Таблица I. 1 — цветок; 2 — лист; 3 — раскрывшаяся коробочка советского длинноволокнистого хлопчатника; 4 — раскрывшаяся коробочка советского хлопчатника; 5 — расправленная летучка и семена советского длинноволокнистого хлопчатника; 6 — расправленная летучка и семена советского хлопчатника; 7 — створка коробочки советского длинноволокнистого хлопчатника; 8 — створка коробочки советского хлопчатника; 9 — волокно советского длинноволокнистого хлопчатника; 10 — волокно советского хлопчатника; 11 — типы ветвления хлопчатника. Источник
В продолжение темы парфорсной эволюции
В.В. Суслов, Н.А. Колчанов
Резюме. Говоря об эволюции биоразнообразия прежде всего подразумевают эволюцию линейную и дивергентную. В работе рассмотрены случаи эволюции, связанной с перебором пространств возможностей. Такие пространства формируются на различных уровнях организации живого – от популяционно-ценотического и организменного до уровня биополимеров – белков и последовательностей ДНК и далее – до ключевых мономеров жизни. Ненулевая вероятность полного перебора не позволяет описать такую эволюцию как чисто конвергентную или чисто дивергентную – перед нами дивергентно-конвергентный цикл, но как и всякий процесс дарвиновской эволюции, такой цикл носит индетерминистский характер, иными словами, может быть разорван как на конвергентной, так и на дивергентной ветви при изменении вектора отбора.
Дилемма Холдейна ограничивает одновременную адаптивную эволюцию по многим генам в малых популяциях [Ратнер и др., 1985]. Между тем, нужда в эволюции велика именно в малых популяциях периферии ареала вида, живущих на пределе толерантности. Полногеномные проекты выявили высокую насыщенность генома полиморфизмами: внутри- и межвидовые сравнения геномов дают для разных видов оценки от 3 до 10 млн SNP на геном [Суслов и др., 2010]. Наконец, микрочиповые исследования выявили высокую консервативность наборов коэкспрессирующихся генов – регулогов – в эволюции. Так, сравнение данных из 3182 экспериментов по изучению экспрессии генов-ортологов таких отдаленных видов, как Homo sapiens (6591 ген), Drosophila melanogaster (5802 гена), Caenorhabditis elegans (5180 генов) и Saccharomyces cerevisiae (2434) выявило 22163 консервативных случая коэкспрессии, которые распределились по 11 регулогам [Stuart et al., 2003]. Консервативность последовательностей генов-гомологов, в регулогах разных видов, колеблется в среднем между 30%-60% [Babu et al., 2004].
В элементарном функциональном модуле «ген транскрипционного фактора (ТФ) + ген-мишень» в ходе эволюции возможны: дупликация гена ТФ, дупликация гена-мишени и оба эти события близко по времени. По первому пути у E. coli возникло ~22% регуляторных связей, по второму – ~10%, по третьему – ~6% (для дрожжей соотношения близки) [Teichmann, Babu, 2004]. Но хотя дупликация гена с 5′-фланкирующим районом удваивает не только белоккодирующую последовательностей, но и сайты связывания транскрипционных факторов (ССТФ), дупликации объясняют возникновение менее половины регуляторных связей в этих двух геномах. Как же остальные?
Здесь надо указать на традиционно недооцениваемую конвергенцию. Молекулярная база модуля «ген ТФ + ген-мишень» – соответствие конформаций активного сайта ТФ и ДНК ССТФ. Оба – порядка десятков мономеров (аминокислот или нуклеотидов), что делает вероятным неоднократное возникновение стереохимически сходных структур сайтов при многократном переборе ограниченного числа замен. Исследования in silico нашей лаборатории [Игнатьева и др., 2009, Ponomarenko et al., 2001] показали, что геном насыщен как функциональными, так и латентными ССТФ, чья функционализация требует считанных замен. Так полиморфизм –20 (C → A) в гене ангиотензиногена человека дает новый сайт связывания эстрадиолового рецептора, что повышает риск гипертонии беременных [Ponomarenko et al., 2003].
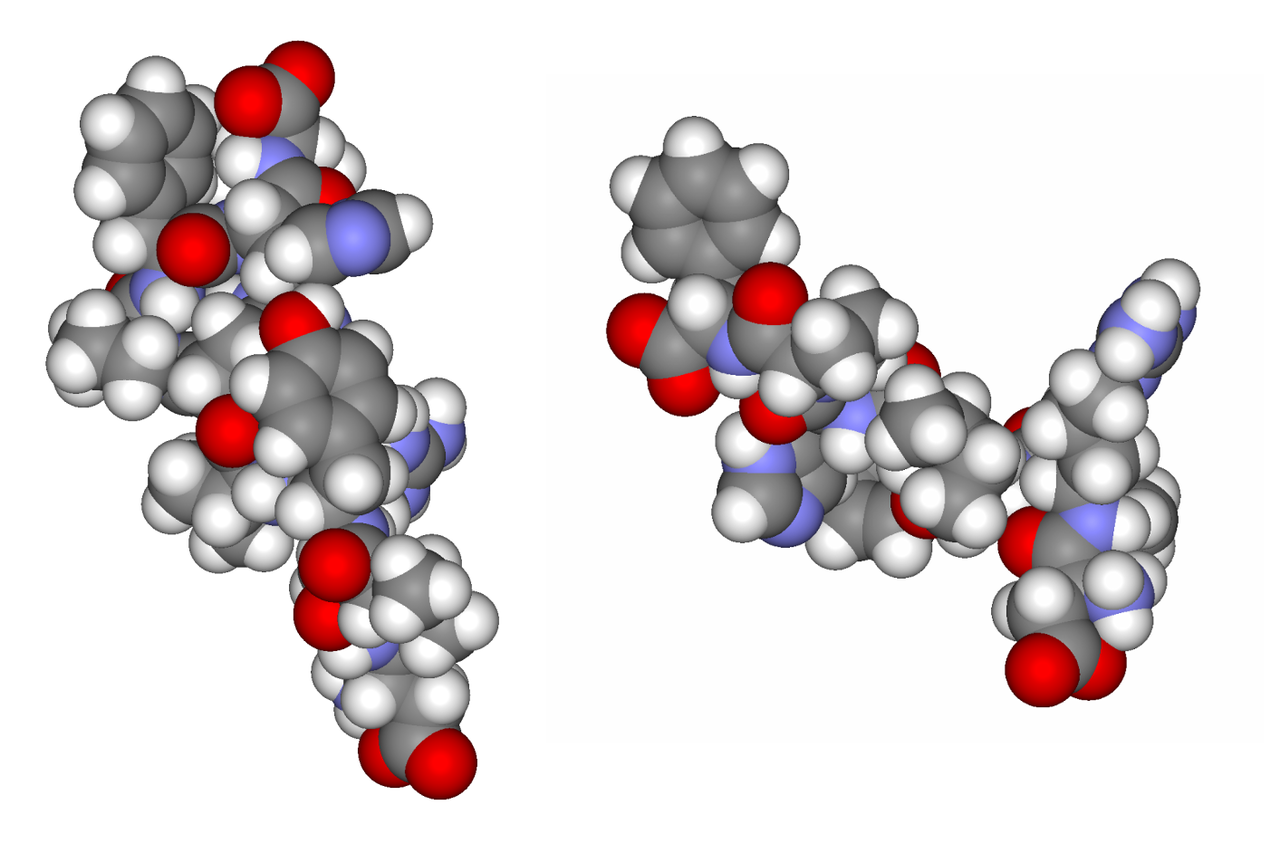
Ангиотензины I и II (образуются из ангиотензиногена действием ренина)
Разрушение ССТФ одиночными заменами также вероятно. Так мутация G → A в некодирующем районе гена триптофан-2,3-диоксигеназы, ведущая к синдрому Туретта, связана с разрушением ССТФ YY1 [Васильев и др., 2000]. Итак, если для протяженных последовательностей, (белоккодирующие районов генов), дупликации остаются единственным путем эволюции функций [Оно, 1973], то для малых последовательностей (ССТФ) важна также конвергенция. Аналогично белки несут латентные активные сайты, чья активация может быть вызвана как аминокислотной заменой [Ivanisenko et al., 2005], так и изменением в спектре наиболее вероятных конформаций глобулы (белковый промискуитет [Nobeli et al., 2009]) при помещении ее в нетипичные условия.
Таким образом, геном и протеом крайне изменчивы и пластичны. Отбор не может полностью очистить их от полиморфизмов (по крайней мере, за приемлемый срок). Как же сохранить единство фенотипа таксона при высоком уровне изменчивости (даже если значительная часть полиморфизмов нейтральна)?
В модуле «ген ТФ + ген-мишень» отбор оценивает функциональный параметр – одна и та же величина аффинности ТФ к ССТФ может быть получена за счет разных комбинаций мономеров. Таким образом, число комбинаций дает ограниченное пространство возможностей (ОПВ) для эволюции. Дивергенция последовательностей соответствует началу перебора ОПВ, конвергенция – исчерпанию ОПВ. Дивергентно-конвергентные циклы выявлены для эволюции ССТФ (ТАТА-бокс вируса ВИЧ-1 [Suslov et al., 2010]), белка (антигенные детерминанты вируса гриппа [Иванисенко и др., 2008], родопсины [Briscoe, 2002, Chang et al., 2002], рРНК [Алёшин и др., 1999]. Итак, ОПВ эволюции делает генотип вырожденным по отношению к фенотипу[1].
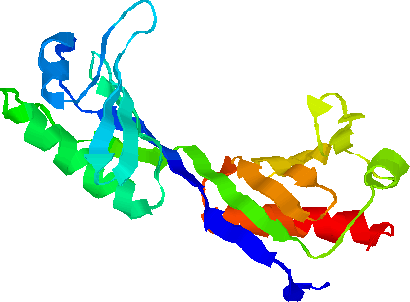
Пространственная структура ТАТА-связывающего белка
Дивергентно-конвергентная эволюция в ОПВ различной природы широко распространена. ОПВ может формировать структура популяции. В популяции из небольших полуизолятов возможно многократное восстановление характерных комбинаций полиморфизмов.
Так, полиморфизмы, характерные для промотора гена tb1 кукурузы, поодиночке распространены в различных популяциях теосинта и выступают как нейтральные или субнейтральные. Скрещивая представителей этих популяций, можно воспроизвести формы початка кукурузы, известные по археологическим находкам [Wang et al., 1999].
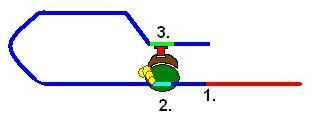
Упрощённая модель транскрипции эукариот. 1 — сайт начала транскрипции; 2 — ТАТА-бокс с РНК-полимеразой и транскрипционными факторами; 3 — энхансер, связанный с активирующим белком
Аналогично, цветы, характерные для цветной капусты и брокколи, возникают у кочанной капусты при объединении нонсенс-мутаций в 4ом и 5ом экзонах генов BoAP1-B и BoCAL, а у арабидопсиса – в их ортологах AP1 и СAL. Различные нонсенсы в данных экзонах существуют как полиморфизмы в популяциях крестоцветных [Lowman, Purugganan, 1999]. Выявлены ОПВ, формируемые взаимодействием аминокислот в пространственных структурах белков [Афонников, Колчанов, 2008]. ОПВ, формируемые «ломкими районами» на границах гетерохроматиновых блоков хромосом, обеспечивают дивергентное формирование кариотипов [Воронцов, 1988]. Пространство клеточной мембраны и внутриклеточное пространство также формируют свое ОПВ, где идет конвергентно-дивергентная эволюция адресации взаиморасположения функциональных блоков – молекул и супрамолекулярных комплексов (ионные каналы, белки-транспортеры, передатчики сигналов и т. д.), чьи «популяции» делают клетку в ходе дифференцировки анизотропной, позволяя выполнять ей те или иные физиологические функции [Уголев, 1988].
Наконец, ОПВ формирует генная сеть (ГС) онтогенеза. Наиболее простой случай – это конвергентное сходство онтогенезов у близких видов, диктующее параллельную эволюцию генов регуляции онтогенеза [Hughes, Friedman, 2003]. Гораздо интереснее случаи параллельного использования общих функциональных модулей в негомологичных органах или у неродственных таксонов. Действительно, конвергенции ССТФ позволяют присоединять к функциональному модулю негомологичные гены. Если это будет поддержано отбором – начнется усложнение модуля, а затем и экспансия его – многократное рекрутирование в разные локальные ГС организма. Общий пул регуляторных молекул позволяет экспансию, а ограничение накладывает повреждающий эффект плейотропных мутаций. Так, регуляторные связи между генами Polycomb-семейства и основными генами морфофункциональной спецификации – Hox для животных и MADS для растений – связаны с генезисом метамерных структур в обоих царствах [Meyerowitz, 2002].
Примером удачной экспансии служат вспомогательные циклины – паралоги основных циклинов[2] [Coudreuse, Nurse, 2010]. Общие регулоги выявлены в крыльях насекомых и конечностях позвоночных[3] [Шаталкин, 2003]. Сравнительная легкость появления или дефункционализации ССТФ разрешает блочно-модульную комбинаторную эволюцию. Так, исследование регулогов плаценты выявило мало ТФ с плацентоспецифической экспрессией. В дифференцирующейся плаценте коэкспрессируется множество генов, регулирующих рост и дифференцировку широкого круга тканей не только у млекопитающих, но и у земноводных, рыб и даже насекомых. Зрелая плацента обогащена таксон-специфичными (от отряда млекопитающих и ниже) семействами паралогов, ответственных за негативную регуляцию ростовых процессов. Таким образом, в ходе эволюции млекопитающих плацента была сформирована блочно-модульным способом из готовых ГС или их фрагментов независимо у сумчатых (бандикуты)[4] [Freyer et al., 2003] и плацентарных. У последних шла независимая эволюционная настройка плацент в ходе становления характерной для каждого отряда репродуктивной стратегии (многоплодие или малоплодие; рождение недоразвитых или зрелых детенышей и т. п.). В итоге плаценты у лемуров и обезьян дивергировали, а у лемуров и непарнокопытных – конвергировали. Конвергировали плаценты хищных, афротериев и неполнозубых, а в пределах парнокопытных произошла дивергенция [Knox, Baker, 2008, Hiden et al., 2007, Old, 2007, Cross et al., 2003].
Дивергентно-конвергентная эволюция в ОПВ молекулярно-генетической, хромосомной или популяционно-ценотической природы хорошо укладывается в рамки Закона гомологических рядов Н.И. Вавилова [1935]:
«1) виды и роды характеризуются сходными рядами наследственной изменчивости с такой правильностью, что зная ряд форм в пределах одного вида, можно предсказать параллельные формы других видов и родов;
2) семейства в общем характеризуются определенным циклом изменчивости, проходящим через все роды и виды».
Следует лишь отметить, что цикл изменчивости, как и всякий процесс дарвиновской эволюции, носит индетерминистский характер, а значит, начавшись, вовсе не обязательно должен закончиться. Разорвать «заколдованный круг» цикла изменчивости может как существенное изменение вектора движущего отбора, так и длительный период эволюции, близкой к нейтральной, разрушающий нефункционирующее пространство возможностей[5]. Кроме того, ввиду вымирания ряда таксонов, мы часто видим часть цикла как дивергентную эволюцию[6]. Наконец, методологически проще на текущий момент выявить дивергентную часть цикла.
Еще один важный универсальный механизмом, формирующий ОПВ – регуляторные контуры в ГС: отрицательной обратной связи, регулирующей концентрацию белка, безразличны причины отклонения от нормы: она равно компенсирует как физиологические отклонения, так и мутации. In silico моделирование эволюции популяции особей с отрицательной обратной связью, гомеостатирующей онтогенез, показало – в стабильных условиях шел отбор на усиление обратной связи. Побеждали особи с узкой нормой реакции, что соответствует теории стабилизирующего отбора И.И. Шмальгаузена [1968].
В нестабильных условиях среды обратная связь, наоборот, ослаблялась отбором до исчезновения, что вело к гиперманифестации – взрывообразному вскрытию замаскированной ранее изменчивости[7]. Это согласуется с работой Д.К. Беляева [1987] по доместикации лисиц, где высокий уровень изменчивости был побочным результатом ослабления нейрогуморальных регуляторных систем в ходе селекции на дружественное поведение к человеку.
Но для малой популяции, живущей на пределе толерантности, поиск адаптивных вариантов в широком спектре изменчивости, вскрытой гиперманифестацией, может недопустимо затянуться (ограничение дилеммы Холдейна). Надо уменьшить пространство перебора, ограничив его наиболее перспективными вариантами. Но как узнать, что будет перспективным?
Тем не менее, особи малых краевых популяций успешно эволюционируют, осваивая соседние биотопы и выходя в новые экологические ниши (часто – в отсутствие жесткой конкуренции внутри собственных ниш) [Фридман, Еремкин, 2008; Мальчевский, 1974, Osborn, 1934] (эффект Г. Осборна, впервые указавшего [1934] на возможную эволюционную роль нерегулярных находок единичных особей вдали от основного ареала вида). Между тем, если реализованная ниша всегда меньше фундаментальной, совпадая с ней лишь в отсутствие неблагоприятных факторов [Hutchinson, 1965], то фактически выход за пределы экониши невозможен без преадаптации. Но и в этом случае малая популяция вида-вселенца, по сравнению с многочисленным видом-аборигеном, отшлифованным эволюцией, будет в заведомо худших условиях[8], если преадаптация – не ароморфоз. Но аромофозы редки, требуют долгого генезиса [Северцов 2008], а инвазия за пределы экониш – явление, хоть и не массовое, но обычное. В.В.Жерихин [2003] объяснил частный случай: инвазия успешна, если вселенцы, нарушив ранние стадии сукцессии, перехватывают нишу у аборигенов, не вступая в прямую конкуренцию. Но и такие случаи редки[9].
Попав в неблагоприятные условия, организм автоматически активирует ГС стрессового ответа. Типичная стресс-реакция состоит из фаз тревоги, резистентности и истощения. Смертоносны для организма первая и последняя фазы. А в фазе резистентности организм способен оказать сопротивление множеству повреждающих факторов, а не только конкретному стрессору, вызвавшему стресс-ответ. Г. Селье [1972] назвал это явление перекрестной резистентностью. Таким образом, изменчивость в фазе перекрестной резистентности маскируется наиболее неспецифично, но кратковременно – в отличие от гомеостаза, стресс в долговременной перспективе не предусматривает возвращение организма к исходному состоянию (отсюда и фаза истощения – дистресс).
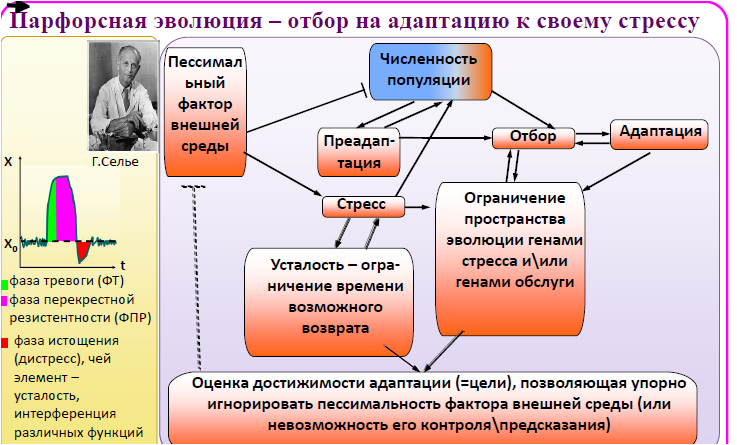
Логично если в крайне неблагоприятной среде (когда нет времени перебирать весь спектр полиморфизмов) или в среде высокоизменчивой[10] или неблагоприятной сразу по нескольким факторам (когда невозможен последовательный поиск вариантов, адаптивных для каждого фактора), для малой популяции наиболее адаптивными будут не мутации, обеспечивающие приспособление к конкретному фактору среды, а мутации, оптимизирующие функционирование ГС стрессового ответа (быстрое достижение фазы перекрестной резистентности) и купирующие дистресс в фазах тревоги и истощения.
Таким образом, в первых поколениях организм адаптируется не к среде, а к собственному стресс-ответу, тестируя мутации в ограниченном пространстве генов, экспрессирующихся в ходе стрессреакции. Лишь затем начинается приспособление к внешней среде – отбор тестирует прочий спектр изменчивости. Возможны также дупликации генов стресса: одна копия отвечает за перекрестную резистентность, другая – специализируется по конкретному стрессору, либо рекрутируются в ГС онтогенеза. В итоге формируется воронкообразный тренд эволюции: в первых поколениях под отбор попадают гены стресса. С ростом числа поколений набор генов, втянутый в орбиту отбора, расширяется, делая каждую конкретную траекторию эволюции все менее предсказуемой.
Мы назвали такую эволюцию парфорсной (от франц. par force – буквально «через силу»). Фактически на первой стадии парфорсной эволюции, в результате функционирования генных сетей стресса, в огромном спектре генотипической изменчивости выделяется ОПВ для эволюции, связанной с генами, чья адаптивность уже многократно опробована в филогенезе таксона. На второй стадии это ОПВ постепенно расширяется.
Эволюция хлопкового волоконца идеально укладывается в двустадийную схему парфорсной эволюции. Волоконца – удлиненные клетки, формирующие микроклимат в коробочке. У примитивных хлопков с геномом F в формировании волоконца основную роль играют гены стресса. Генерируемые ими перекись водорода и активные формы кислорода размягчают клеточную стенку, позволяя клетке удлиняться. Но формирование волоконца быстро завершается, и оно оказывается коротким – клетка попадает в фазу истощения, так как активный кислород повреждает жизненно важные органеллы [Hovav et al., 2008]. У длинноволоконных азиатских хлопков А, под адаптивную эволюцию попали гены, очищающие клетку от активных форм кислорода. У тетраплоидных американских хлопков АD произошло адаптивное перераспределение регуляции формирования волоконца между генами контроля окислительного метаболизма (геном А) и генами нормального онтогенеза (геном D) [Xu et al., 2010, Chaudhary et al., 2009].
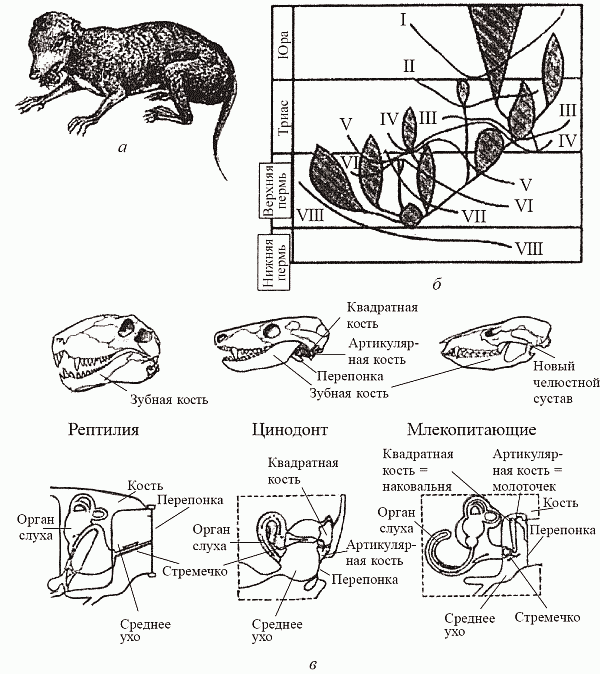
Происхождение млекопитающих: «маммализация териодонтов». (а) — Megazostrodon, древнейшее из известных млекопитающих из верхнего триаса; (б) — схема филогенеза по Татаринову (1976), демонстрирующая очередность приобретения маммальных признаков в различных филогенетических линиях тероморфов: I — звукопроводящий аппарат из трех слуховых косточек, II — вторичное челюстное сочленение между зубной и чешуйчатой костями, III — зачаточная барабанная перепонка в вырезке угловой кости. IV — мягкие, снабженые собственной мускулатурой губы, V — сенсорная зона на верхней губе (протовибриссы), VI — расширенные большие полушария головного мозга, VII — трехбугорчатые заклыковые зубы, VIII — верхние обонятельные раковины; (в) — эволюция челюстных структур и среднего уха (иллюстрирующая предыдущую схему). Источник
Формирование волосяного покрова – элемент ароморфной маммализации терапсид. Однако и в ходе заживления ран запускается формирование волосяных фолликулов (большинство из них резорбируется еще до образования рубца) [Ito et al., 2007]. Таким образом, ароморфный комплекс генов работает и при стрессе. Устойчивость к облучению (редкий в естественной среде стрессор) связана, в частности, с генами оксидативного стресса [Azzam, 2010, Vaquero, Reinberg, 2009, Storz et al. 2000] и генами засухоустойчивости [Лось, 2007] (обычные стрессоры). Генерализованный ответ на такие стрессоры, как гипер- и гипоосмос, низкие и высокие температуры [Кржечковская и др., 2004, Chao et al. 2009, Liu et al. 2009, Vaquero, Reinberg, 2009, Macario et al. 1999], гипоксия и ксенобиотики [Kawaura et al. 2009, Semenza, 2006] включает общую фракцию генов, причем одни и те же гены могут выполнять разные функции в одной стресс-реакции [Chao et al. 2009, Liu et al. 2009, Vaquero, Reinberg, 2009].
Важно, что в фазе перекрестной резистентности организм способен кратковременно покидать пределы экологической ниши вида, чем и объясняется вышеотмеченный эффект Осборна. Например, освоение урболандшафтов [Фридман, Еремкин, 2008] начинается не с развития частных адаптаций к каким-либо ресурсам, а с толерантности к генерализованному стрессору беспокойства. Характерно, что такая адаптация идет очень быстро (за несколько поколений), но далеко не в каждой популяции вида. Таким образом, схему экологической ниши следует дополнить стресс-периферией, выходящей за рамки фундаментальной экониши, но доступной лишь субпопуляции стресс-устойчивых особей. Стереотипность стресс-реакции обеспечивает параллелизм парфорсной эволюции не только в отдаленных популяциях, но и в репродуктивно изолированных таксонах.
Так Ю.О.Раушенбах [1981] показал, что отбор на адаптацию к горным условиям (гипоксия) в ходе отгонов равнинных пород овец на горные пастбища в первых поколениях приводит к росту кислородного запроса (идет отбор на сохранение уровня кислородного снабжения тканей через сеть капилляров), а в последующих – к снижению кислородного запроса (идет отбор на экономное использование доступного кислорода). Аналогичные адаптации выявлены у кратко- и долгоныряющих морских млекопитающих [Петров, 2003, Галанцев, 1988].

Источник (ДХ — дилемма Холдейна)
Наконец, следует ожидать, что вместе с отбором на оптимизацию стресс-реакции будет стабилизирована норма реакции для модификаций, возникающих в результате работы ГС стресса. Теория генокопирования модификаций была разработана в трудах И.И. Шмальгаузена [1968], Е.И. Лукина [1935] и В.С. Кирпичникова [1935]. Согласно ей отбираются мутации, увеличивающие вероятность формирования адаптивных модификаций. Однако, тестирование модификаций на адаптивность, а затем отбор мутаций-генокопий, согласно дилемме Холдейна, затруднительны для малой популяции. Парфорсная эволюция обходит проблему: во-первых, адаптивность стресс-ответа многократно опробована в филогенезе таксона (строго говоря, модификация может быть нейтральной или умеренно вредной – достаточно, чтобы она была физиологически связана со стресс-реакцией; адаптивность обеспечивается не модификацией, а стресс-реакцией в целом); во-вторых, перекрестная резистентность может быть адаптивна к нескольким факторам.
Таким образом, эволюция в ОПВ различной природы широко распространена на различных уровнях организации живого – от популяционно-ценотического и организменного до уровня биополимеров – белков и последовательностей ДНК. Спускаясь еще ниже – на уровень мономеров, следует отметить, что, многие из них (пурины, пиримидины, порфирины и т. д.) принадлежат к резонансным системам, содержащим чередующиеся двойные и одинарные связи. Распределение электронов в таких системах вырождено относительно структуры молекулы [Крепс, 1988]. Таким образом, уже на этапе химической эволюции жизнь (по крайней мере, известная нам) связана с дивергентно-конвергентными процессами в пространствах возможностей.
Литература
1.Алёшин В.В., Владыченская Н.С., Кедрова О.С. и др. 1999. Вторичная структура шпильки 17 низшего многоклеточного животного Rhopalura ophiocomae (Mesozoa: Orthonectida) как пример «прерываемого равновесия» в эволюции 18S рибосомной РНК // Мол. Биол. Т. 33. С. 319 – 329.
- Афанасьева Г.А. 1984. Брахиоподы отряда Chonetida: диссертация д.б.н. М.: ПИН РАН. 290 с.
- Афонников Д.А., Колчанов Н.А. 2008. Координированные замены аминокислотных остатков в белках // Системная компьютерная биология. Новосибирск: СО РАН. C. 591 – 602.
- Беляев Д.К. 1987. Дестабилизирующий отбор // Развитие эволюционной теории в СССР (1917–1970 годы). Л.: Наука. С. 266 – 277.
- Вавилов Н.И. 1935. Закон гомологических рядов в наследственной изменчивости. М.-Л.: Сельхозгиз. 56 с.
- Василевич В.И. 2009. Естественный отбор в растительных сообществах // Чарльз Дарвин и современная наука. Спб.: СпбНЦ. С. 29 – 31.
- Васильев Г.В., Меркулов В.М., Кобзев В.Ф. и др. 2000. Точковые мутации в районе 663-666 п.н. интрона 6 гена триптофаноксигеназы, связанные с рядом психических расстройств, разрушают сайт связывания фактора транскрипции YY1 // Мол. Биол. Т. 34. С. 214 – 222.
- Воронцов Н.Н. 1988. Постепенное или внезапное видообразование: “или – или” или “и-и” // Дарвинизм: история и современность. Л.: Наука. С. 87- 103.
- Галанцев В.П. 1988. Адаптации сердечно-сосудистой системы вторичноводных амниот. Л.: ЛГУ. 220 с.
- Гребельный С.Д. 2008. Клонирование в природе. Спб.: ЗИН РАН. 287 с.
- Жерихин В.В. 2003. Основные закономерности филоценогенетических процессов (на примере неморских сообществ мезозоя и кайнозоя) // Избранные труды. М.: КМК. С. 405–439.
- Жерихин В.В. 1986. Гипотрофные озера – вымерший тип мезозойских экосистем // История древних озер в дочетвертичное время, в голоцене и формирование полезных ископаемых в озерах. Тез. докл. VII симпозиума по истории озер. Т. 1. Л.: ЗИН АН СССР. С. 55– 61.
- Иванисенко В.А., Деменков П.С., Фомин Э.С. и др. 2008. Распознавание функциональных сайтов в пространственных структурах белков // Системная компьютерная биология. Новосибирск: СО РАН. C. 231 – 245.
- Игнатьева Е.В., Меркулова Т.И., Ощепков Д.Ю. и др. 2009. Выявление новых сайтов связывания транскрипционных факторов SREBP в промоторных районах генов позвоночных на основе комбинации биоинформатического и экспериментального подходов // Вестник ВОГиС. Т. 13, №1. С.37–45.
- Кирпичников В.С. 1935. Роль ненаследственной изменчивости в процессе естественного отбора (гипотеза о косвенном отборе) // Биологический журнал. Т.4. №5. С.775–800.
- Крепс Е.М. 1988. О некоторых путях исследования биохимической эволюции // Дарвинизм: история и современность. Л.: Наука. С. 195 – 202.
- Кржечковская В.В., Кубатиев А.А., Наумов Ю.И. 2004. Мембранносвязанный цитохром b5 и метаболизм липидов (реакции, не связанные с участием цитохрома Р-450) // Мембраны, Серия “Критические технологии”, ВИНИТИ. №2(22). С. 9 – 21.
- Лось Д.А. 2007. Восприятие стрессовых сигналов биологическими мембранами // Проблемы регуляции в биологических системах. Биофизические аспекты. М. – Ижевск НИЦ «Регулярная и хаотическая динамика», Институт компьютерных исследований. С. 329 – 360.
- Лукiн Е.И. 1935. Проблема географiчноi мiнливостi организмiв // Ученi записки Харкiвського унiверситету. №2/3. С. 15 – 22.
- Мальчевский А.С. 1974. Отношение животных к территории как фактор эволюции (на примере птиц) // Вестник ЛГУ. Биология. №3. С. 5 – 15.
- Оно С. 1973. Генетические механизмы прогрессивной эволюции. М.: Мир. 222 с.
- Петров Е.А. 2003. Байкальская нерпа: эколого-эволюционные аспекты: Автореф. дис. докт. биол. наук. Улан-Удэ: Изд-во Бурят. ун-та. 38 с.
- Ратнер В.А. Жарких А.А., Колчанов Н.А. и др. 1985. Проблемы теории молекулярной эволюции. Новосибирск: Наука. 260 с.
- Ратнер В.А. 1964. Принцип связности серий и определение порядка оснований в кодонах // Биофизика. Т. 9, №5. С. 621 – 625.
- Раушенбах Ю.О. 1981. Закономерности экогенеза домашних животных // Генетика. Т. 17, №9. С. 1663 – 1677.
- Розанов А.Ю. 1973. Закономерности морфологической эволюции археоциат и вопросы ярусного расчленения нижнего кембрия. М.: Наука. 164 с.
- Северцов А.С. 2008. Причины и условия формирования ароморфной организации // Ж.О.Б. Т. 69, №2. С. 94 – 101.
- Селье Г. 1972. На уровне целого организма. М.: Наука. 122 с.
- Суслов В.В., Пономаренко П.М., Пономаренко М.П. и др. 2010. Полиморфизмы ТАТА-боксов генов хозяйственно важных и лабораторных животных и растений, ассоциированные с их селекционно-ценными признаками // Генетика. Т. 46, №4. С. 448 – 457.
- Уголев А.М. 1985. Эволюция пищеварения и принципы эволюции функций. Элементы современного функционализма. Л.: Наука. 544 с.
- Фридман В.С., Еремкин Г.С. 2008. Урбанизация «диких» видов птиц в контексте эволюции урболандшафта. М.: МГУ. 139 с.
- Шаталкин А.И. Регуляторные гены в развитии и проблема морфотипа в систематике насекомых. Чтения памяти Н.А. Холодковского. Вып. 56(2). СПб. : ЗИН РАН. 109 с.
- Шмальгаузен И.И. 1968. Факторы эволюции (теория стабилизирующего отбора). М.: Наука. 452 с.
- Azzam E. 2010. The impact of bystander effects and adaptive responses in the biological responses to low dose/low fluence ionizing radiation: the modulating effect of linear energy transfer // Modern problems of genetics, radiobiology, radioecology and evolution. Dubna. P. 53.
- Babu M.M., Luscombe N.M., Aravind L. et al. 2004. Structure and evolution of transcriptional regulatory networks // Curr. Opin. Struct. Biol. V. 14, №3. P. 283-291.
- Briscoe A.D. 2002. Homology modeling suggests a functional role for parallel amino acid substitutions between bee and butterfly red- and greensensitive opsins // Mol. Biol. Evol. V. 19, №6. P. 983 – 986.
- Chang B.S., Jönsson K., Kazmi M.A. et al. 2002. Recreating a functional ancestral archosaur visual pigment // Mol. Biol. Evol. V. 19, №9. P. 1483 –1489.
- Chao H.F., Yen Y.F., Ku M.S. 2009. Characterization of a salt-induced DhAHP, a gene coding for alkyl hydroperoxide reductase, from the extremely halophilic yeast Debaryomyces hansenii // BMC Microbiol. V. 9:182. Epub.
- Chaudhary B., Hovav R., Flagel L., et al. 2009. Parallel expression evolution of oxidative stress-related genes in fiber from wild and domesticated diploid and polyploid cotton (Gossypium) // BMC Genomics. V. 10:378. Epub.
- Coudreuse D., Nurse P. 2010. Driving the cell cycle with a minimal CDK control network // Nature. V. 468, №7327. P. 1074 – 1079.
- Cross J.C., Baczyk D., Dobric N. et al. 2003. Genes, development and evolution of the placenta // Placenta. V. 24, №2-3. P. 123 – 130.
- Dunbrack R.L., Ramsay M.A. 1989. The evolution of viviparity in amniote vertebrates: egg retention versus egg size reduction // Amer. Nat. V. 133, №1. P. 138 – 148.
- Freyer C., Zeller U., Renfree M.B. 2003. The marsupial placenta: a phylogenetic analysis // J. Exp. Zoolog. A. Comp. Exp. Biol. V. 299, №1. Р. 59 – 77.
- Gurovich Ya. 2005. Bio-evolutionary aspects of Mesozoic Mammals. Buenos Aires: Univ. de Buenos Aires. 546 p.
- Hutchinson G.E. 1965. The niche: an abstractly inhabited hypervolume // The ecological theatre and the evolutionary play. New Haven: Yale Univ. Press. P. 26-78.
- Hayssen V., Lacy R.C., Parker P.J. 1985. Metatherian reproduction: transitional or transcending? // Amer. Nat. V. 126, №5. P. 617 – 632.
- Hiden U., Bilban M., Knofler M., et al., 2007. Kisspeptins and the placenta: regulation of trophoblast invasion // Rev. Endocr. Metab. Disord., V. 8. №1. P. 31-39.
- Hughes A.L., Friedman R. 2003. Parallel evolution by gene duplication in the genomes of two unicellular fungi // Genome Res. V. 13, №6A. P. 1259 – 1264.
- Hovav R., Udall J.A., Chaudhary B., et al. 2008. The evolution of spinnable cotton fiber entailed prolonged development and a novel metabolism // PLoS Genet. V. 4, №2:e25. Epub.
- Ito M., Yang Z., Andl T., et al. 2007. Wnt-dependent de novo hair follicle regeneration in adult mouse skin after wounding // Nature. V. 447, №7142. P. 316 – 320.
- Ivanisenko V.A., Pintus S.S., Grigorovich D.A., et al. 2005. PDBSite: a database of the 3D structure of protein functional sites // NAR. V. 33, Database issue. Р. D183 – D187.
- Кawaura K., Mochida K., Enju A. et al. 2009. Assesment of adaptive evolution between wheat and rice as deduced from full-length common wheat cDNA sequence data and expression patterns // BMC Genomics. V. 10:271. Epub.
- Knox K., Baker J.C. 2008. Genomic evolution of the placenta using co-option and duplication and divergence // Genome Res. V. 18. №5. P. 695 – 705.
- Lowman A.C., Purugganan M.D. 1999. Duplication of the Brassica oleracea APETALA1 floral homeotic gene and the evolution of domesticated cauliflower // J. Hered. V. 90, №5. P. 514 – 520.
- Liu Q., Wang H., Zhang Z., et al. 2009. Divergence in function and expression of the NOD26-like intrinsic proteins in plants // BMC Genomics. V. 10:313. Epub.
- Macario A.J., Lange M., Ahring B.K., et al. 1999. Stress genes and proteins in the archaea // Microbiol. Mol. Biol. Rev. V. 63, N 4. P. 923 – 967.
- Meyen S.V. 1971. Parallelism and its significance for the systematics of fossil plants // Geophytology. V. 1, №1. P. 34 – 47
- Meyerowitz E.M. 2002. Plants compared to animals: the broadest comparative study of development // Science. V. 295, №5559. P. 1482 – 1485.
- Nobeli I., Favia A.D., Thornton J.M. 2009. Protein promiscuity and its implications for biotechnology // Nat. Biotechnol. V 27, №2. P. 157 – 167.
- Old L.J. 2007. Cancer is a somatic cell pregnancy // Cancer Immunity. V. 7. P. 19-21.
- Osborn H.F. 1934. Aristogenesis, the Creative Principle in the Origin of Species // Amer. Nat. V. 68, №716. P. 193 – 235.
- Ponomarenko J.V., Merkulova T.I., Orlova G.V., et al. 2003. rSNP_Guide, a database system for analysis of transcription factor binding to DNA with variations: application to genome annotation // NAR. V. 31, №1. P. 118 – 121.
- Ponomarenko J.V., Furman D.P., Frolov A.S., et al. 2001. ACTIVITY: a database on DNA/RNA sites activity adapted to apply sequence-activity relationships from one system to another // NAR. V. 29. №1. P. 284 – 287.
- Semenza G.L. 2006. Regulation of physiological responses to continuous and intermittent hypoxia by hypoxia-inducible factor 1 // Exp. Physiol. V. 91, №5. P. 803 – 880.
- Storz H., Hengge-Aronis G., Yura T., et al. 2000. Bacterial stress responses. Washington: ASM Press. 485 p.
- Stuart J.M., Segal E., Koller D., et al. 2003. A gene-coexpression network for global discovery of conserved genetic modules // Science. V. 302, №5643. P. 249 – 255.
- Suslov V.V., Ponomarenko P.M., Efimov V.M., et al. 2010. SNPs in the HIV-1 TATA box and the AIDS pandemic // J. Bioinform. Comput. Biol. V. 8, №3. P. 607 – 25.
- Teichmann S.A, Babu M.M. 2004. Gene regulatory network growth by duplication // Nat. Genet. V. 36, №5. P. 492 – 496.
- Vaquero A., Reinberg D. 2009. Calorie restriction and the exercise of chromatin // Genes Dev. V. 23, №16. P. 1849 – 1869.
- Wang R.L., Stec A., Hey J., et al. 1999. The limits of selection during maize domestication // Nature. V. 398, N 6724. P. 236 – 239.
- Xu Z., Yu J.Z., Cho J., et al. 2010. Polyploidization altered gene functions in cotton (Gossypium spp.) // PLoS One. V. 5, №12:e14351. Epub.
Примечания
[1]Вырожденность генетического кода – частный случай: ОПВ ограничено третьей (реже – второй и третьей) позицией кодона. Дивергентно-конвергентные от-ношения в сериях кодонов были предсказаны В.А.Ратнером [1964] (хотя он и не употреблял этого термина).
[2]Так, у млекопитающих вспомогательный циклин Н/CDK7 активирует киназы – партнеры основных циклинов (СDC2/CDK1, CDK2, CDK4, CDK6), ускоряя прохождение клеточного цикла; циклин С вовлечен в регу ляцию почти всех РНК-роl II – зависимых генов; циклин М связан с регуляцией пре-мРНК сплайсинга.
[3]Экспансия модуля может вести к его различному использованию у личинок и имаго: ложноножки у гусениц по составу коэкспрессирующихся ТФ гомологичны грудным ногам, а у личинок пилильщиков – мандибулам [Шаталкин, 2003].
[4] Если рассматривать эпителиохориальную плаценту бандикутов как анцестральный признак, то плаценту мог иметь уже предок сумчатых. Ряд специализированных черт размножения Metatheria не позволяет считать его более близким к предковому, нежели Eutheria [Hayssen et al., 1985]. Но тогда широкое распространение плацентации, в свою очередь, говорит в пользу параллельного формирования живорождения у ряда вымерших высших терапсид [Gurovich, 2005], конвергентно достигших маммального уровня по целому комплексу морфологических показателей, провоцирующих живорождение [Dunbrack, Ramsay, 1989].
[5]Во всяком случае, наиболее подтвержденные циклы дивергентно-конвергентной эволюции мы имеем либо из вирусологии [Иванисенко и др., 2008, Suslov et al., 2010], где эволюция идет быстро и все потомки налицо, либо из палеонтологии прикрепленных пассивно питающихся организмов (археоциаты [Розанов, 1973], брахиоподы [Афанасьева 1984], растения [Meyen, 1971]), где возможности движущего отбора ограничены.
[6]Именно такой до недавнего времени представлялась эволюция плацентарного размножения. Эволюцию кариотипов в большинстве источников считают дивергентной, несмотря на работы Н.Н.Воронцова [1988].
[7]Подробнее см. тезис Гунбин К.В., Афонников Д.А., Турнаев И.И., Суслов В.В., Матушкин Ю.Г., Колчанов Н.А. «Компьютерный анализ эволюции генных сетей морфогенеза».
[8] Тот факт, что инвазию ведут единицы (за исключением «волн жизни») в перспективе делает инвайдера уязвимыми: генофонд его популяции обеднен по сравнению с популяцией вида-аборигена [Гребельный, 2008].
[9]Тем более, что в качестве основных примеров В.В. Жерихин [2003] опять таки рассматривал классические ароморфозы – формирование цветка и энтомофилию цветковых растений. Пожалуй, только формирование обильного опада [Жерихин, 1986] у цветковых не относится к ароморфозу.
[10] Т.е. скорость изменения факторов среды на порядки превосходит скорость смены поколений: типичная ситуация для сидячих организмов (особенно для долгоживущих растений [Василевич, 2009]).
Развитие жизни в процессе абиотических изменений на земле. Иркутск/Листвянка, Ин-т географии им. В.Б.Сочавы СО РАН, 2011. С. 193-207