах, богатых моллюсками, и защищаются круглогодично от всех водоплавающих птиц, причем настолько рьяно, что не всякой лысухе или чернети, подвергшейся нападению, удается выжить. Внутривидовые стычки на почве раздела и охраны территорий также отличаются крайним ожесточением, почти всегда ведут к кровопролитию и нередко заканчиваются гибелью слабейшего. У ручьевых уток территории вытянуты вдоль русел горных потоков на расстояние 1,5 – 2,0 км и защищаются не менее основательно.
На рис. 4 я попытался изобразить распределение разных типов социальных систем по разным подсемействам и трибам гусеобразных. Филогенетическая приуроченность этих типов очевидна вопреки резким различиям в условиях обитания. Например, «гусиный» тип сообщества с многосемейными стаями, перманентной моногамией и длительной бипарентальной заботой о потомстве распространен от арктических тундр, где живут казарки и гуси, до водоемов тропической зоны, населенных древесными утками. Такое же распространение имеет и «утиный» тип сообщества с краткосрочной сезонной моногамией, гинелатеральной заботой о потомстве и его быстрым отделением от самок. Что же касается перманентной территориальной изоляции семей, она возможна лишь при оседлом образе жизни и, по понятным причинам, распространена почти исключительно у тропических форм.

Рис. 4. Распространение основных типов сообществ по трибам гусеобразных (изображения птиц заимствованы из Lack, 1974).Типы сообществ: МСС – многосемейные стаи; ТИС – территориальная изоляция семей; ПСП – пространственная сегрегация полов; VAR – разные типы.Трибы (слева направо и сверху вниз): древесные утки (Dendrocygnini); гуси, казарки, лебеди (Anserini); полулапчатые гуси (Anseranatini); пеганки, огари и др. (Tadornini); утки-пароходы (Tachyerini); нырковые утки (Aythyini); речные утки (Anatini); крохали, гаги, гоголи и др. (Mergini); мандаринки, мускусные утки и др. (Cairini); савки (Oxyurini); ручьевые утки (Merganettini).
Практически все трибы гомогенны по типу социальной организации. Исключением являются Anatini, Oxyurini и Cairini. У первых отчетливо доминирует система пространственного разделения полов, но некоторые тропические виды практикуют систему территориальной изоляции семей. Триба Cairini с этологической точки зрения изучена поверхностно, и к тому же ее монофилетичность оспаривается многими авторами (Поповкина, Поярков, 2002). У савок известны, по меньшей мере, два контрастных типа социальных систем. По наблюдениям У. Зигфрида (Siegfried, 1976), самцы южноафриканской (Oxyura maccoa) в гнездовой период защищают и активно рекламируют обширные территории диаметром до 80 м и с хорошо обозначенными границами. Система спаривания носит характер типичного промискуитета. Постоянные пары не образуются, оплодотворенные самки в одиночестве отправляются на поиски места для гнезд, а самцы продолжают рекламировать свои территории, привлекая на них новых самок. В то же время социальная система североамериканской савки (O. jamaicensis) вполне однозначно принадлежит к “утиному” типу. Самцы не занимают территорий, их ухаживание за самками обычно носит коллективный характер, после образования пар самцы вслед за своими утками направляются на места устройства гнезд.

- Южноафриканская савка Oxyura maccoa
Филогенетические аспекты социальной организации гусеобразных
В последние годы многие орнитологи склоняются к тому, что на эволюционном древе пернатых ближайшими соседями гусеобразных являются курообразные. С позиций этой гипотезы наиболее примитивными представителями гусеобразных считаются три вида неотропических паламедей, составляющие отдельный подотряд, который, как иногда полагают, связывает гусеобразных с курообразными. Ближайшим родичем паламедей в подотряде пластинчатоклювых считают австралийского полулапчатого гуся (Jutglar, 1992).
Что по этому поводу могут сказать этологи? Прежде всего бросается в гла-за, что аналоги «гусиного типа» социальной организации среди куриных в са-мом деле представлены довольно широко. Например, у цесарок (Numididae) и многих куропаток (Phasianidae) стая является той социальной средой, где образуются постоянные пары и протекают процессы социализации молодых птиц. Накануне начала кладки пары покидают стаю и уединяются на обособленных участках. После вылупления птенцов семьи вновь сбиваются в стаи. (Потапов 1987). В противовес этому, сообщество уларов можно считать аналогом «утиного» типа организации. Пары уларов образуются в стаях и рассредоточиваются по обособленным гнездовым участкам. Вскоре после начала насиживания самцы покидают самок, сбиваются в стаи и откочевывают на линьку (Базиев, 1978). Таким образом, если сравнивать спектры социальных систем куриных и гусеобразных, то между ними легко обнаружить частичное совпадение.
«Гусиный» тип сообщества, присущий наиболее архаичным представителям гусеобразных, таким как пеламедеи, полулапчатые гуси, настоящие гуси и древесные утки, сближает их с более древними ветвями отряда курообразных, к числу которых относят фазановых. Именно он и является, по-видимому, базальным типом в социальной эволюции обоих отрядов. «Утиный» тип устройства сообщества представляет собой явно родственную, но более специализированную, производную форму организации социума. Перманентная территориальная изоляция семейных пар утверждается в роли ведущего принципа социальной организации как относительно быстрая модификация обоих типов – как гусиного, так и утиного, в процессе перехода к полной оседлости. Так выглядит социальная организация паламедей. Среди курообразных подобный тип устройства сообщества господствует у южноамериканских краксов (Cracidae). [это неплохо согласуется с современными данными о переходных формах и родственных связях внутри Galloanseres].
Что же касается курообразных, то их социальная эволюция на продви-нутых этапах привела к коренной перестройке базовых институтов сообщества. Мы видим это у сорных кур (Megapodidae), тетеревиных (Tetraonidae), индеек (Meleagridae) и у многих Phasianidae (фазаны, павлины, трагопаны, дикие куры). Средоточием ключевых социальных процессов и местом образования пар служит у них не только стая, но и территория, которая вновь обретает наиболее типичную для нее функцию брачного посредника, причем как у моногамных, так и у полигамных видов (Потапов, 1987). На мой взгляд, это уже радикальная ломка, равнозначная в какой-то мере смене плана строения в морфологии. Что же касается гусеобразных, то среди них система диффузных токов в сочетании с полигамией и гинелатеральной заботой о потомстве имеется лишь у некоторых савок р. Oxyura и близких к ним австралийских уток р. Biziura (Johnsgard, 1978).
Сравнивая спектры социальных систем гусеобразных и курообразных, нельзя не видеть, однако, и некоторых глубоких различий между ними. Главным, на мой взгляд, является полное отсутствие среди последних тенденций к колониальному гнездованию и весьма ограниченное развитие стайности.
Между тем, в отряде гусеобразных, как уже подчеркивалось выше, колониальность и стайность не только принадлежат к числу базовых принципов социальности, но и, по всей видимости, являются очень древними признаками. В этом отношении гусеобразные больше напоминают аистообразных и фламинго. Фламинго – высокоспециализированные фильтраторы, составляют стоящую несколько особняком группу с неясными эволюционными связями. Иногда их считают промежуточным звеном, связывающим аистообразных и гусеобразных. Недавно получены свидетельства того, что близкими родственниками фламинго могли быть ископаемые Presbyornis (Feduccia, 1978). Это были примитивные околоводные птицы, давшие начало фламинго, а также шилоклювкам и ходулочникам из семейства Recurvirostridae отряда Charadriiformes. Примечательно, что как шилоклювки, так и ходулочники по уровню развития колониальности превосходят всех куликов. Например, у австралийского ходулочника Cladorhynchus leucocephalus известны колонии численностью более 10 000 гнездящихся пар с плотностью населения до 18 гнезд на квадратный метр (Pierce, 1996). Что же касается фламинго, то на пути прогресса стайности и колониальности они обошли практически всех современных птиц. Таким образом, версия о родстве гусеобразных с аистообразными, фламинго и куликами через ископаемых Presbyornis содержит в себе весьма соблазнительные возможности для объяснения происхождения колониального гнездования и стайного образа жизни во всех упомянутых группах.

Австралийский ходулочник Cladorhynchus leucocephalus
В этой связи уместно вспомнить и еще об одной, весьма своеобразной особенности поведения, присущей многим видам гусеобразных. Это ярко выраженная тенденция к объединению и перемешиванию выводков одного и разных видов (Beauchamp, 1998). В наиболее яркой форме эта черта про-является у гаг. В результате слияния выводков у них образуются весьма многочисленные табунки, опекаемые несколькими самками, обычно не проявляющими взаимного антагонизма. Интересно, что интенсивное перемешивание выводков происходит даже у гоголей – вопреки ярко выраженной территориальности самок, причем перемешивание и объединение птенцов происходят именно во время территориальных конфликтов (Savard, 1987). Широкое распространение этого поведенческого феномена среди гусеобразных едва ли можно объяснить просто как эпифеномен их принадлежности к птицам с выводковым типом развития. Скорее всего, здесь мы имеем дело с совершенно особым типом поведения, сходным с формированием табунков птенцов у некоторых птиц с высоким уровнем развития колониального гнездования. Такие табунки чрезвычайно характерны, в частности, для фламинго, а также для шилоклювок и ходулочников (в том числе и для вышеупомянутого австралийского ходулочника) (Pierce, 1996).
Заключение
Начиная с Конрада Лоренца, все ученые, посвятившие себя исследованию гусеобразных, неизменно подпадали под обаяние этих птиц. Все, кому приходилось подолгу общаться с гусями, казарками, лебедями, без колебаний причисляют их к интеллектуальной элите царства пернатых и приписывают им поистине бесконечное совершенство психики.
Один из самых интересных вопросов состоит в том, существует ли устойчивое доминирование в отношениях особей, принадлежащих к одному контингенту и одной категории? В какой мере реализуется принцип индивидуального соподчинения в отношениях равных по касте? Пока что данные на этот счет выглядят скудными и противоречивыми. Никто из орнитологов, подвизавшихся на поприще изучения гусей и казарок, как будто не сомневается в том, что эти птицы наделены выдающимися способностями к запоминанию облика многих социальных партнеров. Самое оптимистическое высказывание принадлежит, пожалуй, Д. Равелингу (Raveling, 1970), полагающему, что гуси способны запоминать и годами хранить в памяти десятки и сотни «лиц» соплеменников, что дает птицам неограниченные возможности выстраивать субординацию исключительно на принципах индивидуального опознавания и сугубо личного соподчинения. Возможно, он прав. Во всяком случае, хорошо известно, что гуси, разлученные в процессе отлова и кольцевания, легко находят друг друга и воссоединяют семьи. Как уже отмечалось выше, в зимних стаях малых лебедей молодые птицы в возрасте двух и даже трех лет проявляют определенное тяготение к своим родителям. Между тем, на местах гнездования никаких связей взрослых размножающихся лебедей со своим потомством прошлых лет не отмечено. Возвращающиеся с зимовок семьи распадаются перед началом распределения пар по гнездовым участкам (Литвин и др., 1999).
Едва ли можно усомниться в том, что прогресс в социальной эволюции позвоночных животных во многом достигался на пути усовершенствования способностей особей к долгосрочному запоминанию индивидуальных признаков многих субъектов. Бесспорно, что крупным гусеобразным удалось продвинуться по этому пути дальше многих других птиц.
И все же присущий гусеобразным уровень социальной компетенции, на мой взгляд, не стоит переоценивать. Многие черты общественной жизни этих птиц, представляющиеся свидетельством их незаурядного интеллекта, в не менее яркой форме проявляются и у других видов. Например, супружеские пары зебровых амадин (Taeniopygia guttata) тоже не распадаются по много лет вопреки высокой степени стайности, колониальному гнездованию и регулярным переселениям из одной в колонии в другую. У африканских белолобых щурок (Merops bulockoides), ежегодно меняющих место колонии, наблюдается многолетнее постоянство сложно устроенных семейных кланов, включающих десятки особей разного пола и возраста. Волнистые попугайчики (Melopsittacus undulatus) после долгой разлуки узнают супруга не только при личной встрече, но и по изображению на слайде. Таким образом, умением долго хранить в памяти облик партнера (или многих партнеров) на фоне бесконечного калейдоскопа анонимных образов соплеменников наделены не только гусеобразные. Можно привести множество примеров коммунально-кооперативных систем размножения, которые, на мой взгляд, служат наиболее убедительным свидетельством способности многих птиц (султанок, дятлов, щурок, тимелий, завирушек и пр.) формировать очень сложную социальную среду, требующую от особей высочайшего уровня социальной компетенции и умелого социального маневрирования (Иваницкий, 2001).
С учетом сказанного можно предположить, что социальная организация «гусиного» типа с присущими ей многосемейными стаями, перманентным браком и длительными связями родителей и потомства в истории эволюции сообществ гусеобразных является наиболее древней. Биологический смысл такого устройства социума состоит в обеспечении наиболее комфортных условий для развития молодняка в условиях жесткого соперничества в многочисленных и плотных стаях крупных, физически сильных и хорошо «вооруженных» птиц.
Главным побудительным мотивом к cоставлению этого обзора стало мое многолетнее общение с Е.Н.Гуртовой, К.Е.Литвиным, А.Б.Поповкиной, Н.Д.Поярковым, Е. В. Сыроечковским и С.П.Харитоновым, которые не только делились со мной результатами своих полевых исследований поведения гусеобразных, но и предоставили мне возможность ознакомиться со многими труднодоступными литературными источниками. В процессе работы автор пользовался частичной финансовой поддержкой фондов «Интеграция» и «Университеты России – фундаментальные исследования».
Литература
Базиев Д. Х. 1978. Улары Кавказа. Экология, морфология, эволюция. Л.: 1–125.
Баранюк В.Н. 1999. Гнездование белых гусей в условиях дефицита гнездо-вой территории. – Казарка, 5: 161–174.
Иваницкий В.В. 1997. Воробьи и родственные им виды зерноядных птиц: поведение, экология, эволюция. – М., Товаpищество научных изданий КМК: 1–148.
Иваницкий В.В. 2001. Социальное поведение и коммунальное гнездование птиц. – Соросовский образовательный журнал, 7: 29–35.
Кречмар А.В. 1995. Шилохвость в среднем течении Анадыря. – Орнитоло-гия, 26: 50–61.
Литвин К.Е. 1985. Взаимосвязь стайного поведения и колониальности у птиц. – Теоретические аспекты колониальности у птиц. М., изд-во МО-ИП: 83–86.
Литвин К.Е., Гуртовая Е.Н., Сыроечковский Е.В. 1999. Структура популя-ции малых лебедей в период размножения. – Казарка, 5: 185–202.
Панов Е.Н. 1983. Поведение животных и этологическая структура популя-ций. – М., Наука: 1–423.
Потапов Р. Л. 1987. Отряд Курообразные. – Птицы СССР. Курообразные, журавлеобразные. Л., Наука: 7–260.
Симкин Г.Н. 1988. Происхождение и эволюция колониальности у птиц. – Орнитология, 23: 36–51.
Сыроечковский Е.В. 1976. Особенности поведения белых гусей (Anser caerulescens) в гнездовой период. – Зоол. журн., 55 (10): 1485–1494.
Сыроечковский Е.В. 1979. Подкладывание белыми гусями яиц в чужие гнезда. – Зоол. журн., 58 (5): 1033–1038.
Сыроечковский Е.В. 2000. Гипотеза о происхождении гуся-белошея. – Ка-зарка, 6: 45–57.
Харитонов С.П. 1998. Этологическая структура локальной группировки в коло-нии белых гусей (Anser caerulescens). – Зоол. журн., 67 (10): 1530–1537. 120 121
Afton A.D. 1985. Forced copulations as reproductive strategy of male lesser scaup: a field test of some predictions. – Behaviour, 92: 146–167.
Barash D.P. 1978. Sociobiology of rape in mallards (Anas platyrhynchos): re-sponse of the mated male. – Science, 197: 788–789.
Beauchamp G. 1998. The relationship between intra- and interspecific brood amalgamation in waterfowl. – Condor, 100: 153–162.
Black J.M., Owen M. 1987. Determinants of social rank in goose flocks: acquisi-tion of social rank in young geese. – Behaviour, 102: 129–146.
Black J.M., Owen M. 1989. Parent-offspring relationships in wintering barnacle geese. – Animal Behaviour, 37: 187–198.
Ely C. R. 1993. Family stability in Greater White-fronted Geese. – Auk, 110 (3): 425-435.
Feduccia A. 1978. Presbyornis and the evolution of ducks and flamingos. – Amer. Sci., 66: 298–304.
Gauthier G. 1988. Territorial behaviour, forced copulations and mixed reproduc-tive strategy in ducks. – Wildfowl, 39: 102–114.
Johnsgard P.A. 1965. Handbook of waterfowl behaviour. – London. Constable Co., Ltd.: 1–378.
Johnsgard P.A. 1978. Ducks, geese, and swans of the world. – Univ. of Nebraska Press. Lincoln and London: 1–404.
Jutglar F. 1992. Anatidae. – Handbook of the birds of the world. Vol.1. Ostrich to Ducks: 574–624.
Kear J., Steel T.H. 1971. Aspects of social behaviour in the Blue Duck. – Notor-nis, 18: 187–198.
Lack D. 1974. Evolution illustrated by waterfowl. – Oxford–London–Edinburg, Blackwell Sci. Publications: 1–93.
Livezey D., Humphrey J. 1985. Territoriality and interspecific aggression in steamer-ducks. – Condor, 87: 134–136.
Nuechterlein T.G., Storer F. 1985. Aggressive behaviour and interspecific killing by flying steamer-ducks in Argentina. – Condor, 87: 97–98.
Perrins C.M. (ed.) 1990. The Illustrated Encyclopaedia of Birds. – London, Mar-shall Editions Developments Limited.: 1–420.
Pierce R. J. 1996. Family Recurvirostridae (stilts and avocets). – Handbook of the birds of the world. Vol.3. Hoatzin to Auks: 332–347.
Raveling D.G. 1970. Dominance relationships and agonistic behaviour of Canada geese in winter. – Behaviour, 37: 291–319.
Savard J.-P. 1978. Causes and functions of brood amalgamation in Barrow’s Goldeneye and Bufflehead. – Canadian Journ. Zool., 65: 1548–1553.
Scott D.K. 1980. Functional aspects of prolonged parental care in Bewick’s Swans. – Animal Behaviour, 28: 938–952.
Seymour N. R., Titman R. D. 1978. Changes in activity patterns, agonistic behav-iour, and territoriality of black ducks (Anas rubripes) during the breeding sea-son in Nova Scotia tidal marsh. – Can. J. Zool., 56 (8): 1773–1785.
Siegfried W.R. 1976. Social organization in Ruddy and Maccoa Ducks. – Auk, 93: 560–570.
Stewart G.R., Titman R.D. 1980. Territorial behaviour by prairie pothole blue-winged teal. – Canadian Journ. Zool., 58: 639–649.
Veselovsky Z. 1975. Comparative ethology in the order Anseriformes. – Activ. Nerv. Sup. Praha., 17: 70–71.
Warren S.M., Fox A.D., Walsh A., Sullivan P. 1993. Extended parent-offspring relationships in greenland white-fronted geese (Anser albifrons flavirostris). – Auk, 110: 145–148.
Welsh D., Sedinger J. 1990. Extra-pair copulations in Black Brant. – Condor, 92: 242–244.
Источник Казарка. 2002. Т.8
Рекомендуем прочесть
 «Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве
«Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве Как сыграл «фактор Пиночета» на президентских выборах в Чили
Как сыграл «фактор Пиночета» на президентских выборах в Чили Восприятие шумов
Восприятие шумов Винтовка — это праздник: откуда в Колумбии тысячи детей-солдат и какая их ждет судьба
Винтовка — это праздник: откуда в Колумбии тысячи детей-солдат и какая их ждет судьба Отбор адаптивной и неадаптивной пластичности в экспрессии генов при приспособлении анолисов к городскому «острову тепла»
Отбор адаптивной и неадаптивной пластичности в экспрессии генов при приспособлении анолисов к городскому «острову тепла»![Разоблачение [The Grayzone]: британская разведка спланировала подрыв Керченского моста](http://www.socialcompas.com/wp-content/uploads/2022/10/Snimok-ekrana-ot-2022-10-12-20-50-44-150x150.png) Разоблачение [The Grayzone]: британская разведка спланировала подрыв Керченского моста
Разоблачение [The Grayzone]: британская разведка спланировала подрыв Керченского моста Человеческая деятельность формирует зимнюю экологию перелётной птицы, славки-черноголовки
Человеческая деятельность формирует зимнюю экологию перелётной птицы, славки-черноголовки Иван Владимиров — художник-перевертыш, который славил Сталина и клеймил военный коммунизм
Иван Владимиров — художник-перевертыш, который славил Сталина и клеймил военный коммунизм
