

Одно из немногих исследований спонтанной кооперации у шимпанзе, для чего опыты с обменом жетонов пришлось сильно упростить. Источник
В продолжение темы
(обзор исследований последних 10 лет)
Федорович Е.Ю.
1. Изучение социо-когнитивных способностей животных – немного истории
Содержание
- 1 1. Изучение социо-когнитивных способностей животных – немного истории
- 2 2. Экспериментальные парадигмы исследований социо-когнитивных способностей человекообразных обезьян – эволюционных предпосылок «нравственных основ» поведения людей
- 3 3. Сотрудничают или решают собственные проблемы при помощи другого?
- 4 В заключение
Вопросы изучения «социо-когнитивных способностей животных» занимают существенный объём в современных научных журналах, освещающих вопросы сравнительной психологии. Появилось целое направление научных исследований, названное «Social Cognition» (Shettleworth, 2010). Экспериментальные парадигмы, используемые при изучении социально-когнитивных способностей животных, могут быть прослежены от работ N. Humphrey (1976) и D. Premack и G. Woodruff (1978).
В своей работе «Социальная функция интеллекта» N. Humphrey высказал идею, что главными движущими силами в эволюции человеческих когнитивных способностей были те ситуации и задачи, с которыми сталкивались животные, проживающие в сложно структурированных, долго существующих группах, при взаимодействии с другими индивидами.
Высокоразвитые когнитивные способности позволяли им предсказывать и использовать действия других членов группы (Byrne, Whiten, 1988). D. Premack и G. Woodruff (1978), обсуждая подобные возможности, ввели понятие «theory of mind» [теория психического [состояния у другого субъекта]] , под которым понимали способность индивидов принимать решения, основываясь на представлениях о психических или ментальных состояниях (убеждениях, желаниях, намерениях и т.п.) других особей. Наличие подобной способности давало бы животным возможность не только предсказывать действия других членов своей группы, исходя из представлений о том, как другой индивид представляет ту же самую ситуацию (to take the perspective of others), но и намеренно обманывать их, добиваясь собственных целей. Позднее эта идея была развита R. Byrne и А. Whiten (1988) в работе «Machiavellian intelligence» [маккиавеллиевский интеллект — умение пользоваться побуждениями и поступками другого для собственных целей, в первую очередь для приматов — в конкуренции за статус в группе и поддержку/»уважение» других особей, без которых его не заполучить].
Следует заметить, что на сегодняшний день в целом признаётся, что животные обладают лишь отдельными элементами подобной способности, в полной же мере «theory of mind» присуща лишь людям; см., например, Томаселло, 2011; Hare, 2011; Moll, Tomasello, 2007; Shettleworth, 2010; Tomasello, Vaish A., 2013).
В 2000-х годах исследования социо-когнитивных способностей у животных в значительной степени сконцентрировались на проблемах «кооперации» или «сотрудничества», в первую очередь человекообразных обезьян (Moll, Tomasello, 2007). Отчасти, обсуждение возможностей и механизмов координации действий и кооперации у приматов была спровоцирована описанием «коллективных охот» шимпанзе на колобусов, проведённых в природе (Boesch, Boesch, 1989). Отчасти, интерес к отличию «подлинного сотрудничества» у людей от совместных действий животных был поднят представленной в конце 1990-х гг. H. Moll и М. Tomasello гипотезой «Выготскианского социального интеллекта» (Moll, Tomasello, 2007), и её последующей интенсивной экспериментальной разработкой.
Эта гипотеза определяла те характеристики поведения и социо-когнитивные способности, которые принципиально отделяют «сотрудничество» человека от «кооперации» или «совместных действий» других животных, а также приводит к появлению у людей культуры, новых форм социального научения, коммуникации и воспитания потомков (Томаселло, 2011). В последние годы интерес к психологическим механизмам, позволяющим людям, в отличие от их ближайших биологических родственников – антропоидов, вступать в «подлинное сотрудничество», привел к интенсивному исследованию у последних (прежде всего на примере шимпанзе, и в гораздо меньшей степени – бонобо, орангутанов) эволюционных корней таких способностей как «чувства справедливости» (sense of fairness), «чувства равенства» (equality), «альтруизма», «оказания помощи» (helping), «эмпатии» (empathy) и других, тому подобных; общим для них всех является то, что именно эти качества всегда считались отличающими людей от всех других представителей Царства Животных. Наиболее известными научными коллективами, представляющими свои экспериментальные исследования в этой области являются группа Michael Tomasello (Josep Call, Alicia P. Melis, Anke F. Bullinger и др.) c активно сотрудничающим с ними Felix Warneken, Brian Hare, группа France de Waal, группа Elisabetta Visalberghi.
2. Экспериментальные парадигмы исследований социо-когнитивных способностей человекообразных обезьян – эволюционных предпосылок «нравственных основ» поведения людей
Несмотря на то, что у антропоидов изучается достаточно сложный ряд психических «способностей», которые мы бы отнесли в разряд морально-нравственных, этических, подавляющее большинство экспериментальных исследований строятся на основе двух практических задач, которые должные
решить антропоиды: подтягивание досок/платформ с выложенным на них пищевым вознаграждением и обмен жетонов на еду. Кроме того, ограничивающим возможные выводы положением (и с биологической, и с психологической позиций) является то, что, как правило, поиск «нравственных эволюционных основ» строится на пищевой мотивации: обезьяны добывают пищу в различного рода ситуациях, отвергают и делятся ею. Все экспериментальные схемы можно объединить в следующие группы:
2.1 Подтягивание платформ
Перед индивидом (или двумя и более индивидами), вне их досягаемости, помещается платформа/длинная и широкая доска, которую обезьяны должны подтянуть за концы верѐвки, свободно пропущенной через петли на концах этого сооружения. Если потянуть за один конец – верѐвка свободно высвобождается из петель и вознаграждение остаѐтся недоступным.
Соответственно, обезьяны (все индивиды, участвующие в исследованиях, предварительно получают соответствующий опыт) могут достать приманку только потянув за оба конца верѐвки одновременно. На платформе/доске выкладывают пищевое вознаграждение. В зависимости от целей экспериментаторов пищевое вознаграждение может находиться в одном месте платформы или быть распределено в нескольких её местах; оно может быть «легко монополизируемым», т.е. схваченным одним индивидом сразу, например, большой кусок фрукта, либо же распределено дисперсно, например, в виде множества мелко нарезанных кусочков фруктов.
Кроме того, вознаграждение может быть «равным» (одинаковое количество пищи на противоположных концах доски) или «неравным» (например, один кусочек банана на одной стороне и несколько кусочков банана – на другой стороне доски).
Соответственно, регулируя длину платформы и длину концов верёвки, можно создавать условия, в которых одна обезьяна не может подтянуть платформу самостоятельно, так как не может дотянуться до обоих концов одновременно. Тогда для решения этой задачи требуется усилия двух животных, действующих скоординированно.

Данная задача впервые была использована Карпентером в 1930-хх годах, а своё «второе рождение» она пережила в 2000-х годах. При помощи данной методики решались следующие вопросы:
- понимают ли шимпанзе необходимость партнёра для решения задачи:
— будут ли они выпускать другого индивида из его клетки, если не могут сами подтянуть платформу или выберут соло-платформу, если партнёр отсутствует (напр., Melis et al., 2006а, b);
— какого партнёра они будут «рекрутировать» в случае необходимости. Например, в ситуации, когда нельзя в одиночку подтянуть платформу, у шимпанзе есть возможность выпустить из соседних клеток одного из двух возможных партнёров, которые ранее, находясь в такой же ситуации, либо выпускали, либо не выпускали его самого (Melis et al., 2006а);
— есть ли у антропоидов т.н. просоциальное поведение, т.е. такое поведение, которое выполняется одним индивидом, чтобы облегчить нужды других индивидов или улучшить их благополучие (напр., Bräuer et al., 2009; Amici et al., 2014). Это т.н. Prosocial Choice Test. Например, в задаче с подтягиванием платформ, субъектам на выбор предоставляются две возможные конструкции: сцепленные платформы, подтягивание одной из которых приводит к приближению к клетке
соседа прикреплённой к ней второй (Silk et al., 2005).Субъекты (действующие индивиды) всегда получают одно и то же количество пищи, вне зависимости от выбранного варианта конструкции, однако при одном варианте платформы пищу получает только действующий индивид, а при другом – также и партнёр. Выбирая последний вариант, субъект может, таким образом, принести пользу партнѐру, при этом не совершая дополнительных затрат.
— зависит ли количество полученной пищи от того, как она была добыта (при совместном подтягивании платформы или в ситуации, когда один тянет, а другой смотрит) (напр., Melis et al., 2011).
Особый класс экспериментов с подтягиванием платформ представляют сконструированные для животных варианты так называемых «экономических игр» (economic games) – схемы исследований, пришедшие из изучения чисто человеческих сфер деятельности (экономики, социологии, политологии и пр.)
и применяющиеся для объяснения и предсказания поведения «экономических агентов» (в нашем случае – отдельных индивидов человекообразных обезьян), ведущих борьбу за реализацию своих интересов в ситуациях, требующих от них «распределения ресурсов». В основе «экономических игр» лежит «теория игр» – раздел математической экономики, изучающий оптимальные стратегии разрешения конфликта интересов.
Примером экспериментально оформленной для исследования
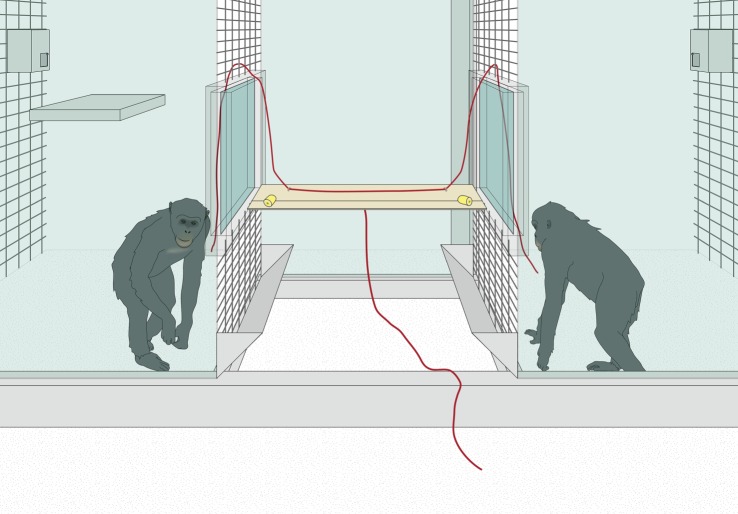
сотрудничества (а точнее – согласовывания действий) у антропоидов «экономической игры» является игра «Охота на оленя» (Stag Hunt game) (Bullinger et al., 2011b). Двое шимпанзе находятся в клетках друг напротив друга и могут независимо друг от друга добывать малоценную для них пищу («зайца»; в данной ситуации – выковыривать изюминки из панели, укреплённой на стене). Через некоторое время у них появляется возможность «поймать» высокоценную пищу («оленя»; в данной ситуации – подтянуть платформу с кусками банана). Последняя ситуация требует обязательных совместных согласованных действий, при этом сольная попытка кого-либо из шимпанзе получить «оленя» приводила к потере всех возможных источников пищи. Кроме того, после того, как шимпанзе отходили от ящика с изюмом, чтобы «поймать оленя», ящик закрывался и становился им недоступен. При одном условии видимость между партнёрами была открытой, в то время как в другой – блокировалась барьером (Duguid et al., 2014).
Другим примером «экономических игр» является «ultimatum game», адаптированная для антропоидов, чтобы продемонстрировать у них наличие или отсутствие «чувства справедливости» (Jensen et al., 2007). В этом случае получить вознаграждение обезьяны могут лишь в случае, когда по очереди подтянут один и тот же поднос. Однако то, какой поднос выбрать для подтягивания, может решить только первый индивид. Он выбирает из двух подносов, пища на которых распределена по-разному – «справедливо» (поровну на две кучки), либо «несправедливо» – на стороне платформы, которая будет ближе к первому индивиду лежит заметно больше кусочков, чем на стороне платформы, который будет ближе ко второму индивиду (например, 8 и 2). Второй индивид может подтянуть выбранную первым платформу так, что еда будет доступна обоим, либо же «отвергнуть» предложенный вариант. В этом случае еда не достанется никому.
При помощи «экономических игр» в особом, разработанном для
животных экспериментальном дизайне, пытались найти эволюционные основы «нравственности» у приматов – имеется ли у них «чувство справедливости» и учитывают ли они «интересы других» (other-regarding preferences), а также исследовали, каковы механизмы согласования скоординированных действий и факторы, ограничивающие сотрудничество, чувство справедливости.
2.2 Обмен жетонов на вознаграждение
В этом варианте исследований эволюционных основ «нравственного» поведения у антропоидов индивиды предварительно обучаются обменивать пищу на жетоны. Как правило, экспериментатор, сидящая перед клеткой, протягивает обезьяне жетон, та берёт его. Сигналом к тому, что жетон можно
обменять на пищу, является повернутая кверху ладонь руки экспериментатора, которую она вытягивает по направлению к окошку в передней стенке клетки.
Наличие у индивидов разных видов приматов «чувства справедливости» и «отвержение ими неравенства» при данной схеме эксперимента проверяется следующим образом: при обмене экспериментатором жетонов на пищевое вознаграждение присутствуют одновременно две обезьяны. После того, как обезьяны протягивают жетон одного и того же типа, они получают еду разного качества (например, огурец/яблоко vs. виноградину). Предполагается, что отказ обменивать свой жетон тем индивидом, который получал худший вариант вознаграждения, может говорить о наличии у него «чувства справедливости/несправедливости». В литературе представлены несколько вариантов подобного рода обменов, как и дискуссии относительно того, как интерпретировать полученные результаты.
Впервые подобный эксперимент был проведѐн Brosnan, de Waal (2003) на капуцинах. В их работе «субъекты» – те индивиды, которым первым обменивали жетон на пищу – всегда получали огурец за жетон, однако их «конкуренты» после этого либо получали тот же тип пищи (условие «справедливости» – equity), либо более желанную пищу за жетон (условие «несправедливости»). Было обнаружено, что «субъекты» отвергали впоследствии потенциальные обмены своих жетонов на менее желанную пищу, если их конкуренты получали лучшую пищу за тот же самый жетон.
Авторы посчитали, что так эти обезьяны отвергают несправедливость «разного вознаграждения за одну и ту же работу» и, следовательно, неприятие несправедливости появляется рано в эволюции. Однако эти данные, а также условия проведения эксперимента были подвергнуты сомнению по большому количеству эмпирических и методологических оснований (в том числе и вследствие «низкой статистической мощности») (напр., Roma et al., 2006; Fontenot et al., 2007), завязалась оживлѐнная дискуссия, были и проведены «улучшенные» экспериментальные исследования (например, Brosnan et al., 2005, 2006), а также исследования на разных видах как человекообразных обезьян, так и других видах приматов. Например, в исследовании с разными видами антропоидов (Bräuer et al., 2011), более часто «несправедливое»
вознаграждение за обмененный жетон отвергали лишь бонобо, но не шимпанзе и не орангутаны.
В целом, создаётся впечатление, что авторы до сих пор не пришли к однозначному решению относительно правильной процедуры проведения подобного рода исследований, а лучшим объяснением наблюдаемого поведения может являться не отвергание неравенства/несправедливости, но ожидание того, что получишь такую же пищу, как и другой индивид, за которым только что пронаблюдал.
Экспериментальная схема по типу «обмена жетонов» использовалась также для изучения «альтруизма». В работе сотрудников группы Fr. de Waal (Horner et al., 2011) у самок шимпанзе была возможность обменивать с экспериментатором либо «эгоистичные» жетоны (в этом случае пищевое вознаграждение получали только они сами), либо «просоциальные» жетоны (в этом случае пищу получали и их партнѐры). Авторы утверждают, что самки шимпанзе имели тенденцию обменивать «просоциальные» жетоны, что свидетельствует, по их мнению, о наличии у шимпанзе «альтруизма».
2.3 Предоставление партнѐру орудий и доступа к месту для получения вознаграждения
В экспериментах, построенных по типу GAT (Giving Assistance Tests) участники-приматы имеют возможность сделать выбор: предоставить «инструментальную помощь» другому или ничего не предпринимать (Amici et al., 2014). F. Warneken и М. Tomasello (2006) создавали ситуации, когда выращенные людьми ювенильные шимпанзе доставали для людей, активно показывающих свою несостоятельность, находящиеся вне их досягаемости предметы (например, залезая на несколько метров вверх). Позднее было продемонстрировано, что шимпанзе могут помогать и конспецификам. [см. видео]
Например, в работе F. Warneken et al. (2007) шимпанзе должен был открыть дверь для своего партнёра, что он и делал более чем в 2 раза чаще, чем в условии «Контроль», когда партнёр либо отсутствовал, либо пытался пройти сквозь другую дверь. Очень важно, что шимпанзе никогда не помогали другому, если тот никак не демонстрировал своѐ намерение пройти через дверь. В дальнейшем было продемонстрировано, что такого рода альтруизм носит «реципрокный» характер: шимпанзе проявляли тенденцию открывать дверь с большей готовностью, если их партнѐр до этого открывал дверь им самим (Melis et al., 2008).
В более поздней работе этой группы авторов (Melis et al., 2011) шимпанзе высвобождали крюк, чтобы пища могла по наклонному пандусу скатиться к находящемуся в клетке напротив них конспецифику, но если только для них было очевидным, что сами они получить пищу не могут, и если реципиент явно сигнализировал о своих потребностях, активно тряся решѐтку. И, наконец, Yamamoto с коллегами (2009) наблюдали, как шимпанзе давали орудия другим, которые нуждались в том, чтобы подгрести пищу, а совсем недавно показали, что шимпанзе демонстрируют «целевую» (targeted) помощь, а именно – дают конкретное орудие, в котором нуждается их конспецифик, выбирая его из ряда возможных (Yamamoto et al., 2012). F. Warneken проводил работы, непосредственно сравнивая «помогающее поведение» у человеческих детей разных возрастов и ювенильных шимпанзе (например, Warneken, 2013; Warneken, Tomasello, 2006, 2007).
2.4 Другие методики.
Jan Engelmann, проводя c соавторами из группы М. Томаселло
сравнительное исследование, использовали особую методику, позволяющую, с их точки зрения, проверять влияние на поведение пятилетних детей и шимпанзе того, «как выглядит твой поступок со стороны» или «что о тебе подумают другие (т.н. «impression management»). В отличие от детей, которые
изменяли своѐ поведение в присутствии свидетеля (беспокоились о своей репутации) – больше делились и меньше украдкой брали себе, если за ними наблюдал сверстник, шимпанзе не меняли своего поведения – они направляли вознаграждение в свою сторону, вне зависимости от присутствия свидетеля (Engelmann et al., 2012).
3. Сотрудничают или решают собственные проблемы при помощи другого?
В целом, при том, что существуют разногласия относительно того, могут ли человекообразные обезьяны кооперироваться на основании понимания необходимости совместных действий и/или альтруистических побуждений, а также понимать вклад каждого в «общее дело» и наказывать «мошенников» (напр., Boesh et al., 2010; Calcutt et al., 2014; Horner et al., 2011), наиболее убедительной выглядит точка зрения, последовательно излагаемая группой М. Томаселло. Согласно ей, сотрудничество у людей должно отвечать следующим критериям:
(1) Понять и согласиться, что цель – общая.
(2) Договориться заранее о комплементарных (взаимодополняющих) ролях.
(3) Хотеть и уметь помочь партнѐру, если тот не может выполнить своей роли.
(4) Важная роль принадлежит намеренной коммуникации с целью координации действий (Moll, Tomasello, 2007; Tomasello, Vaish, 2013).
В соответствии c этим М. Томаселло видит два значимых ограничения «сотрудничества» у антропоидов: коммуникативное и мотивационное.
3.1. Коммуникативное ограничение «сотрудничества» у животных
М. Томаселло настаивает, что одним из ключевых ограничителей подлинного сотрудничества у приматов является отсутствие у них намеренной коммуникации с целью предварительно, заранее скоординировать свои действия. Коммуникативные действия шимпанзе и других антропоидов имеют преимущественно побудительную функцию, при том, что целенаправленное информирование (например, как если бы они сообщали: «я закончил есть изюм и иду вытягивать банан») и декларирование («появилась платформа с едой») отсутствуют (Томаселло, 2011). Иными словами, обезьяны не договариваются между собой заранее, прежде чем начать совместные действия.
Наглядно это было продемонстрировано в экспериментальной ситуации «Охота на Оленя» (Bullinger et al., 2011). Шимпанзе никогда не коммуницировали между собой до того, как они заканчивали есть изюм и шли подтягивать доску с бананом («давай закончим есть изюм и пойдём тянуть доску»….), чтобы скоординировать своё поведение (и обезопасить себя от того, что потеряв «зайца», не достанут «оленя»). Коммуникация происходила лишь после того, как один из шимпанзе находился уже у «оленя» (платформы для подтягивания), тогда он криками и визгами пытался побудить своего партнёра присоединиться к нему. Они тем более вероятно коммуницировали, чем дольше ждали своего партнѐра у доски (Bullinger et al., 2011).
Парадокс заключался в том, что при отсутствии «договорённости о намерениях» уровень координации пары шимпанзе в ситуации «Охота на оленя» был достаточно высок – они успевали подтянуть платформу с вознаграждением в течение трёх минут, пока та не исчезала, в 91% случаев.
Шимпанзе применяли стратегию, которую авторы назвали «Лидер-Последователь»: один индивид рисковал (теряя возможность есть изюм) и шёл к доске первым, вероятно, прогнозируя, что это вынудит другого присоединиться. В целом подобная стратегия является более когнитивно простым способом разрешать проблемы кооперации. Отсутствие предумышленного сообщения о намерениях и активного отслеживания поведения партнёра до принятия решения – характерные для многих координации действий в группе людей – приводило к тому, что по мере совершения всё большего количества проб шимпанзе никак не изменяли стратегии своего «сотрудничества» в сторону максимизации успеха.
Чтобы ещё раз подтвердить эти выводы, сотрудники группы М.
Томаселло (Duguid et al., 2014) недавно продолжили данное
экспериментальное исследование, усложнив для обезьян условия «сотрудничества»: теперь между индивидами находился непрозрачный барьер, а дверца с «зайцем» (который стал более желанным – вместо изюма – бутылочка с соком) захлопывалась бесшумно. Поэтому индивиды не могли видеть и слышать, что делают их партнёры (в частности, едят ли они «зайца» или уже направляются к «оленю») до того, как сами не окажутся рядом с доской для совместного подтягивания вознаграждения. Стратегия «Лидер-Последователь» оказалась в такой ситуации менее эффективной – шимпанзе гораздо менее охотно покидали более значимого для них «зайца», и успешно координировались лишь в 53% случаев. Предварительной коммуникации «о намерениях» до принятия решения оставить «зайца» и идти «ловить оленя» не наблюдалось и в этом случае.
В отличие от шимпанзе, человеческие дети в сходных экспериментальных условиях «Охоты на оленя» координировали свои действия заранее, ещё поедая «зайца», при этом интенсивность коммуникационных обменов значимо возрастала в более сложных условиях – когда не было видно друг друга, а «заяц» был более значимым (Duguid et al., 2014).
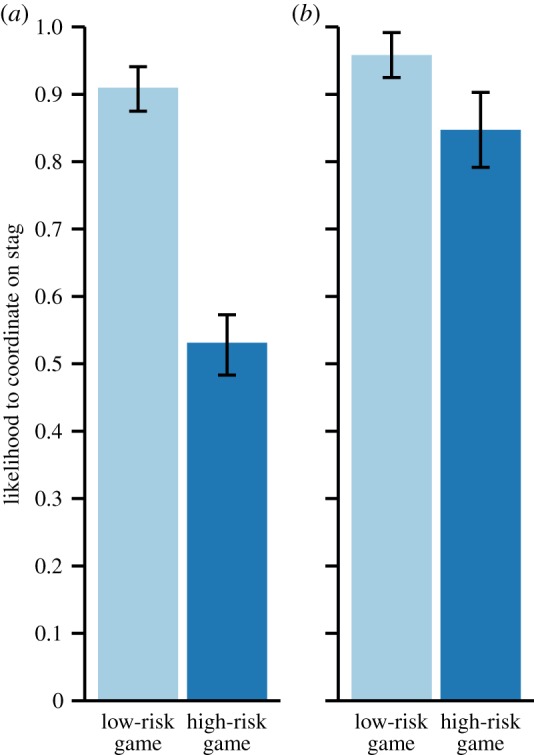
Вероятность кооперации в «охоте на оленя» у шимпанзе (а) и детей (б) при низком и высоком риске игры (и зайца потерять и к оленю не успеть). Diguid et al., 2014
Об отсутствии предварительной коммуникации о намерениях сообщается и в другой, ранее упомянутой нами, работе (Melis et al., 2009). В ситуации, когда доминирующий и подчинённый шимпанзе могли выбирать между «справедливой» (с равным распределением вознаграждения) и «несправедливой» (с неравным распределением вознаграждения) платформой, подчинённый шимпанзе не соглашался тянуть платформу, на которой находилось слишком мало еды с его стороны, а шёл к той платформе, на которой находилось равное количество еды с обеих сторон. Авторы сообщают, что никогда не наблюдали никаких внешне заметных сигналов между партнёрами для координации их конфликтующих предпочтений. Это удивительно, пишут далее они, так как партнёры ждали друг друга у разных поддонов (будучи неспособны договориться о том, какой поддон тянуть) до того момента, когда поддоны убирали (по истечении трѐх минут), прибегая к стратегии «кто кого пересидит». Любой тип коммуникативных усилий повлиять на партнёра мог бы облегчить или ускорить процесс переговоров (например, как жесты, которые шимпанзе используют для вербовки партнёров по коалициям в ходе агрессивных взаимодействий).
Даже в случаях, когда шимпанзе способны «договориться»,
доминирование ограничивает способность шимпанзе решать проблемы кооперативно, если вовлечена делёжка пищей: доминанты не меняют своей стратегии, несмотря на предыдущие отказы подчинѐнных тянуть ту платформу, которую они выбрали. Чем более «несправедливы» условия распределения вознаграждения на платформе, тем больше доминанты упрямятся, и тем более вероятно кооперация разваливается (Melis et al., 2006b).
3.2. Мотивационное ограничение «сотрудничества» у животных
Проводимые в последние несколько лет исследования эволюционных предпосылок «нравственных основ» поведения у животных подвели к постановке вопроса: «действительно ли, сходно с человеческими детьми, шимпанзе находят объединённую активность саму по себе вознаграждающей?».
В целом, шимпанзе (и другие приматы) продемонстрировали достаточно сложные когнитивные способности при решении задач, требующих совместных действий с партнёрами: в экспериментах с подтягиванием платформы шимпанзе понимали необходимость партнёра для успешного решения задачи (Melis et al., 2006a, b, 2008, 2009; Hare et al., 2007), хорошо считали («максимизировали) собственную выгоду и применяли, учитывая её, разные поведенческие тактики (Melis et al., 2009), до определѐнной степени понимали, какое орудие
следует передать своему партнёру для совместной добычи вознаграждения (Melis et al., 2014). Однако, шимпанзе выбирали вариант «сотрудничества» лишь в ситуациях, когда этот выбор был связан с более высоким «выигрышем» для них самих и всегда предпочитали действовать в одиночку, если могли получить точно такое же вознаграждение, вне зависимости от его типа, как и при совместных действиях с другим индивидом (Bullinger et al., 2011). В разных типах экспериментов, даже требующих от участников задействования сложных когнитивных способностей (как, например, в эксперименте Melis et al., 2014, где шимпанзе обменивались друг с другом орудиями, выполняя разные действия по добыче вознаграждения), другой индивид выступал скорее как «социальное орудие» для достижения желанной приманки. Шимпанзе объединяют свои действия с партнёром, но для достижения своих собственных индивидуальных целей; они не интересуются достижением «общих» целей, и если в ходе совместной активности партнёр становится пассивным и теряет интерес к выполнению той или иной задачи, то шимпанзе, как правило, ищут других способов достичь цели или становятся неактивными, редко прилагая усилия, чтобы вновь включить своего партнёра в продолжение данной деятельности (Melis et al., 2009; Warneken et al., 2006).
Что касается «альтруизма», или учитывающего интересы других индивидов поведения («other regarding behavior»), то показательны результаты двух исследований, между которыми почти 10 лет, но которые продемонстрировали сходные результаты (Silk et al., 2005; Amici et al., 2014). В экспериментах учѐных из группы D. Povinelli (Silk et al., 2005; Vonk et al., 2008) шимпанзе могли доставлять пользу другим индивидам, при этом, сами не неся дополнительных затрат. Субъектам – 18 шимпанзе (семь из которых жили вместе на протяжение не менее 15 лет) – предоставлялись на выбор две платформы. Если субъект подтягивал платформу 1, одновременно пищевое вознаграждение получали он и другой шимпанзе (1/1). Если субъект выбирал другой вариант, то сам он получал пищевое вознаграждение того же типа и в таком же количестве, но другой шимпанзе не получал ничего (1/0). В качестве контроля субъектам давалась возможность сделать точно такой же выбор в ситуации, когда другой шимпанзе отсутствовал. Результаты экспериментов показали, что шимпанзе в целом не делали больше т.н. «альтруистических» выборов, даже если их партнѐр проявлял интерес к пищевому вознаграждению, например, вытягивая к нему руку. Никто из 18 протестированных шимпанзе не выбирал вариант 1/1 значимо более часто, когда присутствовал другой шимпанзе, по сравнению с теми пробами, когда они были в одиночестве; они выбирали вариант 1/1 в 56% проб, когда были одни, и в 58% проб, когда присутствовал другой шимпанзе. На вероятность выбрать вариант 1/1 сильно влияло не присутствие другого индивида, а положения платформы: субъекты практически всегда выбирали платформу с вариантом 1/1, когда та находилась с правой, а не с левой стороны от них (в 95% случаев), или, в другом варианте эксперимента – на верхнем ярусе, а на нижнем, в то время как наличие потенциального реципиента усиливало возможность выбора варианта 1/1 лишь на 11%.
При повторении подобной схемы эксперимента на нескольких видах приматов были получены не столь однозначные результаты, как в случае с шимпанзе. Например, бурые черноголовые капуцины (Sapajus apella) и игрунки (Callithrix jacchus) были чутки к потребностям своих партнѐров и предпочитали «просоциальные» варианты аппаратов (Burkart et al., 2007; Takimoto et al., 2010). Авторы делают вывод, что «учитывание интересов других индивидов» может отражать особенности питания или выращивания потомства и быть обнаружено у тех биологических видов, которые полагаются более значительно на кооперативную стратегию при добывании пищи, чем шимпанзе. В работе Amici с сотрудниками (Amici et al., 2014) неожиданным стало то, что некоторые виды приматов (орангутаны, паукообразные обезьяны, капуцины, в отличие от предыдущих исследований) были не просто равнодушны к партнёру в задаче с подтягиванием платформы, но даже
совершали меньше т.н. «просоциальных» выборов (т.е. выбирали вариант 1/1), когда партнёр присутствовал, чем когда их партнѐра не было. Сходные результаты были получены в работах (Jensen et al., 2006; Yamamoto, Tanaka, 2010).
При проведении, после адаптации, данной экспериментальной
парадигмы с детьми, было показано, что 18-месячные дети выбирали варианты 1/0 и 1/1 случайно, 24-месячные дети более часто выбирали варианты 1/1; с 6 лет дети отдавали предпочтение при выборе варианта 1/1 «члену группы» (Brownell et al., 2009).
Альтруизм может проявляться в бескорыстной помощи другому
индивиду. Известно, что шимпанзе делятся пищей с другими при некоторых обстоятельствах: матери – со своими детѐнышами (хотя по большей части они делятся «пассивно», позволяя своему детёнышу забирать у них часть пищи), если пищу нельзя монополизировать (например, ветка с большим количеством листьев), группа обезьян может мирно кормиться, и иногда может происходить более активная делѐжка среди «приятелей». Если пища ценится высоко и отчасти еѐ можно монополизировать (напр., мясо), то тогда обычно не обладающие ею выпрашивают и досаждают обладающим пищей индивидам до тех пор, пока не получают некоторое количество. Как уже отмечалось нами выше, для того, чтобы инициировать «помощь» другим индивидам в экспериментальных ситуациях, шимпанзе требуются прямые сигналы от нуждающегося в помощи (Melis et al., 2011). В тех же самых условиях человеческие дети (но только начиная с трѐх лет) помогают другим, которые могут даже не знать, что нуждаются в данный момент в помощи, делая выводы о целях другого при отсутствии явных внешних сигналов с его стороны (Warneken, 2013).
В целом, когда затраты индивида на «помощь» незначительны, а
реципиент ясно сигнализирует о своём желании, человекообразные обезьяны могут помогать другим. Когда затраты больше, как при делёжке пищей, альтруизм человекообразных обезьян наиболее правдоподобен в контексте
реципрокности, или взаимного обмена. При этом, скорее всего речь идёт о своего рода «реципрокности отношений», или
реципрокности психологических установок на другого, при которой индивиды имеют более позитивные эмоции и, соответственно, ожидания по отношению к тем, кто помогал им или делился с ними в прошлом (de Waal, Suchak, 2010).
Остановимся на итогах исследований «эволюционных корней
справедливости» (sense of fairness). В литературе представлено несколько работ, изучавших влияние совместного (обе обезьяны подтягивают платформу с вознаграждением) или одиночного (одна обезьяна подтягивает, а другая наблюдает) «труда» на распределение пищевого ресурса. Показательной является красиво спланированная экспериментальная работа сотрудников группы М. Томаселло (Melis et al, 2011). Они помещали шимпанзе парами, комбинируя 2 условия в 4 разных варианта: обезьяны могли подтягивать платформу в одиночку или парой, при этом они находились либо в одной вольере, близко друг к другу, либо в разных вольерах, поэтому, чтобы подойти к подтянутой к решётке платформе, второму индивиду требовалось некоторое время, чтобы обогнуть стенку и приблизиться к ней (в то время как первый находился рядом с платформой). Авторы этой работы не обнаружили каких-либо доказательств, что шимпанзе получают больше пищи, когда вносят «трудовой» вклад в совместно решаемую задачу. Однако, на то, как много получал пищи второй индивид, влияло то, где он находился на момент подтягивания платформы – в другой комнате или рядом с первым индивидом.
Даже если он не подтягивал платформу, а просто сидел рядом с тем, кто подтягивал, он успевал захватить больше еды, чем в случае, когда спешил после «работы» из соседней клетки. Также эти индивиды не выпрашивали больше после совместного решения задачи. В целом, «поймавший» и «не поймавший» добычу никак не изменяли своего поведения в зависимости от того, как была «поймана» пища. Авторы не обнаружили явной связи между уровнями настойчивого выпрашивания и делѐжкой: почти не было случаев активной делѐжки; большинство передач пищи носило пассивный характер, инициированный просящими, как уже сообщалось во многих других исследованиях (например, Boesch C., Boesch H., 1989). «Не владеющие» получали немного пищи лишь после того, как сидели в непосредственной близости от обладателя, тянулись к фрукту и/или к его обладателю, собирали кусочки, по большей части шелуху, оброненную обладателем (Melis et al.,
2011).
В исследовании, изучающем поведение делѐжки маленьких детей Katharina Hamann с сотрудниками группы М. Томаселло задавали 3 разных условия: «Сотрудничество» (дети должны были одновременно подтягивать за две верѐвки платформу с вознаграждением), «Параллельная работа» (каждый подтягивал собственную платформу), «Ничего не делали» (вознаграждение
становилось доступным без каких-либо действий со стороны детей –платформа уже была подтянута). Дети делились поровну (если одному доставалось вознаграждение, а другому – нет) в зависимости от своего возраста. Трёхлетние дети: 75% пар уравнивали призы в условии «Сотрудничество», 25% – в условии «Параллельная работа», 5% – в условии «Ничего не делали». У двухлетних детей различий при делѐжке в условиях
«Сотрудничество» и «Параллельная работа» не было.
Проведя множество исследований, М. Томаселло приходит к выводу, что то, что мы наблюдаем у наших ближайших из ныне живущих родственников – человекообразных обезьян – не является «подлинным сотрудничеством», с пониманием и сопереживанием нужд партнёров, с выработкой общей цели и распределением обязанностей по еѐ выполнению, а также готовностью взять, в случае необходимости, на себя труд по замещению товарища – т.е. того, благодаря чему возможны культура, еѐ кумуляция, и достижения современного социо-культурного и технического прогресса у человека в целом. У антропоидов нет способности в полной мере понимать представления других о текущей ситуации и использовать это понимание для планирования своих действий (theory of mind), но главными ограничениями возможностей «сотрудничества» у антропоидов, как считает М. Томаселло сегодня, являются скорее мотивационные, чем когнитивные, факторы – подлинная готовность помогать, делиться и жертвовать собой ради интересов другого. Чтобы сопереживать другому, необходимо не только понимать, почему тот страдает, но, что очень важно, и желать учитывать это при принятии решений (Томаселло, 2011; Melis et al., 2014; Tomasello, Vaish, 2013). Антропоиды, и другие животные, могут эффективно действовать, учитывая поведение других индивидов, однако действовать скоординировано далеко не всего означает сотрудничать (cooperation ≠ collaboration).
М. Томаселло считает, что сотрудничество не является обязательной частью жизни приматов, а социально-когнитивные
навыки у обезьян эволюционировали как всё более изощрѐнная конкуренция, но не сотрудничество (Moll, Tomasello, 2007).
В заключение
Представленными нами работами, конечно же, не исчерпывается ни огромный объём современных экспериментальных и проводящихся всѐ чаще вне лабораторий, в природе, исследований социо-когнитивных возможностей приматов, ни разнообразие всех существующих ныне точек зрения на то, насколько близки нам «нравственные основы» поведения наших ближайших из ныне живущих родственников среди животных. В статье сделан акцент на теоретических взглядах одного из ведущих специалистов сравнительной психологии – Майкла Томаселло, которые близки нам по ряду причин, одной из которых является сходство его видения данной проблемы отечественной школе культурно-исторической психологии, в том числе представлениям Л.С. Выготского и А.Н. Леонтьева.
Значительным недостатком существующего ныне массива данных и теорий, построенных на их основании, является то, что вопросы предыстории «сотрудничества» изучались преимущественно на примере одного биологического вида – шимпанзе, тогда как существуют лишь отдельные работы по рассматриваемой тематике с участием других видов приматов. В
связи с этим имеется огромный риск, что представленные ограничения и особенности «нравственных эволюционных основ» поведения шимпанзе могут объясняться в значительной степени особенностями образа жизни именно данного вида.
Проведённый нами обзор основан, за редким исключением, на
литературных источниках, свободно размещённых в Интернете, поэтому заинтересовавшиеся ими читатели сами могут ознакомиться с нюансами организации экспериментов и более подробно представленными результатами.
Как нам представляется, многие из разработанных экспериментальных дизайнов можно использовать в работе с содержащимися в зоопарках приматами с целями не только, и даже не столько научными, сколько для обогащения и повышения благополучия жизни этих высокоорганизованных животных. Однако, делать это надо с осторожностью, при обязательном мониторинге влияния подобных «экспериментальных интервенций» человека на жизнь животных (как было показано в одной из работ, представленных в данном сборнике (Федорович, Подтуркин), не всегда «когнитивное обогащение» приматов может приводить к желанной динамике социальных взаимодействий в группе).
Литература
Томаселло М. 2011. Истоки человеческого общения. – М.
Amici F., Visalberghi E., Call J. 2014. Lack of prosociality in great apes, capuchin monkeys and distribution tasks spider monkeys: convergent evidence from two different food: //Proc. R. Soc. B. Vol. 281, 20141699, http://dx.doi.org/10.10998 rspb.2014.1699
Boesch C., Boesch H. 1989. Hunting behavior of wild chimpanzees in the Tai National Park //Am. J. Phys. Anthropol. Vol. 78. P. 547-573.
Boesch C., Bole ́C., Eckhardt N., Boesch H. 2010. Altruism in Forest Chimpanzees: The Case of Adoption. PLoS ONE 5 (1): e8901. doi:10.1371/journal.pone.0008901
Bräuer J., Call J., Tomasello M. 2009. Are Apes Inequity Averse? New Data on the Token-Exchange Paradigm // American Journal of Primatology 71:175–181.
Brosnan S.F., Shiff H.C., de Waal F.B. 2005. Tolerance for inequity may increase with social closeness in chimpanzees // Proc Biol. Sci. Vol. 272. P. 253-258.
Brosnan S.F., de Waal F.B. 2006. Partial support from a nonreplication: comment in Roma, Silberburg, Ruggiero, Suomi //J. Comp.Psychol. Vol. 120. P. 74-75.
Brownell C., Svetlova M., Nichols S. 2009. To Share or Not to Share: When Do Toddlers Respond to Another‘s Needs? // Infancy. Vol. 14. P. 1–14.
Byrne R.W., Whiten A. 1988. Machiavellian intelligence. Social experience and the evolution of intellect in monkeys, apes, and humans. New York. Oxford University Press.
Bullinger AF, Melis AP, Tomasello M. 2011а Chimpanzees (Pan troglodytes) prefer individual over cooperative strategies toward goals. Animal Behaviour. Vol. 82. P. 1135–1141.
Bullinger A.F., Wyman E., Melis A.P., Tomasello M. 2011b. Coordination of Chimpanzees (Pan troglodytes) in a Stag Hunt Game // Int. J. Primatol. Vol. Vol. 132 (6). P. 1296-1310.
Burkart J.M., Fehr E., Efferson C., van Schaik C.P. 2007. Other-regarding preferences in a non-human primate: common marmosets provision food altruistically // Proc. Natl Acad. Sci. USA. Vol. 104. P. 19762-19766.
Calcutt S.E., Lonsdorf E., Bonnie K., Milstein M., Ross S. 2014. Captive chimpanzees share diminishing resources // Behaviour. Vol. 151. P. 1967-1982.
De Waal F.B., Suchak M. 2010. Prosocial primates: selfish and unselfish motivations // Philos Trans R Soc Lond B Biol Sci, Vol. 365 (1553). P. 2711-2722.
Duguid S., Wyman E. , Bullinger A., Herfurth-Majstorovic K., Tomasello M. 2014. Coordination chimpanzees and human children in a Stug Hunt game // Proccedings of the Royal Society B: Biological Series. Vol. 281 (1796). 20141973.
Engelmann J.M., Herrmann E., Tomasello M. 2012. Five-Year Olds, but Not Chimpanzees, Attempt to Manage Their Reputations // PLoS ONE 7(10): e48433.doi:10.1371/journal.pone.0048433.
Fontenot M.B., Watson S.L., Roberts K.A., Miller R.W. 2007. Effects of food preferences on token exchange and behavioural responses to inequality in tufted capuchin monkeys, Cebus
paella // Anim Behaviour. Vol. 74. P. 487–496.
Hare В. 2011. From Hominoid to Hominid Mind: What Changed and Why? // Annu. Rev. Anthropol. Vol. 40. P. 293–309.
Hamann К., Warneken F., Greenberg J. R., Michael Tomasello M. 2011. Collaboration encourages equal sharing in children but not in chimpanzees // Nature. Vol. 476 (7360). P. 328-331.
Horner V., Carter D., Suchak M., de Waal F.B. 2011. Spontaneous prosocial choice by chimpanzees // PNAS. Vol. 108. # 33. P. 13847-13851.
Humphrey N.K. 1976. The social function of intellect. In: «Growing Points in Ethology,» ed. P.P.G. Bateson and R.A. Hinde, pp. 303- 317, Cambridge University Press, Cambridge.
Jensen K., Call J., Tomasello M. 2007. Chimpanzees are rational maximizers in an ultimatum game // Science. Vol. 318. P. 107-109.
Jensen K., Hare B., Call J., Tomasello M. 2006. What‘s in it for me? Self-regard precludes altruism and spite in chimpanzees. Proc. R. Soc. B. Vol. 273. P. 1013–1021.
Moll H., Tomasello M. 2007. Cooperation and human cognition: the Vygotskian intelligence hypothesis // Phil. Trans. R. Soc. B. 362, 639–648.
Melis A. P., Hare B., Tomasello M. 2006 a. Chimpanzees recruit the best collaborators // Science. Vol. 311. P. 1297–1300.
Melis A. P., Hare B., Tomasello, M. 2006b. Engineering cooperation in chimpanzees: Tolerance constraints on cooperation // Animal Behaviour. Vol. 72 (2). P. 275–286.
Melis, A.P., Hare B., Tomasello M. 2008. Do chimpanzees reciprocate received favours? // Animal Behavior. Vol. 76. P. 951–962.
Melis A.P., Hare B., Tomasello М. 2009. Chimpanzees coordinate in a negotiation game // Evolution and Human Behavior. Vol.30. P. 381–392.
Melis A. P., Schneider A.- N., Tomasello М. 2011. Chimpanzees, Pan troglodytes, share food in the same way after collaborative and individual food acquisition // Animal Behaviour. Vol.82. P. 485-493.
Moll H., Tomasello M. 2007. Cooperation and human cognition: the Vygotskian intelligence hypothesis // Phil. Trans. R. Soc. B. 362, 639–648.
Premack D.G., Wooruff G. 1978. Does the chimpanxee have a theor of mind? // Behaviorla and Brain Science. Vol. 1. P. 515-526.
Roma P.G., Silberberg A., Ruggiero A.M., Suomi S.J. 2006. Capuchin monkeys, inequity aversion, and the frustration effect. J Comp Psychol 120: 67–73.
Shettleworth S. J. 2010. Cognition, Evolution, and Behavior (Second Edition). Oxford.
Silk J.B., Brosnan S.F., Vonk J., Henrich J., Povinelli D.J., Richardson A.S., Lambeth S.P., Mascaro J., Schapiro S.J. 2005 Chimpanzees are indifferent to the welfare of unrelated group members // Nature. Vol. 437. P. 1357–1359.
Takimoto A., Kuroshima H., Fujita K. 2010. Capuchin monkeys (Cebus apella) are sensitive to others‘reward: an experimental analysis of food-choice for conspecifics // Anim. Cogn. Vol.13. P. 249–261.
Tomasello М., Vaish A. 2013. Origins if human cooperation and morality Annual // Reviews of Psychology. Vol.64. P. 231-255.
Tomasello M., Carpenter M. 2007. Shared intentionality //Developmental Science. Vol. 10 (1). P. 121-125.
Vonk J., Brosnan S.F., Silk J.B., Henrich J., Richardson A.S., Lambeth S.P., Schapiro S.J., Povinelli D.J. 2008. Chimpanzees do not take advantage of very low cost opportunities to deliver food to unrelated group members // Animal Behaviour. Vol. 75. P. 1757–1770.
Warneken F. 2013. Young children proactively remedy unnoticed accidents // Cognition. Vol. 26 (1). P. 101–108.
Warneken F., Hare B., Melis A., Hanus D., Tomasello M. 2007. Spontaneous altruism by chimpanzees and young children // PLoS Biol. Vol. 5 (7). P. 1414–1420.
Warneken, F., Tomasello, M. 2006. Altruistic Helping in Human Infants and Young Chimpanzees // Science. Vol. 311. P. 1301–1303.
Yamamoto S., Tanaka M. 2010. The influence of kin relationship and reciprocal context on chimpanzees‘ other-regarding preferences // Animal Behaviour. Vol. 79. P., 595–602.
Yamamoto S., Humle T., Tanaka M. 2012. Chimpanzees‘ flexible targeted helping based on an understanding of conspecifics‘ goals. Proc. Natl. Acad. Sci. USA 109, 3588–3592.
Вопросы прикладной приматологии. Выпуск 2. // Межвед. сб. науч. и науч.-метод. тр. – М: Московский зоопарк, 2015. С.33-49.
Рекомендуем прочесть
 Влияние «Великой Рецессии» на неравенство материального положения в обществе с учетом возрастной и расовой принадлежности
Влияние «Великой Рецессии» на неравенство материального положения в обществе с учетом возрастной и расовой принадлежности Горим…
Горим… Теоретические основы единой стратегии охраны природы и природопользования
Теоретические основы единой стратегии охраны природы и природопользования Приступ франкофобии
Приступ франкофобии «На женском дне»: старая статья на фоне советского акмэ
«На женском дне»: старая статья на фоне советского акмэ Закон о «запрете» заемного труда дискриминирует одиноких и многодетных родителей
Закон о «запрете» заемного труда дискриминирует одиноких и многодетных родителей Причина гибели сайгаков в Казахстане
Причина гибели сайгаков в Казахстане Экосистемные последствия лесоэксплуатации: Восточная Финляндия vs Карелия
Экосистемные последствия лесоэксплуатации: Восточная Финляндия vs Карелия
