
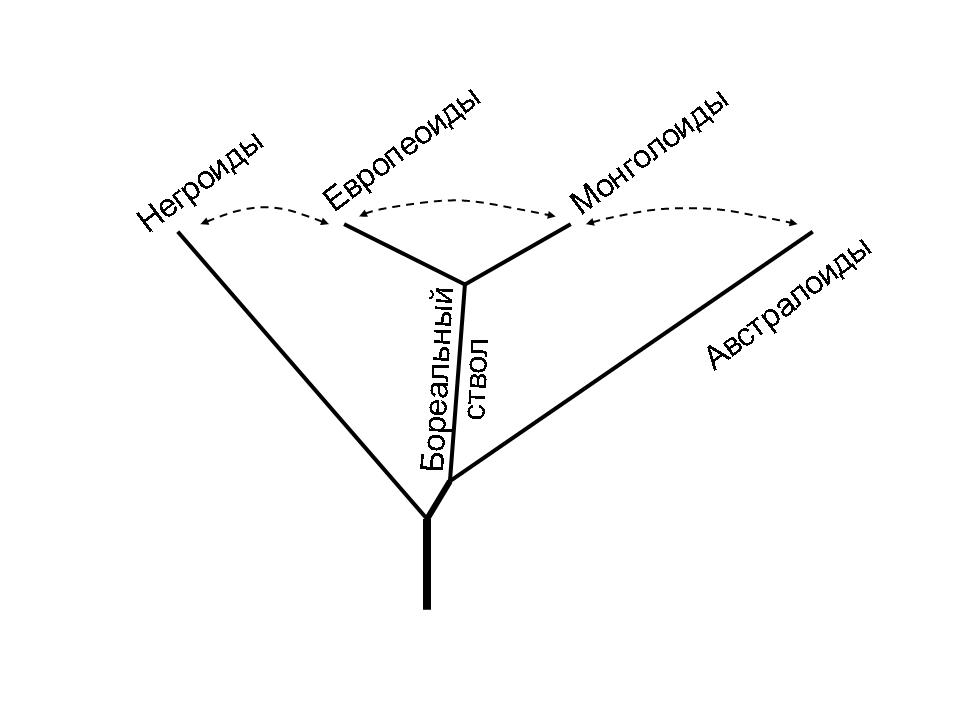
Схема генеалогических отношений основных человеческих рас. Древнейшие африканские линии не показаны. Пунктиром обозначены главные направления метисации. Из: А.Г.Козинцев. Европеоиды, монголоиды, австралоиды: стадиальность или метисация?
М.О. Вергелес
В статье в исторической перспективе рассматриваются различные подходы к пониманию биологической изменчивости человечества в американской физической антропологии. Промежуточным итогом многолетних дебатов в 2007 г. стал симпозиум «Согласование понятия «раса»: как биологические антропологи видят изменчивость человечества», на котором все ученые согласились с существованием различий между популяциями, а единственным фундаментальным разногласием оказался взгляд различных специалистов на характер и причины географической изменчивости человека [Edgar, Hunley, 2009]. Понятие «раса» в США, по мнению многих ученых, включает в себя представление о дискретности расовых вариантов, их внутренней единообразности и более того, существование определенной иерархичности среди больших рас [Lieberman, Kirk, 2004]. Это не согласуется с определением расы как популяции с общим генофондом и историей, которое существует в российской антропологии [Проблема расы… 2002]. Таким образом, понятие расы в российской науке более соответствует американскому термину «происхождение» (ancestry), нежели «раса» (race). Первые этапы изучения изменчивости человечества происходили в рамках типологического подхода, который оставался широко распространенным до середины XX в. Ключевую роль в смене парадигмы сыграли генетические исследования, в частности, исследования Ричарда Левонтина. Этот новый подход предполагал отрицание расовых категорий ввиду того, что большая часть изменчивости приходится на внутригрупповой, а не на межгрупповой уровень [Lewontin, 1972].
Подходы генетиков к изучению изменчивости не раз подвергались критике. Например, отмечается, что Левонтин рассматривал каждый генетический маркер по отдельности, не делая поправки на возможную корреляцию признаков между собой и соответственно на зависимость их распределения в группах [Edwards, 2003]. На настоящем этапе развития науки изучаются особенности генетического разнообразия человечества, специфика распределений различных признаков, а также строятся различные модели, способные объяснить возникновение именно такого рисунка изменчивости. Наиболее точно реальность отражают сложные модели, сочетающие в себе процессы разделения популяций, миграции, эффекты «бутылочного горлышка», а также постоянные взаимосвязи и обмен генами между популяциями. Широко исследуется раса как социальный конструкт и влияние расовой дискриминации на показатели здоровья. Неоднократно была показана возможность разделения современного человечества на популяции, используя генетические, краниометрические и другие данные. Таким образом, на данный момент большинством ученых признается существование отдельных групп в составе человечества, которые, однако, имеют высокий уровень перекрытия изменчивости по многим признакам. Исследования рас и этносов находят свое применение в медицине, криминалистике, палеоантропологии, фармакологии и других отраслях знания. Дальнейшие исследования биологической изменчивости человечества представляют широкий научный и практический интерес.
Введение
Содержание
Концепция расы, согласно исследованиям историков, существует только с XVI в. При этом активное развитие и распространение этих идей приходится на следующий, XVII век [Lieberman, 1968]. Изначально деление на расы основывалось на нескольких морфологических признаках и на географическом ареале их распространения. Продолжительный колониальный период придал термину «раса» политическую и культурную окраску. В этот же период появились представления о неравноценности отдельных рас по отношению друг к другу. Это обстоятельство привело к еще большей политизированности проблемы рас, которая стала скорее социальной проблемой, нежели антропологической. Для стран, в которых население состоит не только из разных этносов, но и из разных расовых вариантов, вопрос об их происхождении и их равенстве или неравенстве всегда был и остается важным. Наиболее болезненно это проявлялось на американском континенте и, в первую очередь, в США. Данная работа посвящена основным особенностям представлений о расе как о биологическом и социальном конструкте в американской физической антропологии с середины XX в. до настоящего времени. Также обсуждаются сходства и различия в использовании термина «раса» в американской и отечественной традициях.
Определение понятия «раса»
На волне вновь возникшего в XXI в. широкого интереса к проблемам расы в мае 2007 г. в Университете Нью-Мехико состоялся симпозиум «Согласование понятия «раса»: как биологические антропологи видят изменчивость человечества» (Race reconciled? How biological anthropologists view human variation).
В результате обсуждения все участники конференции согласились со следующими положениями:
- существует значительная внутрипопуляционная (индивидуальная) изменчивость; также существует межпопуляционная изменчивость и различия между группами популяций;
- форма внутри- и межгрупповой изменчивости в значительной мере сформирована культурой, языком, экологией и географией; расы не являются точным или продуктивным способом описания биологической изменчивости человека;
- изучение изменчивости человечества важно для медицины, судебных наук и культуры в целом;
По сути, единственным фундаментальным разногласием в рамках симпозиума оказался взгляд различных специалистов на характер и причины географической изменчивости человека [Edgar, Hunley, 2009, р. 2].
В научной литературе существует множество определений понятия раса, вот лишь некоторые из них:
«Расы суть популяции, отличающиеся по частоте некоторых генов и обменивающиеся либо потенциально способные обмениваться генами через барьеры (обычно географические), их разделяющие» [Dobzhansky, 1944]. «Раса есть совокупность людей, обладающих общностью физического типа, происхождение которого связано с определенным ареалом» [Рогинский, Левин, 1978].
«Раса — это большая популяция индивидов, отличающаяся от других рас общим генофондом» [Фогель. Мотульски, 1986, цит. по: Проблема расы… 2002, с. 13].
Общим для всех определений является то, что раса — это популяция с общим генетическим фондом и/или фенотипом, особенности которой статистически отличаются от других популяций, что является результатом общей территории и предка в прошлом. В настоящее время такое понимание расы характерно, скорее, для отечественной антропологии.
В США к термину «раса» относятся несколько пренебрежительно и с недоверием. В американской антропологии, согласно L. Lieberman и R.C. Kirk, в определение расы также входят следующие понятия[1]:
- расы представляют собой дискретные группы индивидов, между которыми есть четкие биологические границы; внутри рас изменчивость мала, популяции являются гомогенными; культура каждой группы определяется ее биологией;
- уровень развития цивилизаций иерархичен и связан с представлениями о европейцах как о высшей расе, «азиатах» как вторых в цивилизационной иерархии, и негроидах как о наименее цивилизованных [Lieberman, Kirk, 2004, р. 138].
С.С.Gravlee определяет расу как культурно обусловленный систематический способ рассмотрения, восприятия и интерпретации реальности. По его словам в Северной Америке центральным принципом понимания концепции рас является представление о расах как о естественном разделении человечества на подвиды, которые оказываются дискретными, неизменными и относительно гомогенными. Также к такому классически американскому пониманию расы относится и представление о том. что признаки, на основе которых разделение рас возможно, определяют и отражают некоторые фундаментальные внутренние биологические различия [Gravlee, 2009, р. 48].
При этом происходит некая подмена понятий, предлагается увидеть огромное различие и несовместимость двух терминов-«расы» и «происхождения» (в оригинале — ancestry). «Раса» в понимании многих американских ученых — социально обусловленный механизм самоидентификации и идентификации принадлежности к группе и биологически не несущий смысла концепт, в то время как «происхождение» — научно обоснованный подход для описания биологического компонента популяционной изменчивости [Konigsberg et al., 2009, р. 77-78].
Также существует бытовое (folk) понятие расы, которое частично основывается на биологическом понимании, но в большей мере примешивает этническую, культурную и национальную составляющую. [В том числе люди в соответствующем обществе при определении расы другого человека смотрят не только на цвет кожи, форму носа и губ и т.д. морфологию, но также оценивают, как одет, как держится и другие сигналы классовой принадлежности]. Например, в США с практическими целями существует законодательный акт о минимальном разделении человечества на расы при необходимости их классификации: американский индеец или эскимос, монголоид, негроид или афроамериканец, коренной житель Гавайских и других тихоокеанских островов, европеоид. Также отдельно отмечается этническая принадлежность — испанец/латино или не испанец/латино [Revisions… Электронный ресурс]. Такое разделение используют в медицинских исследованиях, и оно оказывает большое влияние на выбор брачного партнера [Edgar, 2009].
Для сравнения приведем определение расы в современной российской физической антропологии [Проблема расы… 2002, с.14]:
- раса есть популяция;
- признаки, отличающие расы друг от друга имеют наследственный характер;
- раса определяется как открытая популяция, репродуктивно в той или иной мере изолированная от других групп (рас) и потенциально способная к смешиванию с ними;
- все расы принадлежат к одному виду;
- расы имеют свой географический ареал или единый ареал происхождения.
Понятно, что понятие расы в российской науке скорее соответствует американскому термину «происхождение» (ancestry), нежели «раса» (race). Дело в том, что развитие американской и российской антропологии происходило различными путями. В американской антропологии главенствующая роль всегда отводилась изучению культуры, и человек соответственно рассматривался как существо двойственное — с одной стороны, культурное, с другой — биологическое. То есть антропология в американском смысле —
«это комплексная наука, включающая в себя изучение разных аспектов культуры народов, поэтому американские антропологи одновременно занимались этнографией, фольклористикой, лингвистикой и археологией» [Подзюбан, 2014, с. 329].
В российской традиции физическая антропология всегда существовала как относительно самостоятельная независимая дисциплина, рассматривавшая именно биологию человека.
Тем не менее, в дальнейшем речь пойдет именно о таком американском понимании расы как о социальном конструкте, имеющем долгую историю и негативную окраску.
Против использования расовых классификаций в американской научной литературе выдвигалось и выдвигается три «классических» аргумента [Gravlee, 2009, р. 50]:
- тот факт, что большая часть изменчивости признаков у Н. sapiens клинальна, то есть изменения признака происходят плавно, резкие границы между популяциями отсутствуют [Livingstone, 1962];
- отсутствие у генетической изменчивости конкордантного характера (nonconcordant), т.е. признаки, используемые для выделения рас, не имеют значения для предсказания других аспектов биологии;
- лишь малая часть генетического разнообразия приходится на межрасовую изменчивость.
Подходы к изучению изменчивости человечества в XX веке
В XVII в. в результате великих географических открытий перед европейцами предстал мир, заселенный огромным многообразием различных людей, имеющих различные обычаи, верования и культуры. Концепция расы в какой-то мере появилась в ответ на это, как попытка принять и понять новую информацию. Слово «раса» начинает широко использоваться как в научной литературе, так и в повседневном общении. В научных кругах эта концепция на протяжении истории вызывала немало дебатов. Так, еще в XVII и XVIII вв. спорили о том, имеют ли все расы единое происхождение. В XIX в. этот спор сменился на спор о равенстве или неравенстве представителей различных расовых типов. В первые десятилетия XX в. антропологи, социологи и психологи пришли к выводу о несостоятельности любых концепций неравенства рас и под их влиянием большинство ученых и образованных людей сменили свои взгляды с расизма на эгалитаризм [тут надо уточнить – лишь после мая 1945-го; до этого расизм был легитимен для образованной публики и западных стран, и колоний; антирасистами были лишь подозрительные и радикальные элементы, леваки или, храни Аллах, коммунисты, вроде авторов «Манифеста генетиков»].
Активно участвуя в доказательствах равенства всех современных людей и в пропаганде этих идей, многие антропологи пошли еще дальше и начали полностью отрицать существование рас как таковых. Две крайние точки зрения представлены, с одной стороны, учеными, отстаивающими факт существование рас, и с другой стороны, учеными, утверждающими, что раса есть миф, который должен быть окончательно уничтожен. Этот спор продолжается уже более четырех десятилетий [Lieberman, 1968].
До начала 1960-х гг. в США существовало некое разделение сфер интересов между антропологами и социологами: большинство антропологов занималось изучение примитивных обществ на окраинах цивилизованного мира, в то время как социологи имели дело с проблемами социума, в том числе и с взаимоотношениями рас в индустриальных городах капиталистического мира. Те же антропологи, кто интересовался проблемами рас (Н. Powdermaker, A. Davis, St.C. Drake, Е. Leacock) публиковались либо как члены отделений социологии или образования, либо же вообще занимались этими вопросами вне своей академической карьеры [Baker, Patterson, 1994].
F.B. Livingstone был одним из первых антропологов, яростно пропагандировавших отказ от использования понятия расы при изучении разнообразия человечества. В переписке с Т. Dobzhansky [Феодосий Добжанский, один из создателей синтетической теории эволюции] в 1962 г. на страницах журнала «Current Anthropology» он настаивал на том, что если вид и может быть разделен на подвиды (расы) на основании некого одного признака, то, в это же время, использование множества признаков будет давать разнонаправленные и противоречащие друг друга разделения. В результате, по его словам, после того, как учтены более 3-4 различных признаков, уже не будет деления большего, чем популяция [Livingstone. 1962].
Десятью годами позднее R.C.Lewontin [Ричард Левонтин, ученик Добжанского, классик эволюционной и популяционной генетики, один из наиболее мощных критиков панадаптационизма в теоретической биологии], эволюционный биолог из Чикагского университета, впервые применил метод измерения генетической вариабельности к вопросу о существовании рас. Он исследовал 16 белков групп крови, сывороточные белки и ферменты красных клеток крови. В результате обнаружилось, что по отношению к полной изменчивости для человека как вида внутригрупповая изменчивость составляет 85.4%, изменчивость популяций внутри одной расы — 8.3% и межрасовая изменчивость — всего 6.3%. То есть, фактически, расы не имеют генетической или таксономической значимости [Lewontin, 1972]. В работе использовалось два уровня популяционных структур: уровень больших рас (их было выделено 7: европеоидная, африканская, монголоидная, южноазиатские аборигены, коренные американцы, тихоокеанская, коренные австралийцы) и уровень отдельных популяций (китайцы, навахо, португальцы и т.д.).
При рассмотрении подобных работ по генетике всегда возникают вопросы по поводу использованных классификационных схем. Так, у R.C. Lewontin к монголоидной расе относятся, например, айны, которые в российской антропологии выделяются в самостоятельную группу. Также в качестве примера сомнительного использования накопленных об изменчивости человечества данных при разделении человечества на группы можно привести работу N. Yu из Чикагского университета и его коллег из разных стран мира [Yu et al., 2002]. Ими было проведено исследование SNP (single nucleotide polymorphism, однонуклеотидный полиморфизм) в 50 некодирующих участках ДНК. При этом выборка не только была крайне малочислена (всего 30 человек при попытке доказать отсутствие различий на уровне вида!), но и объединяла индивидов, не относящиеся к одной группе ни в одной из существующих расовых классификаций. 10 африканцев составили 2 пигмея, ганец, представитель бантуязычного народа кикийю, бушмен племени !kung, представитель народа luo, 2 нигерийца, 1 зулус, 1 бантуязычный южноафриканец. 10 европейцев были представлены финном, французом, немцем, венгром, итальянцем, португальцем, русским, испанцем, шведом и украинцем. И, наконец, 10 азиатов: индийцы — пенджабец и бенгалец, камбоджиец, 2 китайца, японец, монгол, вьетнамец, якут и тайванец. Полученные нуклеотидные различия показывают, что различия внутри африканской группы больше, чем между африканцами и европейцами, из чего делается вывод об отсутствии рас как таковых. Результаты абсолютно неудивительны, ведь бушменов зачастую выделяют чуть ли не в отдельную большую расу [Хрисанфова, Перевозчиков, 2005, с. 271], а индийцы во всех существующих классификациях относятся к европеоидам.[Характерно, что работа Левонтина свободна от такого рода изъянов и цитируется до сих пор; эту же автор амальгамирует с ней, только затем чтобы выразить неприятие его подхода].
Тем не менее, данные схожие с результатами R.C. Lewontin на протяжении последних десятилетий были получены учеными неоднократно (табл. 1). Интересно, что учеными из раза в раз обнаруживается наличие статистически достоверных различий между группами людей, которые они, однако, признают слишком слабыми для таксономического уровня.Таким образом, в американской антропологии. начиная с 1960-х гг., происходит изменение господствующего подхода к пониманию изменчивости человечества. Отрицается типологический подход, подразумевающий строгое и однозначное разделение всех людей на отдельные расы. Более широко распространенным становится взгляд, основанный на новейших достижениях генетики. Такой взгляд предполагает отрицание расовых категорий ввиду того, что большая часть изменчивости приходится на внутригрупповой, а не на межгрупповой уровень.
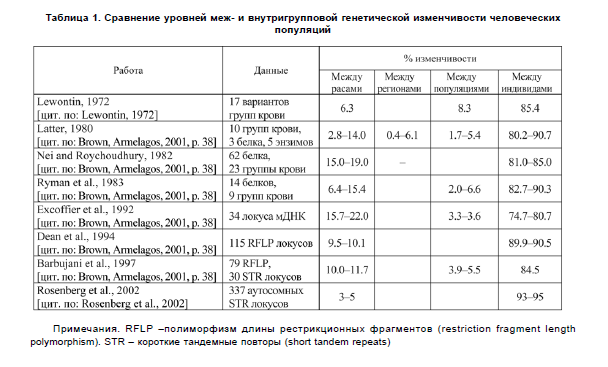 Однако у этого подхода, как и любого другого, есть свои слабые стороны, вызывающие вопросы и споры:
Однако у этого подхода, как и любого другого, есть свои слабые стороны, вызывающие вопросы и споры:
- F.B. Livingstone, по сути, не подкрепил своей позиции никакими эмпирическими доказательствами. В переписке 1962 года Т. Dobzhanskyсогласился с F.B. Livingstoneв том, что дискордантные распределения частот генов часто встречаются в человеческих популяциях. Однако, по мнению Т. Dobzhansky, физические и социальные преграды для свободного обмена генами между популяциями регулярно создают разрывы в непрерывной клинальности и, напротив, конкордантность в распределении частот генов в группах людей. Также вероятно, что противоположно направленные клины, наблюдаемые F.B. Livingstone, могли сформироваться в результате чисто филетического процесса через случайную потерю особенностей и признаков.
- Учитывая, что лишь 6.3% изменчивости нейтральных генетических локусов приходится на межрасовую изменчивость, кажется справедливым утверждение об отсутствии у подобного деления таксономического значения. Однако J.C. Long, R.A. Kittles[Long, Kittles, 2003] показали, что добавление данных о геноме шимпанзе в выборку, представляющую пять основных географических регионов, лишь незначительно увеличивает долю межгрупповой изменчивости в общей изменчивости выборки. Для выборки только из современных людей общая межгрупповая изменчивость составляет 11.9%, для выборки, включающей еще и шимпанзе — 18.3%. Данный факт ставит под сомнение ту генетическую и таксономическую интерпретацию низких значений изменчивости, которую дают R.C. Lewontinи другие генетики [Hunleyetal., 2009, р. 35-36]. [Это верно только в том случае, если шимпанзе относить к роду Homo, с чем легко согласятся генетики, но вряд ли морфологи].
Подход, использованный R.C. Lewontin, как таковой, имеет ряд недостатков, что неоднократно обсуждалось в научной литературе. Так, например, отмечается, что он рассматривал каждый генетический маркер по отдельности, не делая поправки на возможную корреляцию признаков между собой и соответственно на зависимость их распределения в группах. Генетик A.W.F. Edwards из Кембриджского университета [Edwards, 2003] утверждает, что данные R.C. Lewontin верны для уровня отдельных единичных локусов, которые действительно будут давать большой уровень перекрытия между группами. Анализ же множества локусов даст более реалистичную картину с меньшим перекрытием, так как важна не только изменчивость отдельных локусов, но и корреляция между признаками, дающая определенный паттерн общей изменчивости.
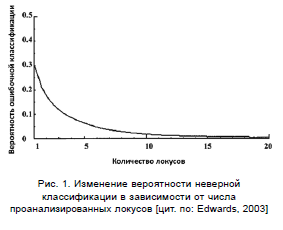
При рассмотрении множества локусов, индивиды группируются сообразно со своим географическим происхождением. На рис. 1 представлена зависимость вероятности ошибочного отнесения к определенной группе (ось Y) от количества рассматриваемых локусов (ось X). Важно, что при этом внутри- и межгрупповая изменчивость не меняется, но точность классификации возрастает. Становится понятным, что при рассмотрении достаточного количества маркеров, разделить выборку возможно даже при высоком уровне индивидуальной изменчивости.
[Однако нельзя объяснять непонятное через неизвестное: а когда было ли выяснено, действительно ли распределение разных генных вариантов коррелирует между собой, ответ скорей был отрицательный. Современные данные показывают следующую диалектику взаимодействия «морфологии» и «генетики». Первая эффективно выделяет расы и расовые подразделения, характеризует их, но не очень пригодна для реконструкции истории их дифференциации. Данные по структурным генам, особенно связанным с адаптацией к местным условиям, не очень успешны для выделения рас и оценки истории, в отличие от полных сиквенсов ДНК, позволяющих отследить т.н. филогенетический сигнал.
В т.ч. на его основе сейчас необходимо вернуться к идее бореального надрасового ствола, объединяющего европеоидов и монголоидов, т.е. к делению человечества на «север vs юг» против прежнего «западный ствол vs восточный». Это оказывается столь же непривычным, сколько плодотворным, т.к. снимает многие противоречия между морфологами, археологами и лингвистами, особенно в отношении уральской расы и других промежуточных групп между европеоидами и монголоидами. См. также книгу ученика Р.Левонтина Ч.Уэллса «Генетическая одиссея человечества». Что отражает общее правило независимости эволюции на двух уровнях, «генов» и «форм»]
Подходы к изучению изменчивости человечества в XXI веке
С развитием генетических методов и развитием науки появляются новые подходы к изучению проблемы рас. Так, в исследованиях, приходящихся уже на начало XXI в., начинаются поиски новых альтернативных способов описания изменчивости современного человечества. Ряд работ показывает наличие географически определенного распределения генетических маркеров, таких как STR, SNP, VNTR (variable number tandem repeat, тандемные повторы с переменным числом звеньев), Alu-повторов [Allocco et al., 2007; Bamshad et al., 2003; Pritchard et а1.^2000; Ramachandran et al., 2005; Rosenberg et al., 2002] несмотря на высокий уровень изменчивости внутри популяции. При использовании кластерного анализа индивидуальные данные группируются в соответствии с происхождением индивидов [Jorde. Wooding, 2004].
 В этих работах показано наличие сильной связи между генетическими и географическими расстояниями. Во многих работах наличие этой связи интерпретируется как подтверждение верности модели «изоляции расстоянием», которая подразумевает отсутствие серьезных разрывов в географическом распределении изменчивости нейтральных для отбора признаков. Географическая отдаленность, основанная на наиболее вероятных путях миграции, по некоторым данным объясняет 78% генетических различий между группами [Ramachandran et al., 2005]. Другие исследования показывают, что расстояние от Восточной Африки (географическое) объясняет уже 82-85% генетических различий [Li et al., 2008; Prugnolle et al., 2005].
В этих работах показано наличие сильной связи между генетическими и географическими расстояниями. Во многих работах наличие этой связи интерпретируется как подтверждение верности модели «изоляции расстоянием», которая подразумевает отсутствие серьезных разрывов в географическом распределении изменчивости нейтральных для отбора признаков. Географическая отдаленность, основанная на наиболее вероятных путях миграции, по некоторым данным объясняет 78% генетических различий между группами [Ramachandran et al., 2005]. Другие исследования показывают, что расстояние от Восточной Африки (географическое) объясняет уже 82-85% генетических различий [Li et al., 2008; Prugnolle et al., 2005].
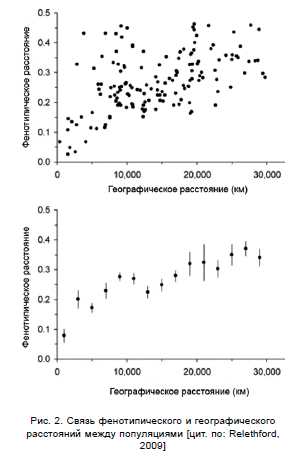
Также высокая степень зависимости от географических расстояний показана для более классических антропологических признаков — краниометрических данных и цвета кожи. Так на рис. 2 показана взаимосвязь фенотипического и географического расстояний для ряда популяций. Для построения графика использовались краниометрические данные, собранные W.W.Howells в 1965-1980 гг. [Howells, 1996]. Были проанализированы 907 мужских и 827 женских черепов из 6 географических регионов: Европа, Субсахара, Австралазия, Полинезия, Северная и Южная Америки, Восточная Азия. Каждый регион представлен тремя популяциями. На верхнем графике изображена связь фенотипического и географического расстояний для каждой из 18 популяций, на нижнем графике точки представляют собой средние фенотипические расстояния для каждых 1000 км географического расстояния вплоть до 20000 км, и для каждых 2000 км далее. Вертикальные линии отмечают стандартное отклонение от среднего значения [Relethford, 2009].
Многие из авторов, соответственно, приходят к выводу о том, что клинальность лучше описывает изменчивость человечества, нежели расы [Hunley et al., 2009].
Н.А. Дубова так критикует подход клиналистов:
«…наличие переходных расовых форм, подтверждая видовое единство современного человека. не отрицает объективного факта существования рас. Отрицать объективность рас на основании большого числа переходных вариантов можно с тем же успехом, с каким кто-то будет утверждать, что не существует ни красного, ни синего, ни желтого цвета, поскольку все они являются составными частями общего спектра, внутри которого не существует четких границ…» [Проблема расы… 2002, с. 34].
S. Ramachandran с коллегами утверждает, что наличие корреляции между генетическими и географическими расстояниями не обязательно должно интерпретироваться как результат действия модели «изоляции расстоянием». Современный паттерн генетической изменчивости человечества может объясняться сложной последовательностью и взаимосвязями между процессами разделения популяций, миграций и изоляций [Ramachandran et al., 2005, р. 15942]. Принципиальное важное различие между моделью «изоляции расстоянием» и сериями разделений в модели S. Ramachandran состоит в том, что модель «изоляции расстоянием» не согласуется с наличием таксономических единиц, в то время как серии разделений вполне допускают их существование.
Схожие результаты получены и K.L. Hunley с соавторами [Hunley et al., 2009]. Главной целью работы был поиск наиболее точной модели для описания специфической изменчивости генетических маркеров у современного человека. Для этого были сопоставлены полученный с помощью различных моделей теоретический рисунок изменчивости и реально наблюдаемое распределение, рассчитанное по нейтральным аутосомным микросателлитным генам 1032 индивидов из 53 популяций. Было обнаружено, что ни одна из существующих моделей не описывает изменчивость достаточно точно. Однако с этим справляется сочетание моделей: серия разделений популяций, эффектов «бутылочного горлышка» и миграций на дальние расстояния, связанных с заселением основных географических регионов. Также важным фактором является наличие обмена генами между соседними популяциями. Сложность подобного процесса не дает возможности выделить отдельные группы популяций так, чтобы они имели одинаковый таксономический уровень, также эта модель не позволяет говорить о наличии независимо эволюционировавших рас.
Е.В. Балановской отмечается, что «если бы расы и генофонд отражали совершенно разные, независимые явления, то различия между расами должны были бы быть равны нулю». Вопрос существования рас, по ее мнению, является вопросом существования популяционного уровня организации материи, он не может быть решен на молекулярно-генетическом уровне [Проблема расы… 2002, с.49].
Несмотря на доминирующее представление об отсутствии рас, существуют некоторые исследования, демонстрирующие, что даже высокий уровень внутригрупповой изменчивости не является препятствием для выделения отдельных групп в выборке. Группой ученых их США и Швейцарии [Novembre et al., 2008] было проведено исследование полиморфизма 500 568 локусов SNP у 3192 европейцев. Пытаясь описывать лишь коренное население каждого региона, когда было возможно, выяснялось место рождения бабушек и дедушек, родителей и самого индивида. Из дальнейшего исследования для проведения максимально возможно точного анализа были исключены индивиды, данные по SNP для которых вызывали сомнения; те, бабушки и дедушки которых были родом из более чем одной популяции и ряд других. В итоговом анализе рассматривался полиморфизм по 197 146 локусам у 1387 индивидов. В результате применения метода главных компонент была получена картина генетической изменчивости в двух измерениях (первых двух главных компонент), которая удивительно точно соответствует географической карте Европы (рис. 3). Полученный график даже отражает подразделения внутри Швейцарии на франко-, германо- и италоговорящих (рис. 4).
 Также возможно разделение индивидов по данным краниометрии. В работе S. Ousley из отдела прикладной криминалистики колледжа Мерсихерст и его коллег было рассмотрены возможности разделения группы из 353 черепов белых и черных американцев, рожденных в XX в. [Ousley et al, 2009). Для каждого индивидуума были доступны данные по полу и возрасту. В качестве сравнительного материала были привлечены уже упоминавшиеся краниометрические данные, собранными W.W. Howells. Была обработана вся имеющаяся база данных — по 2524 индивидам из 28 популяций. Использованные статистические методы — дискриминантный анализ, метод К – ближайших соседей, кластерный анализ, метод главных компонент. Дискриминантный анализ по двум переменным — длине основания черепа (базион-назион) и длине основания лица (базион-простион) — разделяет белых и черных американцев с точностью 80%. Использование пошагового включения переменных для достижения разделения с 95% точностью показало, что для этого необходимо всего 7 переменных — базион-назион (длина основания черепа), базион-простион (длина основания лица), биаурикулярная ширина (аи- аи), ширина носа, ширина неба и высота орбиты. Переменные расположены в порядке их включения в анализ. Такие же результаты получаются и при использовании метода главных компонент и кластерного анализа [Ousley et al., 2009].
Также возможно разделение индивидов по данным краниометрии. В работе S. Ousley из отдела прикладной криминалистики колледжа Мерсихерст и его коллег было рассмотрены возможности разделения группы из 353 черепов белых и черных американцев, рожденных в XX в. [Ousley et al, 2009). Для каждого индивидуума были доступны данные по полу и возрасту. В качестве сравнительного материала были привлечены уже упоминавшиеся краниометрические данные, собранными W.W. Howells. Была обработана вся имеющаяся база данных — по 2524 индивидам из 28 популяций. Использованные статистические методы — дискриминантный анализ, метод К – ближайших соседей, кластерный анализ, метод главных компонент. Дискриминантный анализ по двум переменным — длине основания черепа (базион-назион) и длине основания лица (базион-простион) — разделяет белых и черных американцев с точностью 80%. Использование пошагового включения переменных для достижения разделения с 95% точностью показало, что для этого необходимо всего 7 переменных — базион-назион (длина основания черепа), базион-простион (длина основания лица), биаурикулярная ширина (аи- аи), ширина носа, ширина неба и высота орбиты. Переменные расположены в порядке их включения в анализ. Такие же результаты получаются и при использовании метода главных компонент и кластерного анализа [Ousley et al., 2009].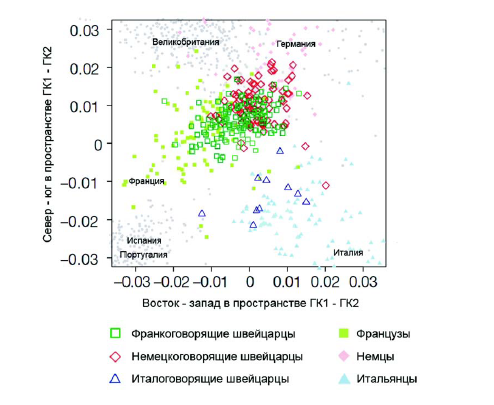
J.H. Relethford использовал дискриминантный анализ для проверки точности отнесения индивидов к определенному географическому региону на основании краниометрических данных. Для этого использовалась краниометрическая база данных W.W. Howells. Были проанализированы измерения 57 признаков у 907 мужских и 827 женских черепов, представляющих 6 регионов Земли. Было обнаружено, что точность отнесения к географическому ареалу составляет 96-97% (табл. 2). Причем, точность классификации не изменяется при разделении или объединении отдельных регионов [Relethford, 2009].
На протяжении последних десятилетий ответ на вопрос о том, существуют ли расы, был однозначно отрицательным. За очень редким исключением с такой позицией соглашались как антропологи, так и генетики. Однако в XXI в. такое положение вещей начало подвергаться сомнению и вопросы о степени и значимости различий между представителями различных рас вновь обретают актуальность. Статьи на эту тему появляются как в научных, так и в популярных изданиях. Важно отметить, что вопрос о реальности существовании рас может быть проинтерпретирован по-разному. Как правило, имеется в виду вопрос о том, существуют ли расы как естественное биологическое деление человечества. Этот вопрос крайне важен, но им не исчерпывается вся проблематика этой темы.
Другой вопрос, изучению которого уделяют в США много внимания, — это вопрос о том, существует ли раса как социокультурный феномен, опосредованно влияющий на жизни людей и имеющий, соответственно, биологические последствия. Это вопрос о том, как раса может стать биологической категорией, не являясь таковой изначально. По сути, раса и биология неразрывно связаны: социальное неравенство влияет на биологию угнетаемой группы, что создает впечатление о естественности различий рас [Gravlee. 2009].
Раса как социальная категория оказала огромное влияние на выбор брачного партнера в США. Количество браков внутри рас гораздо выше, чем между ними. Так до 1970-х гг. количество межрасовых браков между «белыми» и «черными» составляло всего 0.1% для белых и 1% — для черных от общего количества заключенных браков. С тех пор этот уровень вырос, но незначительно: по данным 2000 г. сейчас этот уровень составляет 0.3% — для белых и 4% — для черных. Важно отметить, что заключение брака, конечно, не отражает в полной мере всех межрасовых взаимоотношений. Для более точного изучения этих вопросов следует применять индивидуальное анкетирование. Так, в одном исследовании [Hitsch et al., 2004] анализировались анкеты на сайте онлайн-знакомств. 50% белых участников утверждало, что расовая принадлежность не играет для них никакой роли при выборе партнера, при этом 90% из них отвечало на сообщения только белых же кандидатов [Ousley et al, 2009].
При всей критике существования расовых категорий многими отмечается, что сведения о расовой принадлежности индивида (его происхождении) и его биологических особенностях могут быть полезными в различных биомедицинских исследованиях [Jorde, Wooding, 2004, р. 32]. Во всем мире, в особенности в США, существуют определенные межрасовые различия в показателях здоровья. На индивидуальном уровне на показатели здоровья влияют несправедливое отношение и дискриминация. Сообщается о связи расовой дискриминации (по сообщениям самих обследуемых) и повышенного давления крови, рака груди, кальцификации коронарных артерий, ИМТ, абдоминального ожирения, преждевременных родов, низкого веса новорожденных [Lauderdale, 2006], депрессии и других аспектов физического и психического здоровья [Gravlee, 2009].
Исследования этой темы показывают, что причины этих различий лежат не в области генетики, а в области культурных и социальных особенностей отдельных групп населения [Dressier et al., 2005]. Наличие культурных различий и расовых предрассудков оказывает влияние на биологию притесняемой группы. Соответствующие ухудшение показателей здоровья интерпретируется как наличие генетических основ заболеваний, что провоцирует новый виток расизма. Исследователи здоровья предоставляют большое количество аргументов против однобокого восприятия биологии только через генетику, и обращают внимание на наличие фенотипической пластичности и средового влияния на рост и развитие человеческого организма. Подобные исследования вновь поднимают идею того, что человеческая биология био-культурна и представляет собой результат совместного влияния генетики и среды.[при примате социального неравенства между группами над биологической разнокачественностью индивидов в определении специфики этой самой биологии]
Заключение
С течением времени в американской антропологии изменяется понимание рас: на смену классическому типологическому подходу приходит отрицание существования расовых категорий. Такой подход основывается в первую очередь на генетических исследованиях. Однако дальнейшее развитие этих исследований показало возможность разделения человечества на группы на основании как генетических, так и краниологических и других антропологических признаков. Признавая очень высокую внутригрупповую изменчивость, ученые все-таки начали уделять должное внимание и изменчивости межгрупповой. В настоящее время все реже встречается категорическое отрицание любого разделения человечества и все чаще признается важность и практическая значимость понимания биологических особенностей различных групп населения. Исследования рас и этносов находят свое применение в медицине, криминалистике, палеоантропологии, фармакологии и других отраслях знания. Таким образом, продолжение изучения как существующих расовых концепций и стереотипов, так и поиски новых, более точных способов описания изменчивости человечества, имеют высокое значение и широкие перспективы развития.
Благодарности
Работа выполнена в рамках темы «Комплексные антропологические исследования древнего и современного населения» (№01201351784). Автор выражает благодарность Т.Е. Талапановой и М.В. Поповой за помощь в подготовке публикации.
Вестник Московского университета. Серия XXIII. Антропология 2015. №3. С.82-93
Примечание
[1] Иными словами, здесь имеют в виду «расу» расистской мифологии, а не реальные популяции человека, которые называют «расой» советские и следующие им российские антропологи. В отличие от западных коллег, в их стране расизм был преодолён, благодаря чему они не должны, обжёгшись на молоке, дуть на корову, иногда доходя до смешного, как, скажем, уважаемые авторы журнала Science.
Последнее – лучший пример того, что нельзя играть в шахматы с помощью добрых намерений (злых, впрочем тоже). На беду, их общество устроено так, что все межындивидуальные, а тем более межгрупповые различия обывателю выгодно воспринять в терминах «выше-ниже» и «лучше-хуже» с понятными последствиями. Что ставит антирасистски настроенных исследователей в печальное пголожение отца Кабани из «Трудно быть богом». Прим.публикатора.
Рекомендуем прочесть
 Пружинистая поступь большого финансового краха
Пружинистая поступь большого финансового краха Влияние «Великой Рецессии» на неравенство материального положения в обществе с учетом возрастной и расовой принадлежности
Влияние «Великой Рецессии» на неравенство материального положения в обществе с учетом возрастной и расовой принадлежности Неузнанный Дон-Кихот
Неузнанный Дон-Кихот Автомобильный тупик России и мира
Автомобильный тупик России и мира Темная сторона корейского кино
Темная сторона корейского кино От «энергетического поворота» к угрозе биоразнообразию — о положении с охраной птиц в Германии
От «энергетического поворота» к угрозе биоразнообразию — о положении с охраной птиц в Германии О пагубности ошибок причинно-следственной связи
О пагубности ошибок причинно-следственной связи Заявление о Косыгинской реформе и товарище Худенко
Заявление о Косыгинской реформе и товарище Худенко
