

Деревья и близость видов
Содержание
«Близость» видов на филогенетическом дереве отражается разными способами, зависящими, в том числе, от наших представлений, как следует реконструировать их общую эволюционную родословную. Т. е. среди всех сходств выделить именно те, что отражают родство, и затем перевести их в древовидный граф. Сейчас в реконструкциях филогенеза доминирует кладистический подход, который принимает в расчёт только один тип филогенетических событий — дивергенцию видов («узлы ветвлений» на рисунке 1) но не все прочие (в том числе непоказанное разновременное происхождение нескольких видов А, Б, В… от предкового вида Х, существующего весь этот период).
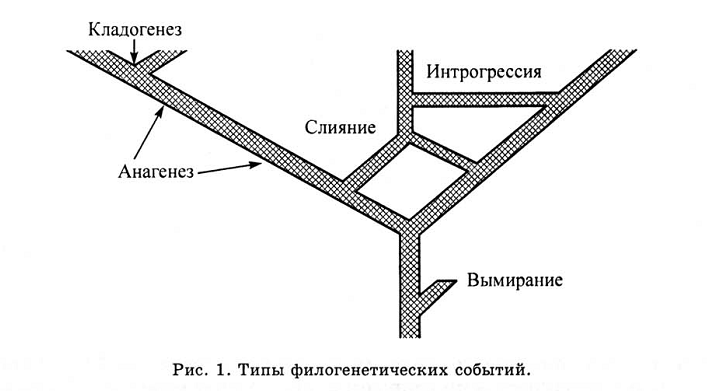 Кладизм — это самоограничение, отказ от анализа прочих аспектов филогенеза для концентрации на дивергенции «ветвей», считающейся самой главной. Этот же принцип лежит в основе молекулярной систематики — построению дерев по текстам ДНК (отдельным «словам» и даже «окончаниям» — рестрикционный анализ — избранным в связи с их определённым «значением», т. е. генам).
Кладизм — это самоограничение, отказ от анализа прочих аспектов филогенеза для концентрации на дивергенции «ветвей», считающейся самой главной. Этот же принцип лежит в основе молекулярной систематики — построению дерев по текстам ДНК (отдельным «словам» и даже «окончаниям» — рестрикционный анализ — избранным в связи с их определённым «значением», т. е. генам).
Хотя эта последняя де факто относится уже к фенетике, направлению, при зарождении бывшему оппонентом кладизма.
Так или иначе, кладизм, как всякое упрощение реальности до модели, «выхватывающей» одну из системных связей, и опускающей прочие («подход Прокруста» по Расницыну), имеет свои сильные и слабые стороны, интенсивно дискутируемые специалистами (см. 1-2). И любят его за первые, а не вторые, о которых не надо забывать — дальше речь именно о них. Скажем, А.П. Расницыным на юбилейной конференции, посвященной 150-летию Русского энтомологического общества «Теоретическая и прикладная энтомология — прошлое, настоящее и будущее»,
«…докладывались результаты исследования по согласованию с палеонтологическими данными филогений [перепончатокрылых] полученных 3 группами методов: 1) традиционным, 2) морфологической кладистикой, 3) молекулярной кладистикой. Насколько я понял, брались филогении насекомых (каких групп — не уловил). Для сопоставимости филогений часть групп (которые присутствовали не во всех кладограммах) выбрасывались. Рассчитывался некий коэффициент соответствия дендрограммы порядку возникновения таксонов, фиксируемому палеонтологией. Кладограммы (всего 38) брались из работ, опубликованных в 2001-2008 гг. Думаю, это будет опубликовано и желающие смогут познакомиться с методикой сравнения более детально.
Результат, по словам докладчика, удивил его самого. Лучшие показатели соответствия палеонтологическим данным практически во всех случаях (т.е. для всех групп) имели филогении, построенные традиционным способом. Затем шли кладограммы, построенные по морфологическим признакам, и худшие показатели имели, как ни странно, молекулярные кладограммы. Надо заметить, что работы брались достаточно современные, когда молекулярная кладистика уже достаточно развилась методически: появился maximum likelihood и прочие продвинутые методы».
Соответственно, в рамках кладизма (в т.ч. по молекулярным деревьям) бонобо и шимпанзе суть «ветвление» филогенетической линии, сестринской относительно ведущей к нам, грешным, и поэтому равно родственны Homo sapiens (рис.2).
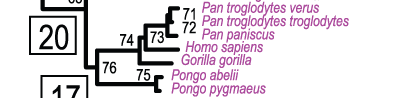
Рисунок 2. Сегмент «кроны» молекулярного дерева 186 видов приматов, с двумя внешними группами. Рис.2. в Perelman et al., 2011.
Это правда, но, как обычно, не вся, и самая неинтересная с точки зрения сходств и различий в биологии с поведением наших трёх видов. В других подходах (эволюционная систематика) филогенетическая близость/отдалённость связана не только с порядком отхода ветвей, как на представленном молекулярном древе (отдельная тема, что морфологическая и молекулярная эволюция в норме протекают по разному), но и с анагенетическим компонентом — дальностью расхождения ветвей друг от друга. См. п. 4.1.2.2 «Введения в филогенетику» И.Я. Павлинова и рис.3, где анагенетический компонент отложен по оси абсцисс.
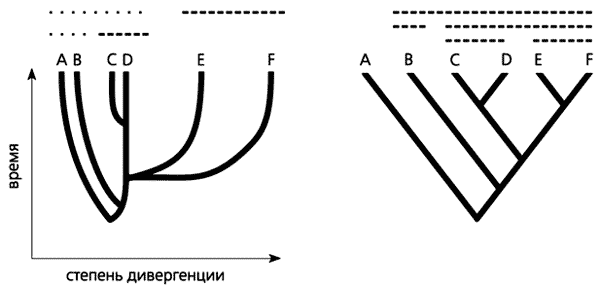
Рисунок 3
И этот момент нельзя не учитывать при переводе филогении в классификацию, иначе все тетраподы будут одним из семейств остеолепиформных рипидистий, или завроморфы — частью радиации лабиринтодонтов (рис.4).
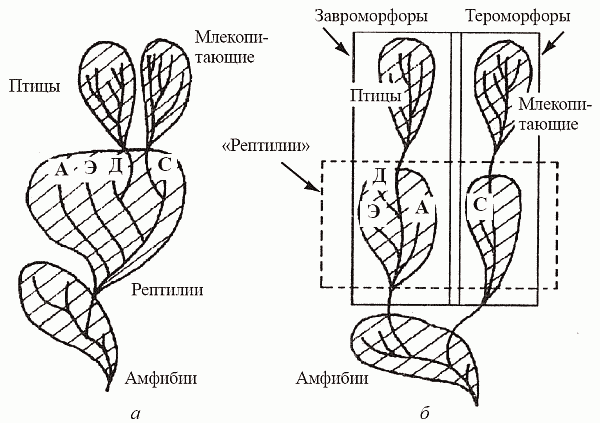
Рисунок 4. Источник. К.Ю.Еськов. История Земли и жизни на ней.
Сказанное верно и для филогенетического дерева антропоидов, где, огрублённо, одна ветвь ведёт к нам, грешным, а другая — к обыкновенным шимпанзе и бонобо. Сравнивая сходства-различия, из них именно бонобо по большинству признаков сближается с человеком — грацильный скелет, красные губы, общие «ювенильные» пропорции черепа и костяка1, долго сохраняющаяся «детскость характера» с присущими ей — что у нас, что у бонобо — «автоматическим» альтруизмом и тенденцией к равенству.См. рис.5.
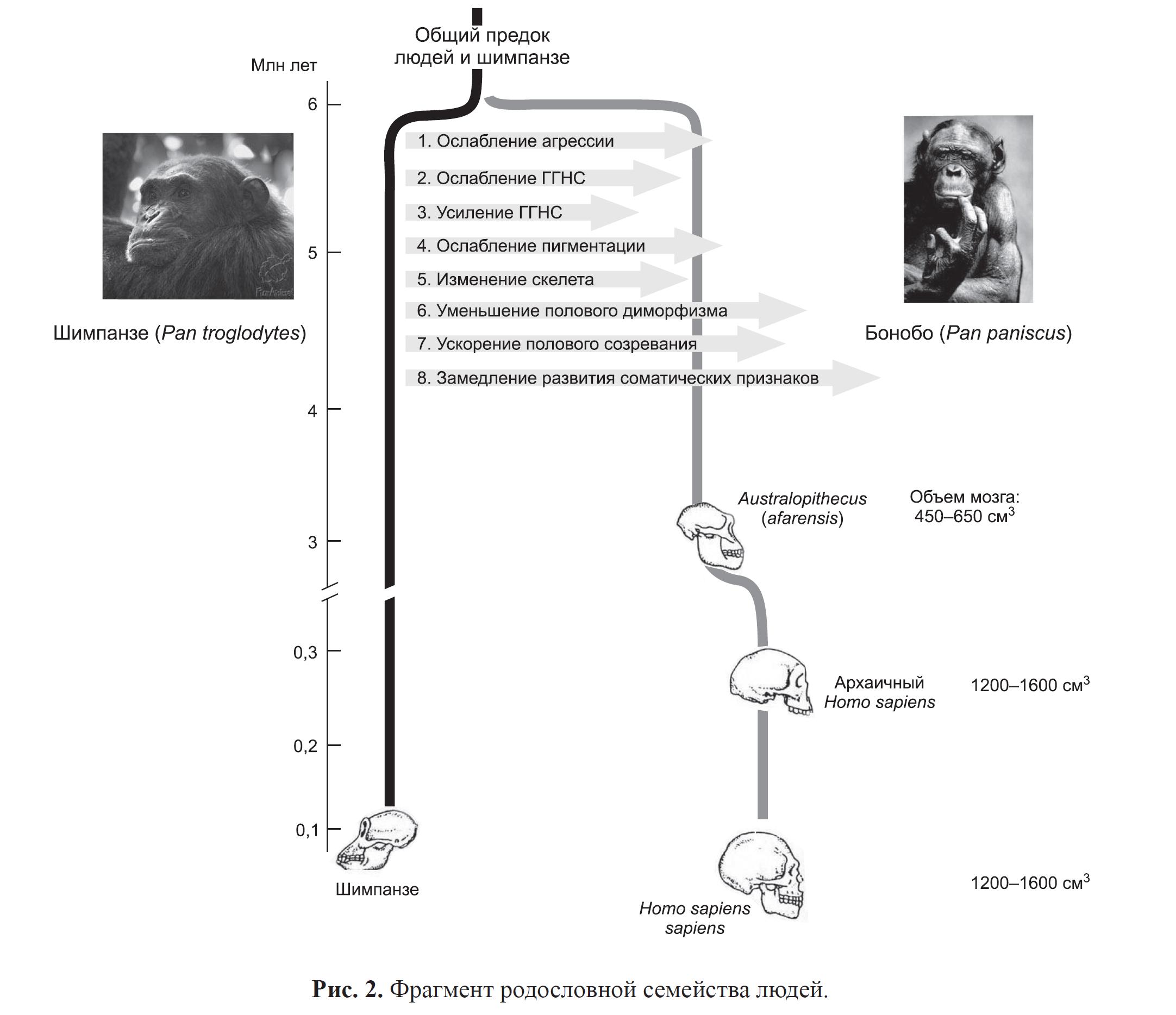
Рисунок 5. Источник. Трут, 2008
Всё это родило т.н. «гипотезу Зильмана», что общий предок людей и шимпанзе был сходен именно с бонобо (Зильман считал, что также и гориллы). С большим шимпанзе нас сближают, в общем, два свойства — использование орудий и агрессивность, доводящая до смертоубийств, и оба нехарактерны для бонобо, см.ниже.
«-зации» и доместикация
Признаки, сближающие нас с бонобо, не уникальны, но закономерно повторяются (большей частью, не все) в ряде других филогенетических ветвей класса млекопитающих, вообще не имеющих отношения к гоминидам или приматам. Это ровно те «веточки», что связаны с доместикацией собак, лисиц, коз, лошадей и пр. видов, а соответствующие признаки отделяют домашние формы от их диких предков и пр. родичей: большее миролюбие, продлённый период обучения, исчезновение стресс-реакции на новизну, повышенная исследовательская активность. Важно подчеркнуть, что параллельное развитие однотипных изменений морфологии, гормонального состояния, в поведении и на уровне генов вполне закономерно и представляет собой частный случай т.н. «-заций» (подробно изученных палеонтологами на примере маммализации териодонтов, орнитизации теропод, ангиоспермизации голосеменных и пр.) — независимого приобретения одних и тех же новшеств в параллельно развивающихся филогенетических ветвях, обычно в рамках прогресса организации. См. рассказ К.Ю.Еськова про автокатализ в запуске микро- и макроэволюционных процессов.
См. обзор Л.Н. Трут (2007) про каскад изменений морфологии, физиологии и поведения, запускающихся независимо во всех этих неродственных линиях вследствие т.н. дестабилизирующего отбора Д.К. Беляева. Основные компоненты каскада представлены на рис.6.
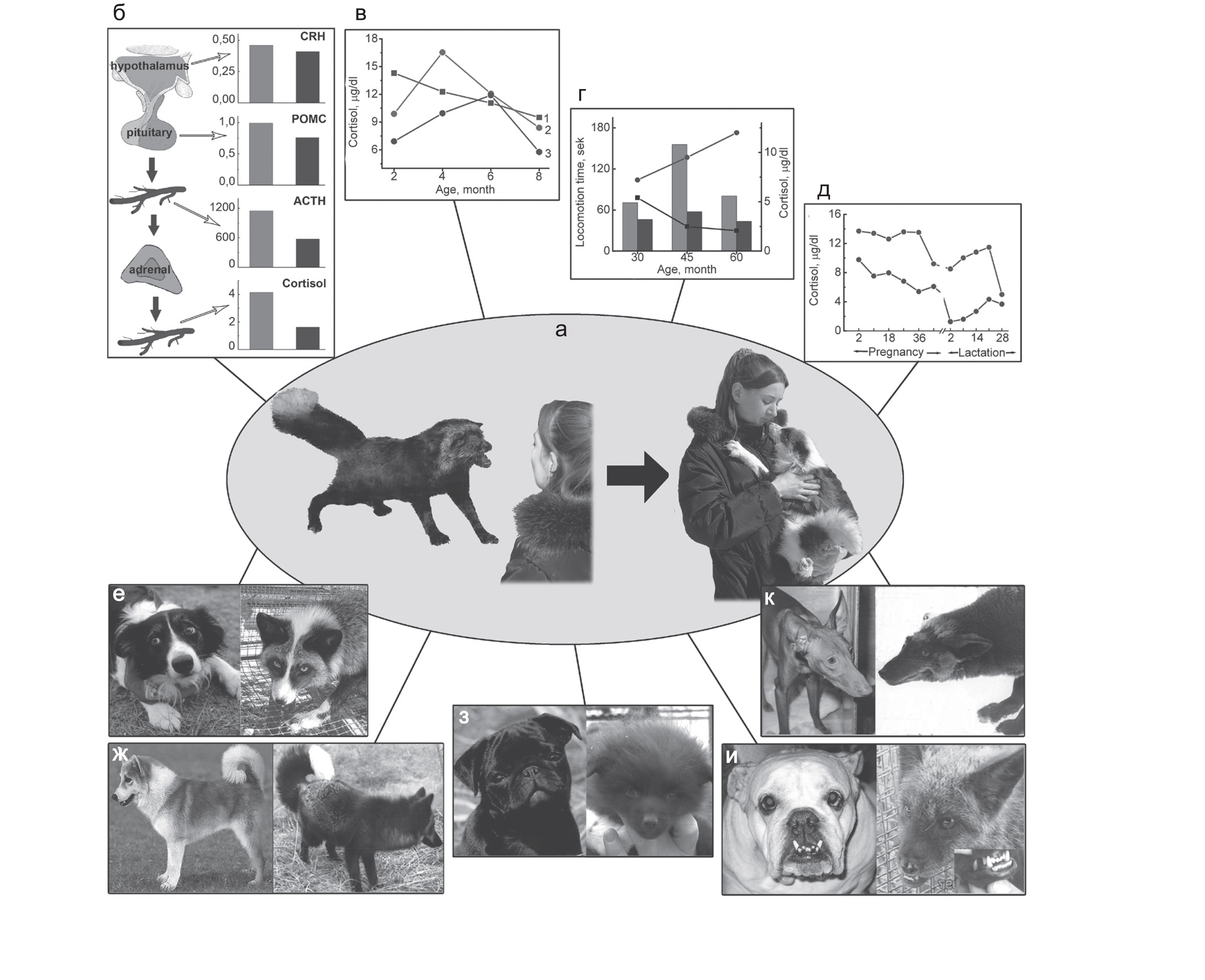
Рисунок 7. Схематическое представление основных результатов эксперимента Д.К. Беляева. Из: Трут, 2008. а – трансформация поведения лисиц в ходе отбора: слева – лисица из неселекционируемой по поведению промышленной популяции. Агрессивная реакция на человека наиболее типична для этих лисиц: справа – лисица из экспериментальной доместицируемой популяции. Поведение, характерное для собак, сформировалось у этих лисиц в результате длительного отбора; б–д – показатели функциональной активности гипоталамо-гипофизарно-надпочечниковой системы (ГГНС); б – экспрессия гена (или уровень мРНК) кортикотропин-рилизинг гормона (CRH) в гипоталамусе и пропиомеланокортина (POMC) в гипофизе; уровни адренокортикотропного гормона (АКТГ) и кортизола в плазме крови у неселекционируе- мых (слева) и ручных (справа) серебристо-черных лисиц; в – возрастные изменения уровня кортизола в плазме крови у неселекционируемых и ручных серебристо-черных лисиц. 1, 2 – неселекционируемые лисицы: 1 – проявляющие агрессивную реакцию на человека, 2 – не проявляющие ее; 3 – ручные лисицы с высокой оценкой доместикации; г – суммарное время двигательной активности – индикатора исследовательского поведения и уровень кортизола в плазме крови у неселекционируемых и ручных серебристо-черных лисиц в возрасте 1–2 месяца: двигательная активность изоб- ражена графически; нижняя линия – неселекционируемые, верхняя – ручные; уровень кортизола – в виде гистограммы (левые столбики – неселекционируемые, правые – ручные); д – уровень кортизола в плазме крови у серебристо-черных лисиц при беременности и лактации: (верхняя линия – неселекционируемые, нижняя – ручные); е–к – некоторые морфологические изменения, имеющиеся у собак и возникшие у лисиц доместицируемой популяции; е – сходный характер депигментации волосяного покрова у собак и лисиц: слева – собака бордер-колли; справа – ручная лисица; ж – манера держать хвост свернутым в кольцо: слева – исландская собака; справа – ручная серебристо-черная лисица; з – у отдельных ручных лисят повисают уши и укорачивается лицевой череп, что характерно для некоторых пород собак: слева – собака мопс; справа – ручной лисенок; к – удлинение лицевого черепа у некоторых пород собак и у ручных лисиц: слева – фараонова собака; справа – лисица; и – относительное удлинение нижней челюсти (пе- рекус), наблюдаемое у английского бульдога, происходит и у некоторых ручных лисиц: слева – английский бульдог; справа – ручная лисица.
В другой статье (Трут, 2008) показано, что сходства человека и бонобо а) носят системный характер и б) также могут быть поняты как доместикация, поскольку представляют собой органичную часть синдрома признаков, связанных с этой последней.
«Следует еще раз отметить тот факт, что варьирование скелетных параметров у лисиц так же, как и у собак, в основном связано с соотношением ширины и длины костей. Оно отражает, вероятно, разную скорость перехода ювенильной формы, для которой характерны более широкие и короткие кости, во взрослую. То же самое можно сказать и о варьировании поведенческих компонент. Они также отражают темпы трансформации детской эмоциональной привязанности к человеку во взрослое поведение. Все это наводит на мысль, что если и существуют локусы, корегулирующие поведение и морфологию, то они, как уже обсуждалось, вероятно, обладают более общей функцией регуляторов скоростей развития.
Свое понимание генетико-эволюционных основ доместикации Д.К. Беляев связывал с эволюцией человека. Он подчеркивал, что в эволюции предков современного человека и в эволюции домашних животных существуют общие закономерности. Они становятся более объяснимыми, если принять, что предоминантную роль в ходе антропогенеза так же, как и в ходе доместикации, играл отбор по поведению (Беляев, 1981). Как известно, наиболее значительные этапы эволюции семейства гоминид произошли около 5–6 млн лет назад. Они привели к разделению ветви людей и шимпанзе от общего предка (рис. 4). Возможным прототипом основателя родословной людей некоторые эволюционные антропологи2 считают бонобо, или карликового шимпанзе (Pan paniscus), который удивительно близок австралопитеку (Australopitecus) – самому древнему нашему предку.
Но принципиально то, что между шимпанзе и бонобо выявлен тот же комплекс поведенческих и морфофизиологических различий (Wrangham, Pilbeam, 2001), который выявлен нами между фермерскими и домашними лисицами (рис. 4). Этот комплекс изменений представляет собой также неотенический или педоморфный комплекс.
Механизмом же, продуцирующим неотению у бонобо, как и у лисиц, мог быть отбор на «миролюбие», на приручаемость. Такой отбор удерживает во взрослой жизни не только ювенильные черты поведения, но, по-видимому, и многие другие системы и признаки. Все это дало основание полагать, что бонобообразный предок семейства гоминид попал в такие экологические условия, в которых экстремальным было давление отбора на ослабление агрессии. Это экстремальное селекционное давление и могло сыграть решающее значение в эволюционном отделении бонобо от шимпанзе.
Отбору по поведению Д.К. Беляев отводил решающую роль во всей дальнейшей эволюции отделившейся ветви. Он отмечал, что эта эволюция характеризовалась теми же специфическими особенностями, что и эволюция домашних животных, прежде всего, огромными темпами и масштабами морфофизиологических преобразований (Беляев, 1981). Особенно отмечают драматическое увеличение размера мозга, объем которого от стадии австралопитека до современного разумного человека увеличился от 350 см3 до 1,400–1500 см3 (рис. 5). В настоящее время имеются данные, указывающие на то, что одним из существенных молекулярных коррелятов ускоренной фенотипической эволюции размеров мозга может быть ускоренная эволюция мозгоспецифических генов, особенно генов, вовлекаемых в детерминацию размеров мозга, например, ASPM и MCPH (Dorus et al., 2004; Evans et al., 2005; Mekel-Bordov et al., 2005).
Имеются доказательства в поддержку адаптивного характера ускоренной эволюции мозгоспецифических генов, т. е. их ускоренная эволюция может быть результатом экстремального селекционного давления на них. Это в свою очередь может быть иллюстрацией того, что отбор порождает изменчивость в направлении своего действия».
Вестник ВОГиС. 2008. Т.12. №1/2.
См. также более позднее (и подробное) обоснование гипотезы самодоместикации, чем цитированное Трут, со схемой последовательных этапов процесса. Другое дело, что у бонобо (в отличие от человека) мы видим не все поведенческие изменения, связанные с доместикацией.
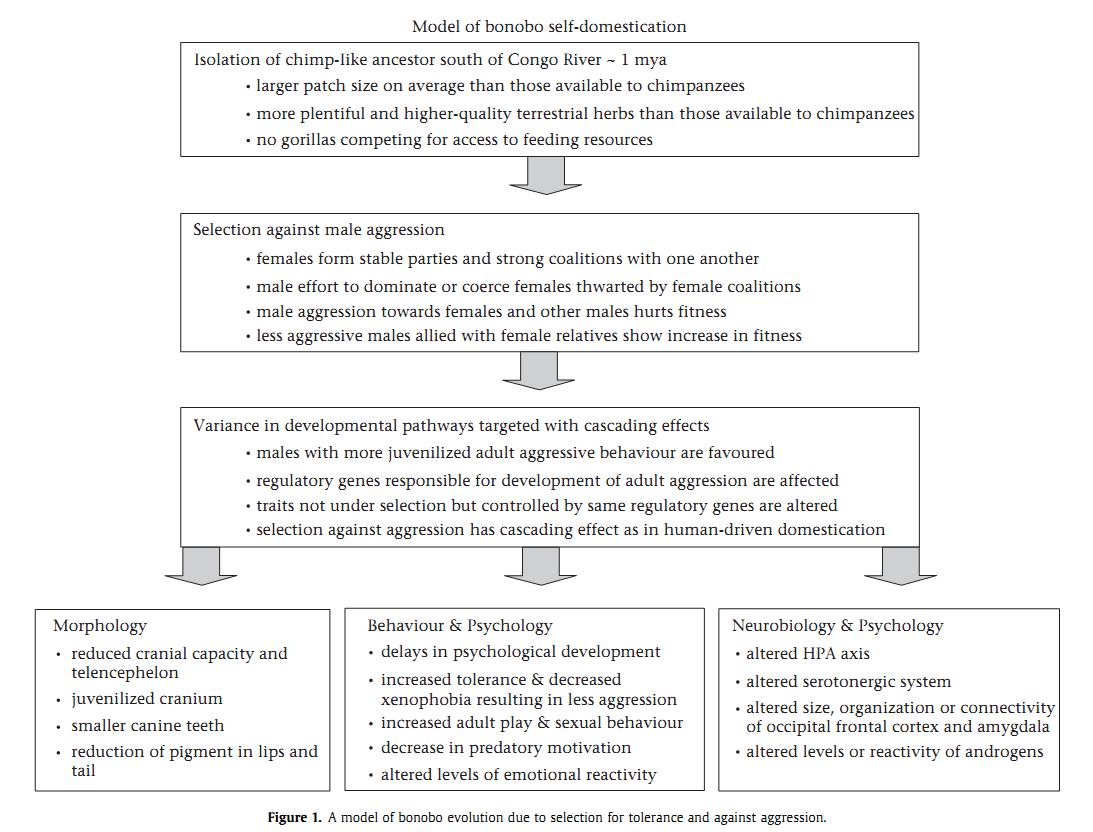 Скажем, он, как и обыкновенный шимпанзе, отнюдь не показывает способность «считывать» и использовать социальные «подсказки», в полной мере присутствующую не то что у собак c лошадьми (и одомашненными лисицами — см.рис.8), но даже у коз. Может быть вследствие эгалитарности его социума (а большой шимпанзе — из-за рассредоточенности): данное свойство развивается при абсолютном превосходстве «хозяина», воспринимаемого как доминант, и непрерывно управляющего животным.
Скажем, он, как и обыкновенный шимпанзе, отнюдь не показывает способность «считывать» и использовать социальные «подсказки», в полной мере присутствующую не то что у собак c лошадьми (и одомашненными лисицами — см.рис.8), но даже у коз. Может быть вследствие эгалитарности его социума (а большой шимпанзе — из-за рассредоточенности): данное свойство развивается при абсолютном превосходстве «хозяина», воспринимаемого как доминант, и непрерывно управляющего животным.
Рисунок 8 из: Трут, 2007
Так или иначе, если всерьёз, не декларативно рассматривать биологическую сторону нашего поведения, исходя из существующих эволюционных сценариев его происхождения, то а) телесно наш предок, то бонобо действительно ближе всего к нам сравнительно с обыкновенным шимпанзе (т.е. он в «ветви шимпанзе» анцестральнее, а большой шимпанзе — продвинутее), б) признаки сходства с бонобо, если даже не унаследованы от общего предка, а возникли независимо в «человеческой» ветви, их параллельное формирование закономерно и представляет собой частный случай закономерно возникающих параллелизмов («-заций»), в) имеющиеся у человека признаки сходства с большим шимпанзе развивались на иной основе, нежели у него.

Сведение вместе разнородных данных о когнитивных способностях обоих шимпанзе, как и агрессии второго вида в сравнении с таковой наших предков хорошо ложится в эту гипотезу и опровергает противоположную ей (об «общей природе» орудийной деятельности и агрессии у нас и Pan troglodytes).
Сходства/различия в использовании орудий
Большой шимпанзе, как известно, более сходен с человеком в использовании орудий — в сравнении с бонобо он много чаще делает это в природе, сами орудия разнообразней и сложней, отдельные сообщества резко дифференцированы по традициям орудийной деятельности («культура») и пр.
Однако недавно выяснилось, что это сходство — разного происхождения «у нас» и «у них». Kathelijne Koops с соавт. впервые исследовала в сравнительном плане возможности орудийной деятельности в природных сообществах шимпанзе и бонобо Уганды и Конго (экологическая необходимость, социальные предпосылки использования орудий, проистекающие из разных форм манипуляции), а также частоту попыток использования орудий у молодых обезьян с вероятность перерастания в устойчивый навык.
Прежде всего, различия в использовании орудий не объясняются экологическими различиями этих двух видов. На участках обитания бонобо имеют доступ ко всё тем же орудиям (камни, палочки для «ужения» термитов) и кормовым объектам (ореховые деревья, годные термитники гнёзда), что и шимпанзе — см. рис.1 статьи.
Разница в социальности также не объясняет различий. Уже появившимся инновациям шимпанзе учатся друг у друга, и новая орудийная практика, как любая другая традиция, распространяется по социальным связям, т. е. устанавливается тем быстрей и надёжней, чем их больше. Однако у молодых бонобо больше социальных партнёров, чем у молодых больших шимпанзе; у них же больше особей находится в непосредственной близости, когда они кушают (поскольку миролюбивей и реже задают трёпку). Они дольше остаются с матерями, а именно самки у антропоидов более изобретательны в использовании орудий и являются главными агентами их передачи. Однако же молодые шимпанзе много чаще манипулируют с объектами, чем бонобо, и играют с объектами в одиночку.
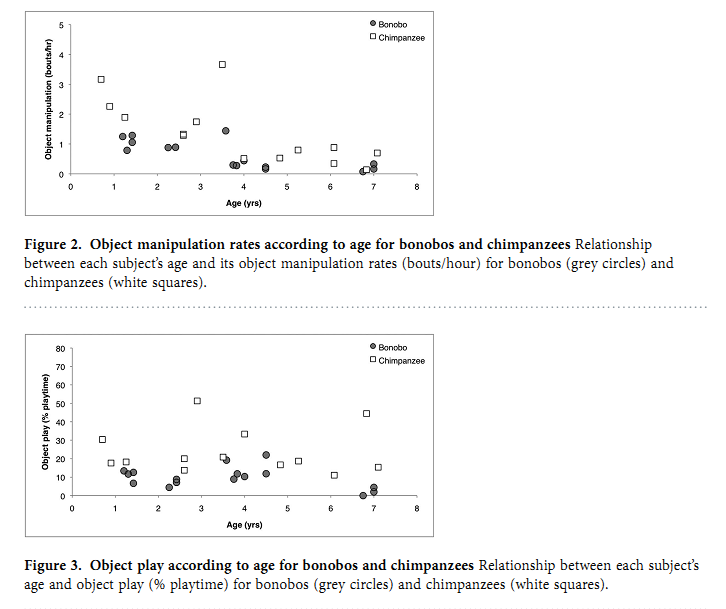 Эти различия между видами появляются в очень раннем возрасте — вроде бы раньше, чем начнётся обучение от других, так что склонность больших шимпанзе к использованию орудий можно (пока) считать врождённой (и, соответственно, несклонность бонобо). Важно подчеркнуть особенности метода исследований — не предрешая заранее, что есть «использование орудий», что нет, фиксировать все акты манипуляции с объектами в разных контекстах, не только кормодобывания, и дальше смотреть, что «вырастает» в орудийную деятельность, что нет.
Эти различия между видами появляются в очень раннем возрасте — вроде бы раньше, чем начнётся обучение от других, так что склонность больших шимпанзе к использованию орудий можно (пока) считать врождённой (и, соответственно, несклонность бонобо). Важно подчеркнуть особенности метода исследований — не предрешая заранее, что есть «использование орудий», что нет, фиксировать все акты манипуляции с объектами в разных контекстах, не только кормодобывания, и дальше смотреть, что «вырастает» в орудийную деятельность, что нет.
Koops с соавторами показала, что с очень раннего возраста деятельность шимпанзе ориентирована на объект и «полезный результат» от взаимодействия с ним. Бонобо же — не на объект, а на социального партнёра (и особенно его эмоции в связи с собственными поступками и другими событиями вокруг, по которым они оценивают чужое поведение и регулируют своё, т. е. вполне «человеческим способом», см. Clay, de Waal, 2013).
Межполовые различия
Также забавно описанное ими межполовое различие в объект-ориентированной деятельности у молодых больших шимпанзе в переходный период ко взрослой жизни, когда орудия используются по-настоящему. Самцы сильно чаще манипулировали с объектами, но в основном для прикола и отдыха, во время игры. Молодые самки делали это реже (и особенно редко в игре), но с более разнообразными способами манипуляции, включая удары объектами и по ним, их поломку и переноску.
Всё это лучше подготавливает самок к реальной орудийной деятельности, где они превосходят самцов. Соответственно, самки быстрей учатся использовать орудия для реальных задач.
Причём если количественные различия в манипуляции между самцами и самками шимпанзе в целом фиксируются также у (современных европейских) детей, то качественные — нет. Почему надо думать, у человека, в отличие от шимпанзе, орудийная деятельность связана не с «биологическим», а с «социальными» — необходимостью решения задач на плановое изготовление и использование орудий, вставшая некогда перед сообществами наших далёких предков, телесно сходных именно с бонобо.
Опровергли ли Выготского?
Если читать только популярные пересказы исследований, можно думать, что этот вывод опровергнут в работе Reindl et al., 2016, вместе с мнением Выготского, что новые навыки использования орудий у человека «воспринимаются лишь через обучение, а не изобретаются в требующей этого ситуации в силу врождённой склонности к орудийной деятельности». Это мнение пересказывающего работу в ScienceDaily вполне неверно — с одной стороны, теория Выготского требует не отрицательной, а положительной связи между навыками, воспринятым из обучения и других форм социальной поддержки, и самостоятельно делаемым потом (иначе не стоило бы вводить понятие зоны ближайшего развития), с другой — шимпанзе, как мы видели, перенимают чужие «изобретения» сообразно связям в сообществе, а не собственной активности такого рода.
Соответственно пересказывающий считает, что в Reindl et al., 2016 показано, что 2,5-3-х летние дети (!, с нормально развитой речью, из семей, нормально обеспеченных игрушками и пр.), которым предложили решать задачи, экологически актуальные для шимпанзе, и требующие использования орудий, показывают такую же врождённую склонность к орудийной деятельности, как и эти обезьяны.
 Сами авторы в утверждениях осторожней, и Выготского интерпретируют разумней, понимают, что скорей подтвердили, чем опровергли его; другое дело, что в работе есть ряд методических просчётов, делающих результат не вполне чётким и однозначно интерпретируемым.
Сами авторы в утверждениях осторожней, и Выготского интерпретируют разумней, понимают, что скорей подтвердили, чем опровергли его; другое дело, что в работе есть ряд методических просчётов, делающих результат не вполне чётким и однозначно интерпретируемым.
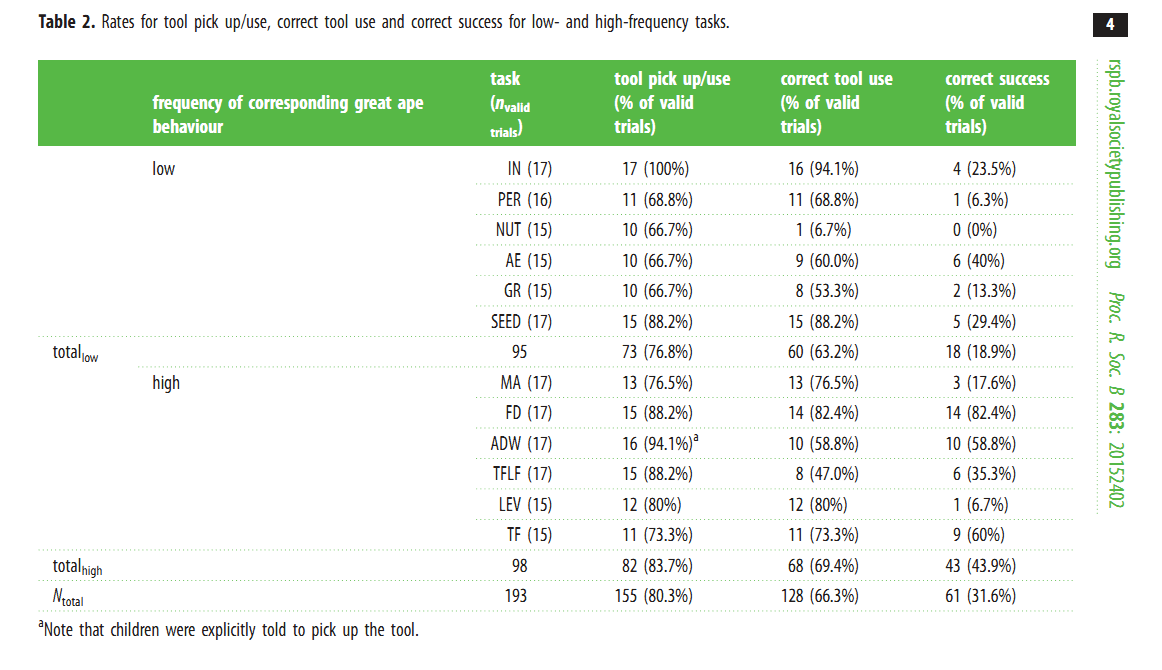
Авторы суммировали данные по разным видам инструментального поведения человекообразных обезьян, прежде всего в природных условиях и относительной их частоте/редкости (табл.1 статьи). К сожалению, они утверждают при этом, что свидетельства имитации и обучения у человекообразных обезьян редки (что просто неверно) и даже делают из этого вывод «tool use in these species is unlikely to be acquired via these mechanisms». Поскольку у человека именно эти факторы играют определяющую роль в приобретении индивидом навыков инструментального поведения, а также в накоплении их культурой, то для авторов встаёт вопрос о том, не ослабляется ли в линии, ведущей к человеку, способность к самостоятельному изобретению приёмов использования орудий.
В методике, с помощью которой авторы решали этот вопрос, есть как сильные, так и слабые стороны. В экспериментах участвовали дети 2-3,5 лет из Великобритании и Германии (очевидно, имевшие игрушки и обучаемые родителями манипуляциям с ними, а также понимающие речь и частично говорящие). На решение задач отводилось 1-3 минуты, притом, что у человекообразных обезьян в природе время никак не ограничено. При этом интересным моментом методики является попытка создать в лабораторных условиях аналог проблем, требующих в природе от человекообразных обезьян использования орудий. Использовались синтетические аналоги природных материалов, а также наклейки (а не пища) в качестве того, что требовалось добыть.
 Не совсем понятно, в какой мере использовалась речь для объяснения стоящей перед детьми задачи, во всяком случае, в описании методики говорится, что детям прямо указывали, чтобы они взяли в руки орудие. Не очевидно также, всегда ли дети могли сами понять, где находится стикер, хотя во многих случаях использовался пластик. Тем не менее, прямо вмешиваться в действия детей наблюдателю запрещалось.
Не совсем понятно, в какой мере использовалась речь для объяснения стоящей перед детьми задачи, во всяком случае, в описании методики говорится, что детям прямо указывали, чтобы они взяли в руки орудие. Не очевидно также, всегда ли дети могли сами понять, где находится стикер, хотя во многих случаях использовался пластик. Тем не менее, прямо вмешиваться в действия детей наблюдателю запрещалось.
Из 12 заданий с 11-ю успешно (в том числе с корректным использованием орудия) справилась хотя бы часть детей. Поскольку каждому ребёнку предъявлялись по 4 задания, некоторые успешно справились со всеми. Лучше выполняли задание дети старшего возраста, но разница между мальчиками и девочками была недостоверна, и соответствующие выборки были объединены. Это вроде бы указывает на сравнительно небольшое влияние внутренней речи, поскольку у девочек речь вообще в среднем развивается быстрее, но тут может быть много дополнительных влияющих факторов. Единственным заданием, при котором только один ребёнок корректно использовал орудие (но не преуспел) было разбивание пластикового «ореха» с наклейкой глиняным молотком. Я не исключаю, что причиной мог быть прямой запрет в большинстве семей на подобное поведение. В качестве подготовки к эксперименту детям предлагалось играть с деревянными палочками, использовавшимися во многих заданиях, в том числе ломать их, поскольку задания предполагали возможность модификации орудий, и требовалось снять запрет на это.
Самым интересным результатом эксперимента было то, что задачи, соответствующие характерному для человекообразных обезьян инструментальному поведению, решались много чаще (44%), чем те, которые соответствовали редко встречающемуся у них инструментальному поведению (19%). В определённой степени это может означать, что успешность в решении подобных задач может определяться задатками, унаследованными, как полагают авторы статьи, от общего предка человека и человекообразных обезьян. В то же время показательно, что высокоразвитая у человека способность к обучению и подражанию не привела к потере способности решать такие задачи самостоятельно, что говорит скорее в пользу концепции Выготского, чем против неё.
Что хорошо согласуется с наличием феномена избыточного подражания у детей, но не детёнышей шимпанзе (которые в соответствующих опытах в основном не подражают «модели», часть действий которой явно нефункциональна, а сообразив «по кёлеровски», как решить, делают это сами наиболее экономным способом). Почему в эволюции усиливалось избыточное подражание и, для его обслуживания, увеличился объём кратковременной памяти человека в сравнении с шимпанзе, как не для обслуживания необходимости выучивать пользование всё большим количеством орудий, ещё и с постоянно обновляющимся ассортиментом?
«Первобытные техники» у бонобо
Не обладая «внутренней» мотивацией к использованию орудий, бонобо, однако, сильно превосходят крупных сородичей в способности перейти к орудийной деятельности, когда ситуация требует многоступенчатого планирования действий, часть которых — изготовление орудий из подручного материала и применение их для добывания скрытой пищи. Скажем, они адекватно использовали орудия олдувайской эпохи (Toth et al., 1993). Сходным образом в недавней работе израильских авторов (Roffmann et al., 2015) имитировали условия, в которых развивалась орудийная деятельность у наших предков.

Бонобо, впервые изготавливающий дротик, дальше используемый в социальных взаимодействиях для атаки или защиты.
Думаю, так потому, что большие шимпанзе, как известно, ничего не могут сделать точно — ни ходить строем, ни бросать камни и палки в цель, слишком уж агрессивные и «дёрганные». Бонобо же спокойней и миролюбивей, проблемные ситуации решает легче; по той же причине, думаю, бонобо опережает шимпанзе в усвоении языка, если судить «по вершинам», Канзи и Панбанише (и особенно, в понимании синтаксической структуры языка, также задействующей навык планирования будущих действий).
Сперва была показана способность бонобо к изготовлению и использованию орудий по «первобытным технологиям» (в т.ч. авторами обсуждаемой работы, как раз на Канзи и Панбанише). Также фиксировали возможность использования орудий как дротиков в социальном контексте, для нападения и защиты.
Здесь же исследовалось, насколько бонобо в вольерных условиях способны совершать последовательность действий из n этапов для добывания корма, где подкрепляется лишь последний этап, а предыдущие подготовительные. Использовали 2 группы бонобо: 8 обезьян в Вуппертальском зоопарке, и 7 обезьян в «Убежище бонобо» в Айове, США, включавшей Канзи и Панбанишу: они росли в культурно обогащённой среде и имели доступ к лесу.
Перед обоими группами ставили одну и ту же проблему: надо добыть скрытый корм — либо спрятанный глубоко в землю (заваленный слоем камней в разных вариантах), либо скрытый в полости крупных трубчатых костей копытных (заполненных высушенными фруктами для имитации костного мозга), либо скрытый под твёрдой оболочкой закрытых капсул. Т.е. фактически вместе с задачей им предлагали «заготовки» к орудиям, позволяющим её решить; одновременно рядом с местонахождением корма набросали ещё «сырого материала» для потенциальных орудий — зелёные ветки и оленьи рога. См. рис. 1 статьи.
Уже через несколько дней бонобо из «Убежища…» стали изготавливать соответствующие задаче орудия в намеренной манере и плановым образом (рис. 2-6 статьи). Так, чтобы добраться до зарытого корма, им надо было сделать цепочку действий, включающих использование орудий. Сперва они откидывали камни, раскатывая их руками или используя палки как рычаги; рога также использовали как грабли или лопатки. Достигнув почвы, тыкали и копали короткой палкой, пока не получится ход в земле. Затем большей палкой расширяли и углубляли ход, используя её как лопату или совок; оленьи рога использовались в том же процессе как шило или мотыга. Наконец, они доставали корм, используя самую длинную ветку как шест и рычаг. См. видео
https://www.youtube.com/watch?v=89rNGhGv3ys
Работа Роффмана c соавт. — первое доказательство, что существа, не относящиеся к нашей филогенетической линии, могут использовать технологии однотипные использовавшимися древнейшими из наших предков — австралопитеками и ранними Homo (и обладают необходимыми для этого умственными способностями). В пещерах где они жили, находятся рога и кости со следами истирания, характер которых показывает употребление их для копки. Больше того, в европейских пещерах, населявшихся кроманьонцами и неандертальцами, найдены куски длинных костей, сломанные в точности в той манере, в какой ломал их Панбаниша.
Цитированные исследования показывают, что столкнувшись с задачей ступенчатого планирования действий, включающих использование орудия (более сложной чем те, с которыми имеют дело большие шимпанзе), бонобо неизменно оказываются не только способны её решить, но и вполне мотивированы решать.
Также показаны резкие различия между группой из зоопарка и из «Убежища…». Условия зоопарка не стимулируют проявление этих «гоминоидных» способностей, видимо из-за недоступности местообитаний типа природных. Вопреки этому, уникальные достижения фиксировались и у вуппертальских бонобо. Доминантная самка Ейя выбирала особенно длинные сучья и манипулировала ими. Отломав мелкие сучья и содрав кору, она обтачивала остриё в своего рода «дротик» и атаковала с ним Роффмана через решётку всякий раз, когда тот подходил.
 Шимпанзе (не эти, большие) пользуются такими дротиками на охоте на галаго, скрывающихся в дуплах, но не в социальном контексте. Роффман теоретизирует, почему на него стали нападать в зоопарке — его условия не позволяют человеку «войти своим» в группу бонобо, он всегда чужак, появляющийся «со стороны». Напротив, в превосходных условиях Убежища возможны персональные связи наблюдателя с обезьянами, в зоопарке же он лишь нарушает их приватность и отношения. Соответственно, один и тот же вид оказывается миролюбивым или же агрессивным-подозрительным в разных социальных контекстах.
Шимпанзе (не эти, большие) пользуются такими дротиками на охоте на галаго, скрывающихся в дуплах, но не в социальном контексте. Роффман теоретизирует, почему на него стали нападать в зоопарке — его условия не позволяют человеку «войти своим» в группу бонобо, он всегда чужак, появляющийся «со стороны». Напротив, в превосходных условиях Убежища возможны персональные связи наблюдателя с обезьянами, в зоопарке же он лишь нарушает их приватность и отношения. Соответственно, один и тот же вид оказывается миролюбивым или же агрессивным-подозрительным в разных социальных контекстах.
Поэтому можно заключить, что «телесно» (или «биологически») наш предок был больше всего похож на бонобо, включая этологию, структуру сообщества и пр., см. об этом в книге де Ваала «Истоки морали». Даже если эти особенности в «нашей ветви» появились независимо, а не унаследованы от общего предка, ввиду системного характера параллелизмов и наибольшей обоснованности идеи самодоместикации появление именно их (и сближающих именно с бонобо) закономерно, а не случайно, на них надо обращать внимание, если хотим вести речь о «биологии поведения» человека всерьёз.
А вот сближения с большим шимпанзе (развитая орудийная деятельность, агрессия) скорей конвергентны, ибо «сделаны» социальными факторами, вроде описанных выше. У больших шимпанзе это часть «биологии», у нас — следствие социальных факторов, двигавших антропогенез. Обоснование этого вывода для агрессии на основе работы Cieri, 2014 про краниофациальную феминизацию см. тут. Неслучайно Cieri следует идеям Беляева про следствия отбора на доместикацию (и даже цитирует его, что у западных авторов нечасто), а изменения рук «в человеческую сторону» под влиянием использования орудий у австралопитеков и недавно открытого Homo naledi появляются раньше, чем увеличивается мозг (и/или структурно приближается «к нам»). В то же время гипотеза синкинезии как модуса происхождения языка В. Рамачандрана позволяет связать с трудовой деятельностью и эту исключительно нашу особенность.
Примечания
1Отличающие также нас от обезьян, почему человека можно рассматривать как продукт педоморфоза. Однако у бонобо этот процесс зашёл глубже: в отличие от людей и больших шимпанзе, у них на всю жизнь сохраняется «детская» продукция гормонов щитовидной железы — трийодтиронина и тироксина (T3 и T4). У людей и больших шимпанзе она сокращается сразу после пубертата, у бонобо — медленно на протяжении всей последующей жизни, хотя и остаётся повышенной. При этом самцы бонобо (в отличие от больших шимпанзу долго сохраняющие связь со своими матерями, дружащие с самками, а не противостоящие им, почти не образующие коалиций) в течение жизни имеют большее высокий уровент Т3 и Т4, чем самки (Behringer et al., 2014).
2Сейчас это уже не «отдельное», а достаточно распространённое мнение: «The self-domestication hypothesis is therefore a potentially powerful tool for understanding the processes by which selection shapes both psychological and other seemingly unrelated traits, including those in humans (e.g. Leach et al. 2003; Dorus et al. 2004)
Рекомендуем прочесть
 «Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве
«Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве Об истории, культуре и современных проблемах коми. Часть 3.
Об истории, культуре и современных проблемах коми. Часть 3. Роль крымских татар в кампаниях Богдана Хмельницкого
Роль крымских татар в кампаниях Богдана Хмельницкого Зааплодировать до смерти
Зааплодировать до смерти «Глобальный подъём вспышек инфекционных болезней человека»
«Глобальный подъём вспышек инфекционных болезней человека» «Биолог-диалектик». Памяти Ричарда Левонтина.
«Биолог-диалектик». Памяти Ричарда Левонтина. Малоизвестная победа короля Фейсала над коммунизмом и СССР
Малоизвестная победа короля Фейсала над коммунизмом и СССР Гринвошинг, зарубежный и местный
Гринвошинг, зарубежный и местный
