

Некоторые млекопитающие позднего мела — раннего палеоцена Северной Америки: слева — эодельфис (Eodelphis), вверху — птилоды (Ptilodus), справа — гобиаконодоны (Gobiaconodon).
Дэвид Гроссникл, Дэвид Полли
Открытия ископаемых за последние 30 лет радикально изменили традиционные взгляды на эволюцию мезозойских млекопитающих. Кроме того, недавние исследования дали более подробную картину мелового распространения цветковых растений. Здесь мы разбираем схемы морфологического разнообразия и функциональной морфологии, связанные с диетой ранних млекопитающих. Было выполнено два анализа: (i) изучение разнообразия, основанное на функциональном типе зубов, а не на таксономии более высокого уровня, и (ii) морфометрический анализ челюстей, с использованием современных аналогов, чтобы найти изменения в морфологическом и диетарном разнообразии млекопитающих. Результаты показывают сокращение разнообразия типов моляров в середине мела, вместе с уменьшением числа триконодонтов, симметродонтов, докодонтов и эупантотериев. У многобугорчатых в середине мела произошла смена в функциональных типах моляров, и в конце позднего мела — переход к растительным диетам. Хотя терии прошли через таксономическое расширение, совпадающее по времени с распространением покрытосеменных, они показывают малый размер тела и низкий уровень морфологического разнообразия, что говорит об эволюционном сдвиге в пользу мелких насекомоядов. Вывод тот, что в среднем мелу, в период стремительного распространения покрытосеменных, млекопитающие подверглись и сокращению разнообразия морфологии, и функциональному сдвигу в диетах, что, вероятно, связано с изменениями в экосистемах.
Введение
Новые открытия последних 30 лет показали, что мезозойские млекопитающие занимали многие экоморфологические ниши, что и современные представители этой клады, что говорит о том, что они достигли значительного разнообразия задолго до кайнозоя. Новые данные о мезозойских млекопитающих сделали возможным установление масштабов и изменений их морфологического разнообразия и его связь с экологическими изменениями в меловом периоде.
Молекулярные свидетельства помещают начало покрытосеменных не позднее, чем в конец триаса. Их пыльца впервые появляется в валанге-готериве (139-131м.л.н.), и макрофоссилии в барреме — самом начале апта (131-124м.л.н.). Но по большинству исследований, цветковые растения не становились разнообразными или многочисленными до конца апта или альба, когда они быстро распространились и стали значительной частью позднемеловой флоры. Подъём покрытосеменных считается самым важным событием «меловой наземной революции» (МНР), по Ллойду, периода около 125-80м.л.н., когда распространялись клады цветковых растений, млекопитающих, ящериц, социальных насекомых и птиц. Насекомые, особенно отряды Coleoptera, Hymenoptera и Lepidoptera, по-видимому, коэволюционировали и расселялись вместе с покрытосеменными.
Будучи таксономически разнообразными в среднем мелу, покрытосеменные могли и не быть экологическими доминантами до его конца. Эта работа делает упор на их таксономическую диверсификацию (118-90м.л.н.), но стоит учесть, что их эволюционные изменения в конце позднего мела могли иметь более прямое влияние на эволюцию диет млекопитающих.
Одновременность распространения покрытосеменных и современных клад млекопитающих поддерживается данными молекулярных часов, и предполагает, что последние развивались в ответ на распространение покрытосеменных и насекомых.
Считать середину мела временем диверсификации млекопитающих, однако, может быть не вполне точно. Молекулярное исследование Мередита показывает её гораздо больший темп в конце МНР (примерно 80м.л.н), чем во время распространения покрытосеменных.
Корреляция между покрытосеменными и выживанием или вымиранием клад млекопитающих может быть прямой (например, на растительноядных повлияла смена источников пищи); косвенной (например, на насекомоядных влияли опылявшие цветы насекомые); и случайной (выживание было связано с факторами, с покрытосеменными не связанными, например, с уровнем метаболизма).
В этой работе мы изучаем, как изменения в морфологии меловых млекопитающих связаны с распространением покрытосеменных. Упирая на функциональные черты, а не таксономическое разнообразие, мы фокусируемся на тех аспектах их эволюции, что развивались прямо или косвенно вместе с изменениями в растительных сообществах. Были составлены кривые относительного разнообразия родов, принадлежащих к 9 функциональным типам. Изменения в разнообразии этих функциональных групп отражают независимую от таксонов смену пищевых специализаций, в отличие от стандартного подхода подсчёта высших таксонов, что может и не иметь экологического значения. Геометрический морфометрический анализ использован для количественного учёта морфологических изменений в челюстях мезозойских млекопитающих, связанных с их диетой путём установления связи между формой и диетой у современных млекопитающих. Длины челюстей и моляров использованы как приближения для размеров тела, которые имеют функциональные и физиологические следствия для диеты млекопитающих.
Материал и методы
(a)Разнообразие функциональных типов зубов
Данные таксономии и временные рамки для мезозойских млекопитающих взяты в основном из Paleobiology Database, с некоторыми дополнениями из первичной литературы и Келан-Яворовской. Данные можно посмотреть в дополнительном материале. Данные о разнообразии основаны только на числе родов в промежутках в 7 м.л.н. Встречаемость в среднее время всех находок рода использовалась для определения его таксономических рамок. Недавние исследования предупреждают, что за пики и плато в разнообразии меловых динозавров могут быть ответственны искажения сбора и сохранности. Так как данные о разнообразии млекопитающих, скорее всего, искажены похоже, мы измерили относительное, а не абсолютное разнообразие, чтобы минимизировать влияние ошибки сэмплирования. Веретенообразные кривые разнообразия на рис.1 сделаны с помощью MATHEMATICA Вольфрама и QUANTITATIVE PALEONTOLOGY v.2.0 Полли. Данные о разнообразии родов мезозойских млекопитающих категоризированы по функциональному типу зубов (см. дополнительные материалы), которые определены главным образом по молярной морфологии (см. там же).
b)Геометрическая морфометрика и m1-длины
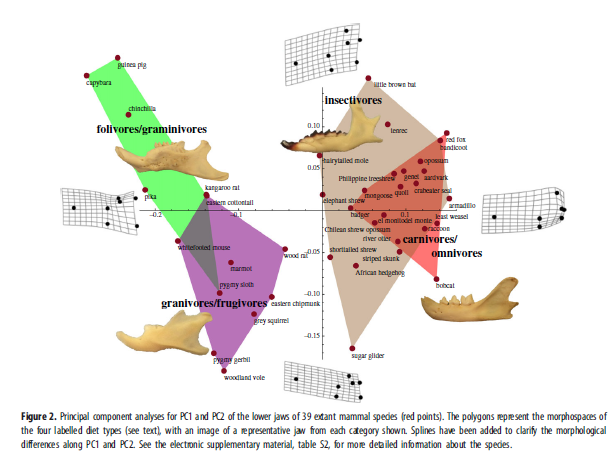
Изучались нижние челюсти 39 современных видов млекопитающих, принадлежащих 32 семействам и 18 отрядам. Щёчные поверхности челюстей 22 видов были сфотографированы в William R.Adams Zooarchaeology Laboratory в Индианском университете. 17 изображений челюстей были взяты из peer-reviewed источников (см. дополнительные материалы). Смотрите дополнительные материалы для обсуждения причин выбора современных видов, использованных в этой работе. Каждый современный вид отнесён к одному из четырёх типов питания: (i)насекомоядные; (ii)хищники и плотоядные всеядные, (iii) листоядные и травоядные, и (iv) фруктоядные и семеноядные (см. дополнительные материалы). Неплотоядные всеядные, что имеют широкий спектр морфологий, были отнесены к одной из этих четырёх категорий на основании их главного источника пищи. Изображения челюстей 87 вымерших родов млекопитающих взяты из первичной литературы и Келан-Яворовской (см. дополнительные материалы). Они включают оригинальные фотографии и реконструкции, и использовались только те, на которых видно большую часть челюсти (около 80% или больше, где как минимум пять из семи отметок видны ясно). Десять родов жили дольше одной эпохи, и их челюсти использовались в каждой временной ячейке, где они встречались. Так вышло 97 пунктов, использованных для получения результатов, показанных на рис.3, хотя было проанализировано 87 отдельных челюстей. На основании функциональной морфологии было выбрано семь ключевых точек (см. дополнительные материалы). Изображения челюстей проанализированы с использованием геометрических морфометрических процедур, таких, как анализ Procrustes и анализ основных компонентов, использующий MATHEMATICA Вольфрама и MORPHOMETRICS v.8.6.4 Полли.
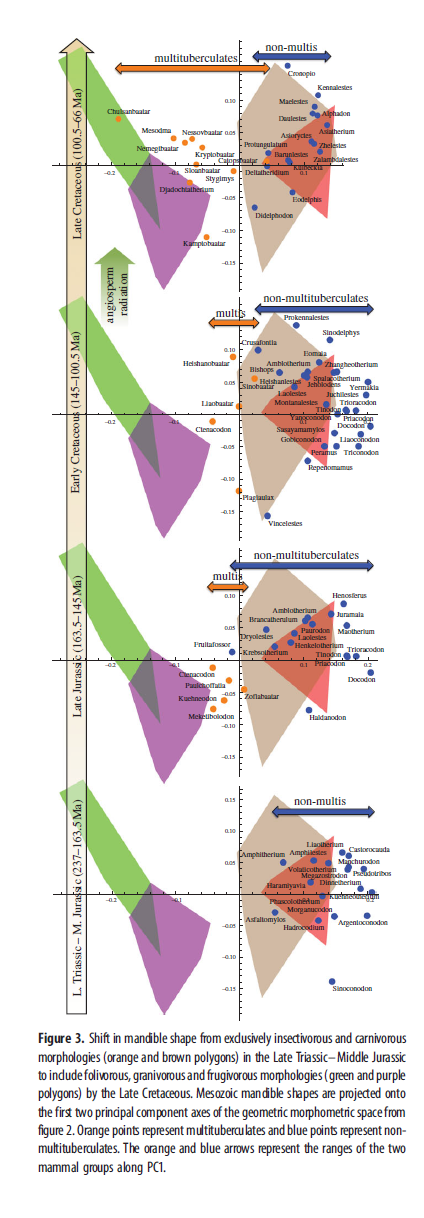
Результаты морфопространства для челюстей современных млекопитающих были представлены в пространстве двух главных компонент PC1 и PC2. Результаты PC1 и PC2 геометрического морфометрического анализа для челюстей вымерших животных были спроецированы на морфопространство, созданное анализом челюстей современных видов. Проецирование вместо сравнения данных позволило сравнить морфологии вымерших и здравствующих видов без искажения данных для последних. Данные для вымерших видов также были разделены по эпохам, кроме периода от позднего триаса до средней юры, которые объединены в одну временную ячейку из-за малого числа образцов (рис.3).
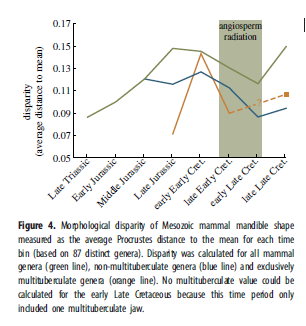
Смотри дополнительные материалы для абсолютного времени временных ячеек, использованные на рис.3–5. Прокрустова форма консенсуса для каждой ячейки вымерших видов был определён и морфологическое разнообразие вычислено как средняя прокрустова дистанция от каждой формы до её среднего значения для каждой ячейки. Для сбора длин m1 для 135 родов мезозойских млекопитающих и длин челюстей 90 их родов взяты из первичной литературы и Келан-Яворовской (см. дополнительные материалы). Разнообразие вычисляется как среднее расстояние между образцами и средним значением для каждой временной ячейки. Стандартные ошибки для длин моляров и челюстей вычислены из 1000 повторений начальных загрузок, при которых каждая временная ячейка была ресэмплирована, и среднее значение пересчитано.
Результаты
Рис.1 показывает относительное разнообразие функциональных групп зубов с поздней средней юры по мел. Точная датировка изменений разнообразия млекопитающих отчасти осложнена небольшим количеством образцов и ограниченным числом местностей с меловыми ископаемыми. Всё же, можно увидеть основные принципы. Млекопитающие с трибосфеническими молярами, особенно эвтерии и метатерии, в мелу резко увеличили разнообразие, при том, что триконодонты, симметродонты и эупантотерии (стем-кладотерии) убывали. Многобугорчатые в среднем мелу перенесли переход от плагиаулакоидных саблевидных к цимолодонтным зубам. Челюсти мезозойских млекопитающих были сравнены с челюстями нынешних аналогичных видов, чтобы определить тип диеты. Были получены результаты геометрического морфометрического анализа для современных линий и PC1 и PC2, что позволило определить морфопространства для четырёх типов диет (рис.2). Есть ясная морфологическая разница между видами хищников (насекомоядные и хищные/всеядные на рис.2) и растительноядных (листо/зерноядные и семя/фруктоядные на рис.2).
Хищники и плотоядные всеядные сильно пересекаются с насекомоядными, если используются любые из первых четырёх главных компонентов. Итоги анализа ANOVA указывают, что эта вариация, связываемая с типом диеты, по вычислениям имела следующие количества: PC1¼38.1%,PC2¼6.9%,PC3¼2.3% и все другие основные компоненты меньше 1%. Таким образом, положение по x-оси (PC1) кажется наилучшим указателем типа диеты. Одно значимое различие между челюстями растительноядных и хищников — расстояние между угловым и мыщелковым выростами, лучше всего различимое по оси PC1. Размер короноидного выроста также значительно меняется по PC1, убывая по длине и глубине по направлению к растительноядному морфопространству. Распределения и образцы челюстей на рис.2 показывают эти различия. Вырисовывается уменьшение разнообразия и частичное сокращение вариативности в хищной/насекомоядной области морфопространства маммальной челюсти в середине мела, за чем последовало расширение растительноядных морфологий.
Это видно на рис.3, который показывает мезозойские таксоны, сгруппированные в четыре временные ячейки, спроецированные на современные морфопространства, где растительноядные морфологии слева, и все прочие справа. Прежде среднемелового распространения покрытосеменных большинство млекопитающих попадало в правую сторону этого пространства. В мелу разнообразие не-многобугорчатых функциональных типов уменьшилось, оставив в основном трибосфенические типы, и их морфопространство сократилось. К позднему мелу, похоже, наличествовали и насекомоядные, и хищные морфологии. Они не очень чётко различимы строением челюсти, оно похоже при этих типах диет, но их можно различить по размеру. В наших образцах современных млекопитающих, средняя длина челюсти насекомоядных значительно короче, чем у хищников и всеядных (см. дополнительные материалы). Из позднемеловых родов, включенных в морфометрический анализ челюсти (рис.3), пять трибосфенических родов, живших в начале позднего мела в насекомоядный размерный класс входят: Alphadon, Barunlestes, Kennalestes, Kulbeckia и Zhelestes (размер тела основан на данных челюстей и зубов, которые есть в дополнительных не-многобугорчатых таксонах в рис.5 и в дополнительные материалы). Их малое разнообразие (рис.4) и малый размер тела предполагает, что эти трибосфеники были ограничены экоморфопространствами небольших насекомоядных во время распространения покрытосеменных. Позднемеловые трибосфеники, такие, как Didelphodon и Deltatheridium, напротив, демонстрируют рост среднего размера тела (рис.5), что указывает на хищную или всеядную диету.
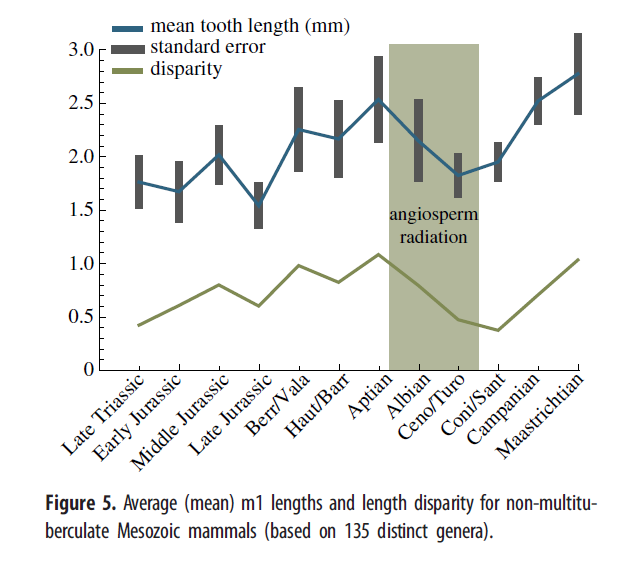 Точная датировка позднемелового перехода к растительноядности у цимолодонтных многобугорчатых трудна, поскольку, к несчастью, в начале позднего мела кроме мезодмы идёт пробел в хронике многобугорчатых. Морфологическое разнообразие в форме нижней челюсти, судя по геометрическому морфометрическому анализу, убывало во время распространения покрытосеменных (рис.4). Надо заметить, данные многобугорчатых основаны на сравнительно малом числе образцов: пять изображений из поздней юры, три из начала раннего мела, четыре из конца раннего мела, один из начала позднего мела (не включен в рис.4) и 10 из конца позднего мела. Практическое отсутствие образцов многобугорчатых в начале позднего мела может помочь оценить общий упадок морфологической вариативности челюсти в то время. Рис.5 и дополнительные материалы, показывают изменения в абсолютной длине и разброс длин (среднее отклонение длины от среднего значения) для первого нижнего моляра (m1) и челюсти не-многобугорчатых мезозойских млекопитающих. Длина и разброс длин также убывает с распространением покрытосеменных. Эти результаты работают как приближение и для размера тела и для разнообразия. Многобугорчатые не участвовали в вычислениях m1 и длины челюсти, но результаты Уилсона показывают, что многобугорчатые оставались относительно мелкими до конца позднего мела, когда произошёл значительный прирост в размерах тела и разнообразии строения.
Точная датировка позднемелового перехода к растительноядности у цимолодонтных многобугорчатых трудна, поскольку, к несчастью, в начале позднего мела кроме мезодмы идёт пробел в хронике многобугорчатых. Морфологическое разнообразие в форме нижней челюсти, судя по геометрическому морфометрическому анализу, убывало во время распространения покрытосеменных (рис.4). Надо заметить, данные многобугорчатых основаны на сравнительно малом числе образцов: пять изображений из поздней юры, три из начала раннего мела, четыре из конца раннего мела, один из начала позднего мела (не включен в рис.4) и 10 из конца позднего мела. Практическое отсутствие образцов многобугорчатых в начале позднего мела может помочь оценить общий упадок морфологической вариативности челюсти в то время. Рис.5 и дополнительные материалы, показывают изменения в абсолютной длине и разброс длин (среднее отклонение длины от среднего значения) для первого нижнего моляра (m1) и челюсти не-многобугорчатых мезозойских млекопитающих. Длина и разброс длин также убывает с распространением покрытосеменных. Эти результаты работают как приближение и для размера тела и для разнообразия. Многобугорчатые не участвовали в вычислениях m1 и длины челюсти, но результаты Уилсона показывают, что многобугорчатые оставались относительно мелкими до конца позднего мела, когда произошёл значительный прирост в размерах тела и разнообразии строения.
Обсуждение
(a)Разнообразие функциональных типов зубов
Итоги на рис.1 демонстрируют смену функциональных типов зубов в течение мела. Это наблюдение поддерживается двумя тенденциями: (i)трибосфенические рода (преимущественно терии) как будто замещают триконодонтов, симметродонтов и эупантотериев, и (ii)цимолодонты замещают плагиаулакоидов как основных многобугорчатых. Смена выглядит наиболее резкой во время распространения покрытосеменных (прим. 118–90Ma) и МНР (прим.125–80Ma). Этот вывод о меловом маммальном перевороте согласуется с результатами Бенсона, хотя тот полагает, что переворот случился в начале позднего мела (100–83.6Ma). Среднемеловые перемены в разнообразии функциональных типов зубов могут быть интерпретированы как отражающие общее уменьшение разнообразия зубов млекопитающих. В раннем мелу были представлены многочисленные типы, например трибосфеники, эупантотерии, триконодонты, симметродонты, докодонты и плагиаулакоиды. В маммальной фауне конца позднего мела, с другой стороны, доминировало меньшее число функциональных типов зубов, особенно трибосфеники, эупантотерии и цимолодонты (рис.1).
Скудость среднемеловой хроники ископаемых может быть отчасти ответственна за это сокращение, но и представленные лучше скалы конца позднего мела тоже показывают низкое разнообразие зубов. Этот упадок также отражён уменьшением длины и разброса длин моляров у не-многобугорчатых (рис.5). Рис.1 показывает небольшой подъём эупантотериев (стем-кладотериев) после роста покрытосеменных. Надо заметить, что это может быть результатом биогеографической полуизоляции Южной Америки в мелу, что привело к эндемическому распространению дриолестоидов, которые обладали многими зубными характеристиками трибосфенических моляров, близко родственны териям, и считаются маленькими насекомоядами. Терии неизвестны из мезозоя Южной Америки. Дриолестоиды могли занимать там те же экоморфологические ниши, что и распространявшиеся повсеместно териевые млекопитающие. Взгляд на рис.1 с этим соображением в уме даёт ещё большую поддержку теории уменьшения разнообразия млекопитающих во время распространения покрытосеменных в целом. Можно представить, что терии распространились сразу же как возникли, быстро захватив ниши в течение среднего мела. Однако ранний терий, юрамайя, был найден в породах, датируемых 160м.л.н., примерно за 40млн. до распространения териев. Кроме того, две работы о молекулярных часах дали приблизительную оценку их появления не позднее 190м.л.н.. Если териевые черты (напр., трибосфенические моляры и внутреннее развитие потомства) сделали их более эволюционно приспособленными, чем другие клады млекопитающих, можно спросить, почему они не могли распространиться намного раньше. Но наши данные показывают, что терии имели значительную радиацию во время распространения покрытосеменных и МНР. Одновременность этих событий поддерживает гипотезу, что процветание цветковых растений сыграло роль, прямую или нет, в успехе териев.
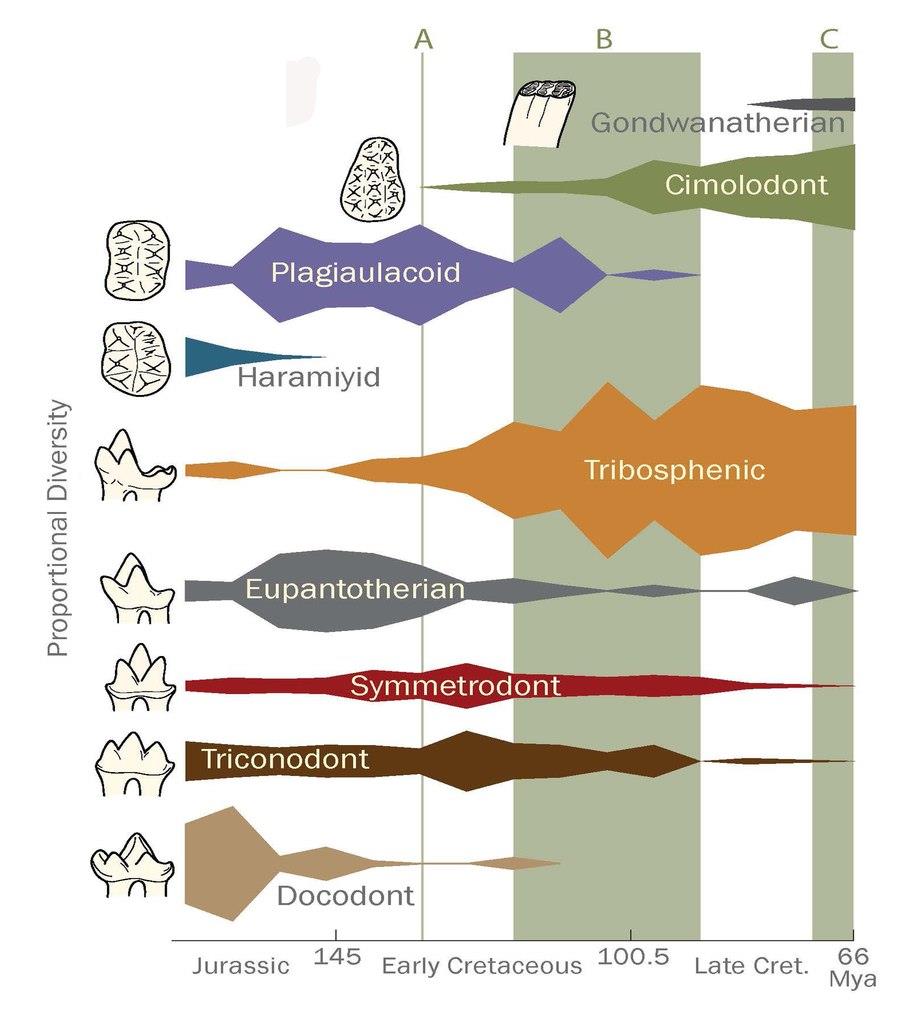
Рис. 1. Относительное родовое разнообразие различных групп мезозойских млекопитающих.
(b)Схемы морфологии челюстей
На основании челюстной морфологии, позднемеловые многобугорчатые, в среднем, морфометрически более похожи на морфопространства современных растительноядных, чем их более ранние представители (рис.3). Некоторые из этих многобугорчатых, например Chulsanbaatar и Nemegtbaatar, находятся в (или близки к) морфопространствах современных листо/зерноядов, что указывает на сдвиг в листоядную экологическую нишу. Эриксон показал палеоботанические свидетельства роста распространения семян животными в позднем мелу. Положение морфопространств таких родов, как Kamptobaatar и Djadochtatherium в или рядом с семя/фруктоядными морфопространствами поддерживает гипотезу, что млекопитающие помогали покрытосеменным в распространении семён. Уилсон продемонстрировал, что сложность зубов многобугорчатых, их разнообразие, средний размер тела и разброс размеров тела не рос существенно до конца начального распространения покрытосеменных, намекая, что их появление не имело непосредственного эффекта на морфологию и диету многобугорчатых. 9 из 10 позднемеловых многобугорчатых в геометрическом морфометрическом анализе челюстей исключительно из кампана (83.5–70.6м.л.н) в конце позднего мела, то есть существенно позже начального распространения покрытосеменных, что значит, что морфологический сдвиг к растительноядности, проиллюстрированный здесь, также мог быть связан с событиями, произошедшими после их таксономического подъёма.
Как постулировано Уилсоном, постепенный сдвиг к растительноядности, на который указывает рост сложности зубов, мог начаться из-за роста густоты прожилок в листве ангиоспермов и их экологического разнообразия в конце позднего мела. Гондванатерии также дополнительно поддерживают гипотезу, что млекопитающие до конца мела не становились значительно более растительноядными. Эти рода имеют гипсодонтные моляры, указатель растительноядности у современных млекопитающих. Гондванатериев не было в хронике ископаемых до конца позднего мела (рис.1), периода, когда покрытосеменные распространялись экологически. Интересно, что челюсти ранних многобугорчатых и современных грызунов показывают не очень сильное пересечение морфопространств, несмотря на значительные морфологические сходства (т.e. на их челюстях есть диастема, небольшой короноидный вырост, и выдающиеся резцы). Многобугорчатые имели направленное назад движение нижней челюсти в момент смычки, а челюсти грызунов двигаются вперёд. Место крепления m. masseter superficialis у последних находится дальше назад на нижней челюсти по сравнению с многобугорчатыми.
Угловой вырост грызунов, таким образом, гораздо более выражен и сильнее отделён от мыщелкового отростка. Расстояние между этими отростками, видимо, является главным отличительным признаком PC1, что может объяснить, почему многобугорчатые не могли в большей степени приблизиться к морфопространствам современных растительноядных. Дальнейшие исследования функциональной анатомии мезозойских млекопитающих и их современных аналогов наверняка поможет провести лучшие анализы морфологических схем в этой работе. Сокращение разнообразия не-многобугорчатых в середине мела (рис.4) случилось из-за исчезновения млекопитающих с правой стороны морфопространств насекомоядных и хищных/всеядных на рис.3. Многие рода, занимавшие это морфопространство, были эутриконодонтами и симметродонтами. У этих таксонов бвли слабо развитые угловые отростки, и большие короноидные (см. рис.2), черты, что говорят об иной функции жевания, чем те, что сохранились. То есть, среднемеловый упадок разнообразия не-многобугорчатых как будто отражает утерю доминировавших прежде, в до-покрытосеменных фаунах млекопитающих, челюстных морфологий. Не-многобугорчатые начала позднего мела, по-видимому, были исключительно мелкими насекомоядными, как видно из их положений по оси PC1 (рис.3), низкий уровень морфологической вариативности челюстей и зубов (рис.3–5), и малый размер тела, судя по размерам челюстей и зубов (рис.5; дополнительные материалы), так как последнее есть очевидное физическое препятствие против хищничества или всеядности. Многие крупные плотоядные млекопитающие мезозоя были эутриконодонтами. Их практическое вымирание в середине мела помогает объяснить отсутствие хищников и всеядных.
Можно было ожидать, что другие таксоны заменят эутриконодонтов и займут опустевшие ниши хищников. Но этого не случилось. Относительно крупные мезозойские млекопитающие не возникали вновь до конца позднего меля (рис.5), когда появились метатерии, такие, как Didelphodon и Eodelphis. Челюсти этих крупных метатериев занимают морфопространства, схожие с таковыми крупных раннемеловых видов Repenomamus (известный хищник), Gobiconodon и Vincelestes, что свидетельствует о том, что они вышли за пределы только насекомоядной диеты. Несмотря на увеличение размеров тела и предполагаемый рост разнообразия диет в конце позднего мела, в этой временной ячейке и у многобугорчатых, и всех прочих сравнительно низкая морфологическая вариативность (рис.4). Но уровень морфологического разнообразия всех млекопитающих в это время значительно возрастает. Изучение рис.3 даёт такое объяснение этим противоречивым результатам: многобугорчатые отклонились от остальных к растительноядности, чем повысили общее разнообразие млекопитающих, при этом сохранив низкое внутригрупповое разнообразие.
(c)Почему терии выжили и распространились?
Хотя терии выглядят морфологически ограниченными во время распространения покрытосеменных, они, кроме цимолодонтов, оказались единственной группой млекопитающих, имевшей в мелу значительный таксономический подъём (рис.1). Наличие трибосфенических моляров — одна из тех их черт, что могла дать им преимущество перед соперниками. Луо говорит, что эти моляры, позволявшие по-новому разгрызать и прокусывать пищу в процессе жевания, были более эффективны для мясоедов или всеядных. Трибосфенические моляры могли быть идеальны для поедания насекомых, некоторые отряды которых, похоже, коэволюционировали и расселялись с покрытосеменными, как опылители. По Кемпу, во время каждой большой смены в фаунах синапсид (т.e. от пеликозавров к терапсидам, от терапсид к цинодонтам и от цинодонтов к млекопитающим), растительноядные и крупные хищники не замещали других растительноядных и крупных хищников. Вместо того, мелкие хищники преуспевали в выживании и занимали заново экологические ниши. Такой сценарий можно применить к не-многобугорчатым млекопитающим в МНР, которые исходно как будто были маленькими насекомоядными плотоядными. Кемп отмечает, что огромные энергозатраты у маленьких млекопитающих могут вызывать больше соперничества за источники пищи, что также может подтолкнуть эволюционные изменения через естественный отбор. Если изменения экосистем в МНР действовали как давление на млекопитающих в целом, то гипотеза Кемпа может объяснить, как мелкие насекомоядные терии возникли в тот период и наиболее успешно распространились. Другая гипотеза успеха териев касается их способа передвижения. Свидетельства говорят, что многие ранние терии были древесными и лазающими, и Шеттак и Уильямс утверждают, что древесность повышает у млекопитающих продолжительность жизни. Госвами предполагает, что древесный образ жизни может иметь сильное давление отбора ввиду избегания хищников или освоения новых тропических ниш, выводя, что возросшая способность к хватанию и гибкость, требуемая от древесных созданий могла сыграть ключевую роль в будущем распространении териев.
Заключение
Сокращение численности млекопитающих с триконодонтными, симметродонтными, докодонтными, плагиаулакоидными, и эупантотериевыми функциональными типами зубов в сочетании с одновременным распространением трибосфенических териев и цимолодонтных многобугорчатых предполагает, что рода последнего типа были более успешны в борьбе с меняющейся экологией мелового периода. Сокращение числа функциональных типов зубов также указывает на общее уменьшение морфологического разнообразия моляров. Результаты геометрического морфометрического анализа челюстей, дополненные измерениями зубов и челюстей не-многобугорчатых как приближением для размера тела, указывает на сдвиг к насекомоядности у не-многобугорчатых в начале позднего мела, и сдвиг к растительноядности у цимолодонтных многобугорчатых в позднем мелу. Логично заключить, что меловые млекопитающие, которые дополняли свои диеты растительным материалом, выиграли от расширения таких источников пищи (напр., фрукты и семена), а таксоны, для которых основным источником пищи были насекомые, выиграли от коэволюции с опылителями покрытосеменных. Но история может быть более запутанной. Многобугорчатые могли на самом деле не склоняться к растительноядности до конца позднего мела, времени, когда покрытосеменные играли большие экологические роли. Не-многобугорчатые, то есть к позднему мелу преимущественно терии, показывают уменьшение морфологического разнообразия и размера тела во время распространения покрытосеменных. Возможное объяснение таксономической радиации териев и одновременно их морфологического упадка может быть в том, что клада обладала чертами (напр., небольшой размер, насекомоядная диета и перемещение по деревьям) были идеальны для выживания при экологических переменах, принесённых МНР и распространением покрытосеменных. Эти соображения указывают, что перемены, случившиеся во время их распространение были чувствительны для млекопитающих и привели ко временному сокращению разнообразия последних.
Дополнительные данные
Функциональные типы зубов
Для наших целей, «триконодонтными» здесь называются моляры с тремя главными буграми, расположенными линейно или тупоугольно, как у эутриконодонтов, амфилестид, и морганукодонтов. «Симметродонтные» моляры — с тремя главными буграми, расположенными треугольно, и без талонида. «Трибосфенические» моляры имеют двухфункциональные тригониды и талониды, как у истинных трибосфеников эвтериев, метатериев, и стем-бореосфенид, или псевдо-трибосфеников австралосфенид. Верхние моляры плагиаулакоидных многобугорчатых имеют два ряда бугров, а цимолодонтных три, хотя эти дентальные типы отличаются и количеством резцов и премоляров. См. Келан-Яворовскую за описанием гондванатериев, харамиид, эупантотериев, и докодонтов. Один из самых спорных примеров функциональных типов зубов — это триконодонты (в самом широком смысле термина). Стоит заметить, однако, что даже если в рода триконодонтов выбрать другие функциональные типы (напр., если включить амфилестид в симметродонтов, а не триконодонтов), общее разнообразие млекопитающих-трибосфеников и многобугорчатых по сравнению с прочими типами в мелу (главный фокус этого раздела исследования) не изменится (рис. 1).
Изображения нижней челюсти: современные виды
Были изучены нижние челюсти 39 видов млекопитающих, принадлежащих 32 семействам и 18 отрядам. При выборе современных видов для исследования в учёт принимались три признака: 1) тип диеты, 2) сходство с мезозойскими млекопитающими, и 3) таксономическое разнообразие. Так как целью работы было использовать информацию о диетах нынешних видов как аналог для вымерших видов, важно включить весь спектр современных типов диет, и поэтому некоторые виды участвовали только лишь из-за своих специфичных диет (напр., травоядная капибара). Некоторые виды выбраны на основании общего экоморфологического сходства с мезозойскими млекопитающими. Например, многие из них (особенно многобугорчатые) выглядят похожими на грызунов (напр., на их челюстях есть диастема, маленький короноидный вырост, и выступающие резцы); и было выбрано 11 видов отряда Rodentia. Другие ранние млекопитающие как будто похожие на землероек насекомоядные (т.е., у них острые бугристые зубы и выраженный короноидный вырост), и для них было включено два вида землероек и много иных мелких насекомоядных (всего 15 видов). Келан-Яворовска описывает эутриконодонтов, среди которых был достоверный хищник репеномам, как плотоядных и относительно крупных, скорее на мясной диете, а не только насекомых. Тогда в работу включили некоторых ныне живущих мелких хищников.
Длины челюстей у современных насекомоядных и хищников/всеядных
Разница между насекомоядностью и хищничеством у млекопитающих больше связана с размером тела, а не специализацией зубов или челюстей. Чтобы помочь различить насекомоядные и хищные морфотипы, мы проверили различия в размере челюсти у современных млекопитающих, принадлежащих к этим типам диет. Источники этих данных те же, что и для сведений о диете. Длина челюсти выбрана как приближение к размеру тела, потому что у вымерших млекопитающих для этого приближения и меры разброса в этом исследовании используются челюсти, так что их результаты более сравнимы с результатами вымерших. Средняя длина челюсти современных насекомоядных (n = 15) равна 39.7 мм, и средняя длина челюсти современных хищников/плотоядных всеядных (n = 10) равна 82.9 мм. Логарифмические значения длин челюстей использовались для статистического сравнения двух типов диет. Двусторонний тест логарифмических значений их длин демонстрируют, что средняя длина челюсти насекомоядного намного короче, чем хищника/всеядного (t = 3.36, p = 0.003). Так как нормальное распределение данных не может быть подтверждено, длины этих типов были также сравнены с помощью критериев Уилкоксона и Манна-Уитни, итоги которых подтвердили значительную разницу между двумя образцами (U = 19, p < 0.05). Эти результаты поддерживают утверждение, что рост средней длины челюсти млекопитающего (т.e., размера тела) отражает рост вероятности хищности у вида.
Геометрические морфометрические точки
Для нижних челюстей было выбрано семь опорных точек. Огромная вариативность форм челюстей ограничила их теми, что универсальны практически для всех млекопитающих. Единственное исключение — угловой вырост (точка 7), который был утрачен у многих мезозойских видов (напр., большинства многобугорчатых и «симметродонтов»). Когда угловой вырост отсутствует или не выражен, используется самая постеровентральная точка ветви челюсти. Эти точки предназначены, чтобы найти три главные характеристики челюсти: общую длину и глубину, область крепления височной мышцы, и область крепления крыловидных/жевательных мышц. Большая височная мышца указывает на хищную диету, и эта мышца крепится к короноидному отростку, вершина которого точка 4. Следовательно, ожидаемо, что короноидный отросток крупнее всего (т.e., точка 4 дальше всех от точек 2 и 5) у хищников. Растительноядные с большей вероятностью имеют мощные крыловидные и жевательные мышцы, идеальные для размалывающих движений челюсти, то есть предполагаем более выраженный угловой вырост и меньший короноидный. Главное различие между челюстями хищников и растительноядных, видимо, расстояние между мыщелковым выростом (точка 6) и угловым (точка 7). Результаты этого исследования говорят, что для современных видов это верно (рис. 2). Описания опорных точек даны здесь:
- Самая антеродорсальная вершина тела нижней челюсти. (Зубы не считаются).
- Точка, где основание короноидного выроста касается горизонтального тела челюсти.
- Прямая между точкой 1 и точкой 2. Ещё одна прямая проведена перпендикулярно ей. Точка 3 определена как место, где эта линия касается вентральной стороны челюсти.
- Самая дорсальная точка короноидного отростка.
- Самая антероаентральная точка промежутка между короноидным и мыщелковым отростками.
- Самая постеродорсальная точка мыщелкового отростка.
- Самая постеровентральная точка углового отростка. Если его постеровентральная сторона широка, то берётся её центральная точка. Если у вида нет хорошо развитого углового отростка, то используется самая постеровентральная точка ветви челюсти.
Длины челюстей
Геометрические морфометрические техники, как выполнены в этой работе, ценны для анализа морфологии, но не учитывают размер образцов. Средний размер не-многобугорчатых требовался как средство отличия насекомоядных от возможных хищников. Таким образом, длины зубов и челюстей были использованы как приближение для размера тела. Эта информация позволила создать вспомогательные кривые разнообразия для не-многобугорчатых (рис. 5). Изображения челюстей для определения длин взяты из тех же источников, что и для геометрической морфометрики, и отдельные дополнительные фрагменты у Келан-Яворовской. Длина нижней челюсти была измерена от самого дальнего конца мыщелкового отростка до самой передней точки горизонтального тела челюсти с использованием изображения и линейной шкалы. Все челюсти не-многобугорчатых, использованные в геометрическом морфометрическом анализе, были измерены. Чтобы увеличить количество образцов во временных ячейках, было оценено 25 дополнительных длин челюсти с использованием фрагментов челюстей, всего 88 пунктов. Так как 12 родов жило больше чем в одной временной ячейке, набор данных включает всего 100 пунктов на шкале времени. Фрагменты, использованные для оценок — те, где имеются зубы и для которых есть целые челюсти таксономически близко связанных видов, что позволяет провести сравнение размеров (через измерение зубов) и форму челюсти (через оценку положения недостающих опорных точек).
Длины зубов
Хотя длина зубов может значительно варьировать относительно размеров тела, и это не такой точный его индикатор, как длина челюсти, нам чаще доступны окаменелости зубов, чем челюстей, и они дают лучшее временное разрешение результатов. Первичная литература была исследована нами на предмет изображений челюстей с целыми первыми нижними молярами (m1). Поскольку нижние моляры одного вида могут значительно варьировать по форме и размеру, учитывались только подтверждённы первые нижние моляры. В попытке улучшить количество образцов в более древних временных ячейках длины нижних моляров голотопов были включены в девять ранних линий и видов докодонтов из позднего триаса и юры. Антеропостериоральные длины m1 были получены либо из измерений, представленных в литературе, либо вычислены с помощью изображения и линейной шкалы. Измерения проведены на уровне рода. Если открыто много образцов одного вида или много видов одного рода, использовалась мера крупнейшего, чтобы минимизировать риск взятия длины от ювенильной особи. Измерения из мелового периода разделены на меньшие временные ячейки (см. рис. 5) для лучшего анализа эффекта распространения покрытосеменных. Всего отмечено 135 длин m1. 29 родов простираются таксономически в несколько временных ячеек, и увеличивают набор данных до 164 пунктов, учтённых на временной шкале.
Разнообразие мезозойских млекопитающих. Автор — KakureRyoshiX.
Источник ВКонтакте
Оригинал в Proc. Royal Soc. B., Oktober 2016.
Рекомендуем прочесть
 Эволюция на двух уровнях: о генах и формах
Эволюция на двух уровнях: о генах и формах Декоммунизация и антиваксеры
Декоммунизация и антиваксеры![[Наступление деникинцев] В Ингушетии](http://www.socialcompas.com/wp-content/uploads/2021/12/ADj4FkHXUEM-150x150.jpg) [Наступление деникинцев] В Ингушетии
[Наступление деникинцев] В Ингушетии «Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве
«Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве Изображая «монархическую» жертву
Изображая «монархическую» жертву Полиция разогнала палаточный городок сторонников референдума перед представительством правительства
Полиция разогнала палаточный городок сторонников референдума перед представительством правительства  Когда мёртвый хватает живых
Когда мёртвый хватает живых «Блэз Компаоре — мёртв, Томас Санкара — жив!»: образы, нарративы и дискурсы народного восстания в Буркина-Фасо (2014-2015 гг.)
«Блэз Компаоре — мёртв, Томас Санкара — жив!»: образы, нарративы и дискурсы народного восстания в Буркина-Фасо (2014-2015 гг.)
