

Ещё необходимая работа над ошибками — в продолжение предыдущих
Каскад доместикационных изменений и бонобо
Содержание
Как известно, когда в руках у тебя молоток, любая проблема кажется гвоздём. Что и случилось с замечательной концепцией доместикации (дестабилизирующего отбора) Д.К.Беляева; её лучшее современное изложение см. статьи Л.Н.Трут в Вестнике ВОГиС (2007, 2008). Соответствующий каскад морфологических, гормональных и поведенческих изменений найден не только у одомашненных видов млекопитающих, но и у кур, т. е. может оказаться всеобщим.
Её стали прикладывать ко всем сколько–нибудь сходным процессам, в т.ч. к урбанизации «диких» видов птиц и млекопитающих (а последнюю путали с синантропизацией, чего делать не следует, почему — обсудим ниже). Сам грешен, отождествлял оба процесса, исходя из сходства гормональных сдвигов, обеспечивающих уменьшение стрессируемости и как следствие — снятие блока на исследование нового, обучение в сложное среде, увеличивающееся бесстрашие и другие составляющие когнитивного развития, такие, как рост уровня базального метаболизма и эффективности усвоения пищи. См. все эти сюжеты в пересказе обзора F.Bonier (2012).
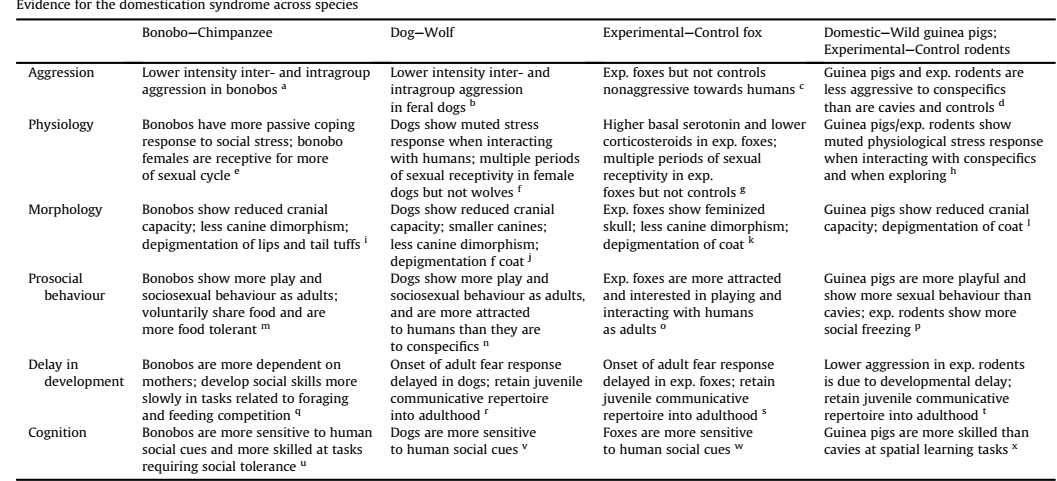
 Я и концепцию самодоместикации бонобо Pan paniscus Брайана Хейра и Ванессы Вудс разделял, хотя уже тогда было ясно, что для её обоснования авторы натягивают сову на глобус. В таблице, что приведена выше, они сравнивают бонобо с обыкновенным шимпанзе, доказывая что его отличия от этого вида того же рода, что домашней собаки от волка, домашней морской свинки — от дикой, лабораторных крыс и мышей — от пасюков и «диких» Mus musculus/M.domesticus, беляевских доброжелательных к нам лис — от диких родичей. Да, однотипная разница обнаруживается везде, кроме восприятия социальных подсказок со стороны людей и сородичей при решении когнитивных задач.
Я и концепцию самодоместикации бонобо Pan paniscus Брайана Хейра и Ванессы Вудс разделял, хотя уже тогда было ясно, что для её обоснования авторы натягивают сову на глобус. В таблице, что приведена выше, они сравнивают бонобо с обыкновенным шимпанзе, доказывая что его отличия от этого вида того же рода, что домашней собаки от волка, домашней морской свинки — от дикой, лабораторных крыс и мышей — от пасюков и «диких» Mus musculus/M.domesticus, беляевских доброжелательных к нам лис — от диких родичей. Да, однотипная разница обнаруживается везде, кроме восприятия социальных подсказок со стороны людей и сородичей при решении когнитивных задач.
В отличие от диких предков, все доместицированные в этом случае «ищут» подсказок людей и/или сородичей (в т.ч. активно стимулируют к этому), и решают не столько своими силами, сколько благодаря «раздельному вниманию» с хозяином и друг другом. Это т. н. «кооперативность познания«, в чём собаки, козы, лошади и пр. «лучше» шимпанзе. А вот бонобо — увы; у них с поиском и предоставлением подсказок также плохо, как у P.troglodytes. Работы же, которыми Хейр и Вудс пробуют этот вывод опровергнуть, показывают лишь большую роль социальной связанности и социальной зависимости у бонобо, безотносительно к когнитивным моментам, где оба вида неразличимы — и чем дальше, тем больше.
Плюс бонобо по всем признакам (морфология, онтогенез, «конструкция» социальной системы) ближе к общему предку, как рода Pan, так и «ветви шимпанзе», с одной стороны, и ветви, давшей австралопитеков, а потом и людей — с другой. Он более архаичен и менее специализирован, обыкновенный шимпанзе — вид более продвинутый и молодой.
Во всех остальных парах таблицы более доместицированный вид оказывается потомком «дикого» предка, а не наоборот.
Преобразования индивидуальности животного при освоении городов
Действительно, здесь стоит задуматься: «в среднем» урбанизационные изменения те же, что при доместикации — «городские» особи «умней», смелей (в новой, потенциально опасной ситуации), «доверчивей» (к людям и технике), но не в частностях — а них-то весь интерес. Несмотря на снижение уровня кортизола, рост серотонина и прочие изменения гормонов и нейромедиаторов по ходу урбанизации (см.главы 6 и 10 Avian Urban Ecology: Behavioural and Physiological Adaptations), особи «городских» популяций не делаются ни «добрей», ни «доверчивей». В среднем агрессивность возрастает пропорционально росту смелости, см. данные по большой синице Parus major. Они относятся к реакции на песни потенциальных захватчиков; то же показано для интенсивности тревожной вокализации, связанной с готовностью окрикивать, не страшась хищников, и клевков при взятии в руки, см. Senar et al., 2017. То же показано для певчего воробья Melospiza melodia, для чёрного коршуна Milvus migrans etc.
Да, дистанция вспугивания снижается, неофобия уменьшается, на новизну «городские» особи реагируют в большей степени исследованием и игрой, нежели страхом и бегством, что отчасти похоже на изменения реакций на человека и новизну одомашненных лис, норок и пр. от агрессивно-оборонительной к дружественной или исследовательской.
См. особенно данные Н.Н.Мешковой и Е.Ю. Фёдорович (1996) о повелении городских домовых мышей в «жилой комнате» в противоположность негородским: первые исследуют компоненты новой и сложной среды, играют с ними, вторые избегают их, поскольку стрессируются. Все эти различия сохраняются при искусственном выкармливании, т. е. отражают изменения на генетическом уровне, поддержанные отбором. Другой пример: смешное исследование интеллекта двух видов врановых, темноклювых сорок Pica hudsonia и короткоклювых ворон Corvus brachyrhynchos, показывающее, что ум и скорость реакции — вещи разные
Сороки в американских городах часто гнездятся поблизости от американских ворон, в т.ч. потому что их агрессивность спасает от разорения гнёзд другими воронами. В Jackson Hole, Вайоминг, исследовалось, как в таком случае обстоят дела с конкуренцией: «платят» ли сороки штраф за обитание на участке более крупного и агрессивного вида, способного отнять корм у мелкого и/или быстрее добыть часть его.
В опытах использовались т. н. Cheeto — популярный у американцев сорт чипсов со вкусом сыра ярко-оранжевого цвета, у обоих врановых он популярен не меньше. Чипсы выкладывали у гнёзд ворон и сорок, а потом смотрели, как быстро оба вида их схватят, кто победит в соревновании за лакомые кусочки и пр. Оказывается сороки быстрей замечают и схватывают чипсы (в среднем на 20 с), вороны же более неофобны и дольше осторожничают; но, определив Cheeto как пищу, они таскают чипсы гораздо ловчее.
Гнездовые пары ворон и сорок быстро выучивают, что чипсы могут быть как у их собственного гнезда, так и у чужого, так что можно «брать своё» и «воровать». Вороны таскают чипсы от сорочьих гнезд в 3 раза чаще, чем сороки от вороньих. Эспозито увеличил сложность тестов, закладывая чипсы внутрь полой колоды; чтобы их взять, птицам надо было тянуть за верёвочку. См. видео этого поведения сорок и ворон соответственно.
Сороки меньше боялись головоломки, смелее брались её решать, и добывали свой чипс где-то на минуту быстрей, чем вороны. Понятно, что для обоих проблема простенькая — в лаборатории врановые решают куда более сложные задачи (скажем, протоорудийные). Но это был один из первых таких опытов с дикими птицами, и автор был более заинтересован в экологических, а не когнитивных аспектах (увы, наши исследования интеллекта разных видов птиц в естественной среде обитания он не цитирует, хотя они куда более совершенны).
Далее Эспозито помещал чипсы посередине между гнёздами ворон и сорок и смотрел, кто среагирует раньше. В этом случае те и другие одинаково быстро отыскивали чипсы и «понимали» что это лакомство (в отличие от выкладывания чипсов на «ничейную территорию»).
Однако вместо того чтобы самим искать чипсы, вороны «использовали» поведение сорок как знак наличия корма, после чего практиковали клептопаразитизм: ждали, когда сорока спустится и попробует взять корм, затем спугивали её и сами брали искомое. Со временем сороки отказываются не только брать что-либо, но даже спускаться на землю. Вороны успешнее в воровстве (а сороки легче им уступают) возможно потому, что (в отличие от примерно вдвое меньших сорок) меньше боятся нападения хищников на земле, менее бдительны там — а спустившиеся сороки им уже «показали», что корм безопасен. Возможно, просто потому что они физически сильнее, им легче отпихнуть конкурентов (довод «против» этого предположения — данное преимущество сохранялось бы и на «ничейной территории»).
В заключение Эспозито показывает, что при гнездовании рядом с воронами сороки действительно получают защиту от хищничества, но общий успех размножения не возрастает, видимо, потому что «плата» от уменьшения поступления корма вследствие вышеописанных проблем с воронами сравнима с «выигрышем». Дальше он будет пробовать расшифровывать конкретные механизмы этого факта (Источник).
Ещё пример: в последние 6-7 лет мы с Г.С.Ерёмкиным наблюдаем у обоих наших ястребов (тетеревятника Accipiter gentilis и перепелятника A.nisus) вполне сложившуюся тактику охоты в городских кварталах. Они гнездятся в лесных массивах, например, в Мытищинском и Пироговском лесопарках, а охотиться летают в городские кварталы. И именно на лесных птиц: на дроздов и пр. питающихся ягодами (тетеревятник) и на «просачивающихся» через город мигрирующих (перепелятник). Ястреб спокойно сидит на границе между лесом и застройкой, на крыше детсада или другого невысокого строения, где его, видимо, жертвы «не ждут», «спиной» к лесу, лицом к городу, и, взлетая, атакует птиц, движущихся «назад», из города в лес. Скорей всего, при движении «из города», настороженность сильно падает сравнительно с движением «в город» или «по городу». Тут-то ястреб и появляется.
 Возвращаясь к домовым мышам, испытавшим «развивающий эффект» урбосреды: это синантропный вид, живущий в наших постройках и непосредственно контактирующий с человеком. У видов-синурбанистов — вяхирей, чёрных дроздов, больших синиц, тетеревятников, черных и серых ворон и пр., формирующих городские популяции, устойчиво живущих в городе, но часто не в застройке, а на «островах» природных территорий внутри урболандшафта или в «прослойках» озеленения домов и кварталов, картина другая, о чём см. книгу и статью.
Возвращаясь к домовым мышам, испытавшим «развивающий эффект» урбосреды: это синантропный вид, живущий в наших постройках и непосредственно контактирующий с человеком. У видов-синурбанистов — вяхирей, чёрных дроздов, больших синиц, тетеревятников, черных и серых ворон и пр., формирующих городские популяции, устойчиво живущих в городе, но часто не в застройке, а на «островах» природных территорий внутри урболандшафта или в «прослойках» озеленения домов и кварталов, картина другая, о чём см. книгу и статью.
C другой стороны, не то чтобы у «городских» видов усиливался интеллект как таковой (или толерантность к людям, или способность к обучению — любые признаки, подаваемые как новообразования, «сходные с доместикацией» — хотя отдельные данные есть и об этом), сколько всегда и везде они лучше «дозируют по цели» интерес и исследование vs недоверие к новому, доверие–недоверие к людям и технике, агрессию и т.д. (Lowry et al., 2013). Так, калининградские вяхири Columba palumbus, в городе подпускающие вплотную как сизари, за городом на кормёжке осторожны не менее «диких». Это же верно для домовых, а тем более полевых воробьёв и пр.
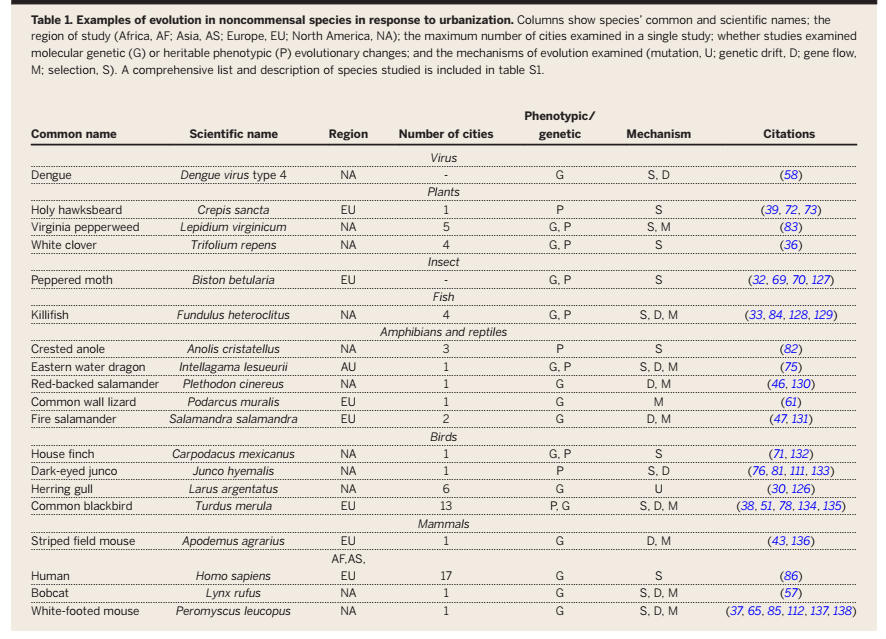
Микроэволюционные процессы, фиксированные у разных видов в городах (G — генетические изменения, Р — наследуемые фенотипические). Источник: Johnson, Mushi-South, 2017. Изменения, обсуждаемые в тексте, и поддерживающий их отбор на уровне генов, представляют собой как бы «отдачу» от этих инноваций, специфических для каждого из рассматриваемых видов. Они приспосабливают организацию к урбосреде «вообще», чтобы могла устойчиво существовать там, почему общи для разных видов птиц
Такие же наблюдения есть в «городских» популяциях разных видов врановых, вне города отличающихся существенной неофобией (Greggor et al., 2016). И наоборот: быстрая и успешная урбанизация чёрной вороны Corvus corone в Иркутске завершилась без какого–либо увеличения толерантности к человеку, по мере освоения жилой застройки осторожность скорей возрастала (Липин и др., 1983, цит.по Фридман, Ерёмкин, 2009). У освоивших «зелёные острова» такие примеры встречаются на каждом шагу: устойчиво обитая там и успешно гнездясь, они «умеют быть» и сторожкими, и незаметными, как европейский тювик Accipiter brevipes в урболандшафте ряда мест Северного Кавказа. Сравнивая популяции ряда видов (домовый воробей, большая синица, чёрный дрозд, певчий воробей и т.д.), в разной степени освоивших урбосреду, видим, что все составляющие поведенческого синдрома урбанизации — неофобия в отношении пищевых объектов и предметов, восприятие риска, большие смелость, активность и склонность к инновациям в экологии/поведении — у более урбанизированных популяций в сравнении с менее показывают значимо большую дисперсию, а не только изменения средних значений (Bókony et al., 2012; Tryjanowski et al., 2015; Tryjanowski et al., 2016). В т.ч. большая продвинутость видов по названным показателям ведёт к всё большему сближению их (в среднем укорачивающихся) дистанций взлёта (Ducatez et al., 2016).

В городе безопасней, чем вокруг - в частности, для канадских казарок
«Городских» птиц здесь уместно сравнить с меланистическими хомяками, изученными С.И.Гершензоном (1941) (они живут с неопределённостью, созданной паводками) и черно–бурыми лисами на Алеутских островах (там сходная неопределённость создана скачками ледовой обстановки). Тем и другим важно сохранять устойчивость поведения, чтобы оно вопреки этим «помехам» достигало целей, в диапазоне от выживания до размножения, а) гарантированно и б) в приемлемое время. Также и в городе особям надо приспосабливаться не к каким–то конкретным воздействиям города вроде шума, света, загрязнения, риска разбиться о стёкла и пр.: ведь при сильной гетерогенности урбосреды действие каждого слишком локально, а место обитания и население вида в каждом из мыслимых «мест» их воздействия слишком непостоянно, чтобы мог идти направленный отбор.
Да и времени на него заведомо не хватает, см. таблицу со сроками из вышеприведённой книги. Анализ Alberti et al., 2017 подтверждает, что из всех форм антропогенных трансформаций ландшафтов урбанизация ведёт к самым быстрым микроэволюционным изменениям, как плане приспособлений, так и в плане дивергенции популяций.

Размеры городских ареалов, происхождение основных синантропных видов птиц и млекопитающих, места исследований микроэволюционных процессов, отмеченных в предыдущей таблице. Источник тот же.
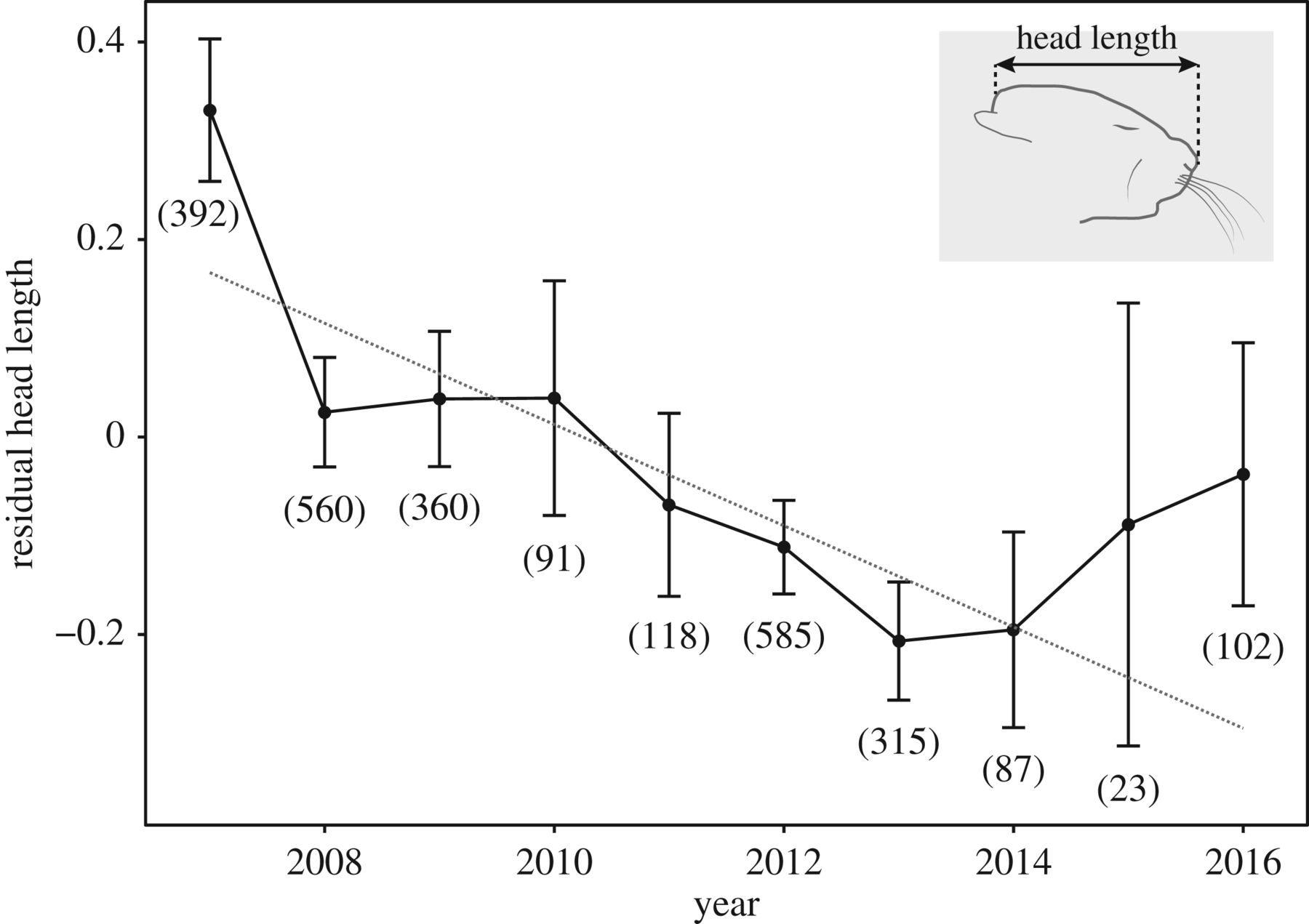
И, действительно, доместикация по Беляеву связана в первую очередь с изменениями клеток нервного гребня, отсюда пегости и укорочение морды (в т.ч. при самодоместикации, вроде обнаруживаемой у мышей, см. Geiger et al., 2018). Даже у синантропных видов пегость возрастает не сильно. А у урбанизированных, живущих устойчиво внутри городских ареалов, но на «зелёных островах», не в застройке (как ястреб-тетеревятник, малый и белоспинный дятлы, и пр.) пегостей больше не становится. В ряде случаев урбанизация, наоборот, сокращает пегости (юнко, освоившие г.Сан-Диего в Калифорнии — там исчезают белые каймы хвоста). А вот что прогрессирует при урбанизации разных видов, кроме понятных адаптивных изменений размеров клюва (1-2), длин крыльев, частоты звучания песни и прочих вокализаций (см.табл.ниже и главу той же монографии), так это меланизм.
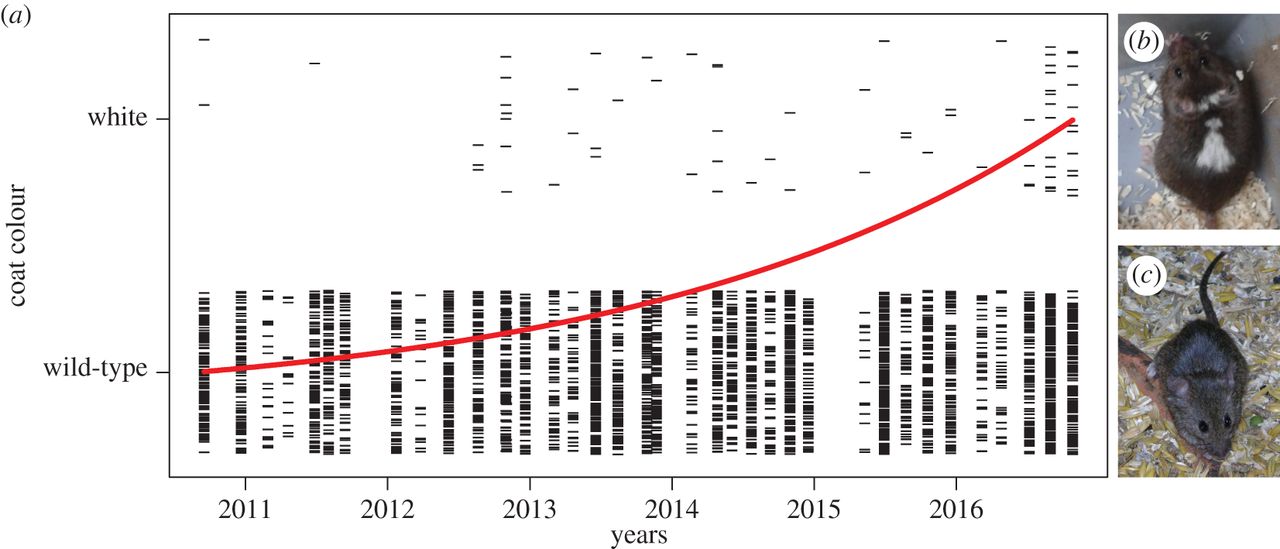
Быстрое увеличение пегостей и укорочение морды при самодоместикации домовых мышей. Из: Geiger et al., 2018
 Другой важный момент: в оценке уровня агрессивности, кроме «мощи» одноимённой реакции на соответствующие стимулы (вид захватчика или конкурента на твоём месте, его пение, демонстрации и пр.) важно последействие, «расталкивающий» эффект произведенной агрессии, как далеко и насколько надёжно она удалит оппонента? И в большинстве работ показано, что даже когда у «городских» особей первая составляющая агрессивности ниже, вторая во всех случаях выше. Даже «обрушивая» на претендентов больше агрессии или более интенсивных действий её репертуара, «городские» птицы в противоположность негородским не удаляют даже побеждённых противников из зоны активности, а выстраивают с ними на той же территории более сложные отношения, частью персонализованные, частью доминантно–субординантные. Это т,н. поведенческий «островной эффект» (island effect): большая терпимость к сородичам, «зажатым» все вместе в «островах» местообитаний.
Другой важный момент: в оценке уровня агрессивности, кроме «мощи» одноимённой реакции на соответствующие стимулы (вид захватчика или конкурента на твоём месте, его пение, демонстрации и пр.) важно последействие, «расталкивающий» эффект произведенной агрессии, как далеко и насколько надёжно она удалит оппонента? И в большинстве работ показано, что даже когда у «городских» особей первая составляющая агрессивности ниже, вторая во всех случаях выше. Даже «обрушивая» на претендентов больше агрессии или более интенсивных действий её репертуара, «городские» птицы в противоположность негородским не удаляют даже побеждённых противников из зоны активности, а выстраивают с ними на той же территории более сложные отношения, частью персонализованные, частью доминантно–субординантные. Это т,н. поведенческий «островной эффект» (island effect): большая терпимость к сородичам, «зажатым» все вместе в «островах» местообитаний.
На деле же «городские» птицы в сложной и разнообразной среде делаются «просто умнее». В ходе успешной урбанизации она для них делается развивающей, тогда как при «входе» в урболандшафт стрессировала, а то и подавляла, см. обзор F.Bonier, 2012. Как дальше увидим, гены, оказывающиеся под отбором в процессе урбанизации «диких» видов, у нас грешных решают примерно те же задачи (многозадачность, мысленное оперирование сразу несколькими объектами, распределение внимания, отслеживание отсроченного вознаграждения и пр.).
Из этой картины урбанизации «диких» видов следует один интересный момент. Смелость вкупе с готовностью исследовать незнакомую и потенциально опасную ситуацию вместо того, чтобы отступать от неё, развивая пассивно-оборонительные реакции, — важная характеристика «лучших» особей в природных популяциях; в городе эти качества обретают все. Носители конкурентной стратегии [3], они поддерживаются отбором не сами по себе (иначе «заполонили бы» популяцию), а для устойчивого воспроизводства пространственно-этологической структуры последней. Там их поведение играет роль индуктора и организатора, управляющего активностью приверженцев противоположной патиентной стратегии. Благодаря ему паттерны активности «худших» особей «помещаются» в пространственные и временные лакуны между активностью «лучших», так что те и другие образуют скоординированную «карусель» или «хор».
Одновременно каждая из альтернатив всё больше специализируется в условиях собственной «лакуны» (которая одновременно коммуникативная ниша). Что делает их ещё более комплементарными друг другу в плане состава поведения и взаимозависимыми в функциональных связях, укрепляет социум и пр. См. обзоры коммуникативных сетей между самцами разного «качества» (или видов, занимающих разное место в межвидовой иерархии) у певчих птиц.
То есть специфика процесса урбанизации «диких» видов состоит не в отборе наиболее умных особей в формирующейся городской популяции (или видов с более развитым мозгом/пластичным поведением в городскую среду), как часто считается. Нет, главное содержание урбанизации – микроэволюционное изменение, в ходе которого с основной массы «худших» особей, приверженных патиентной стратегии, снимается блок на развитие смелости, уменьшается неофобия, увеличивается если не способность к обучению, то готовность учиться и пр.
Точно также как в процессе доместикации, это происходит за счёт снижения реакции стресса, вызванного неопределённостью и новизной, с увеличением способности дозировать её сообразно реальной стрессогенности событий и объектов, с которыми столкнулось животное, без мощной добавки в связи с неофобией. Одновременно сокращается до минимума пассивно-оборонительная реакция в отношении просто новых и неизвестных объектов, падает дистанция вспугивания при приближении безоружных людей и медленно работающей техники, но в случае вооружённых людей или быстро приближающихся машин она остаётся значительной. Когда поведенческие изменения, уменьшающие беспокойство людьми, техникой, или гибель от неё оказываются недостаточны, они «тянут» за собой изменения в морфологии. См. укорочение крыльев у белолобых ласточек Petrochelidon pyrrhonota, снижающее риск гибели на автострадах.
Всё это не отменяет разделения на «лучших» и «худших» особей в ходе конкуренции уже в городской популяции, только идёт оно по другим основаниям, нежели в «сельских» и «лесных» популяциях. В городе «лучшие» особи более управляемы средой и менее зависимы от социальных отношений в группировках, «худшие» — наоборот. Легко видеть, что в негородских популяциях картина противоположная. «Городские» особи тем легче оставляют данное поселение и переселяются в другие с целью увеличить репродуктивный успех (что обычно и удаётся), чем успешней они размножались. «Негородские», наоборот, в случае успешного размножения максимально привязаны к данной группировке и данному составу соседей, консервативны как территориально, так и в отношении уже поддерживаемых социальных связей. Всё это меняется лишь при неудаче репродукции — ненахождении партнёра, гибели гнезда или выводка.
В урбанизацию вовлекаются другие гены
Действительно, как стали исследовать гены, оказывающиеся под положительным отбором при урбанизации «диких» видов (SERT, DRD4, ADCYAP1), выяснилось, что они совершенно иные, чем при доместикации домашних. Как пишет мне В.В.Суслов (Ин-т цитологии и генетики СО РАН):
«В литературе есть работы по поиску генов-кандидатов урбанизации, т.е. генов, для которых в урбанизированных популяциях птиц выявлено усиление адаптивной эволюции или просто усиление изменчивости. Пока выявлено три таких кандидата — SERT, DRD4, ADCYAP1, причем SERT и DRD4 – у разных видов птиц или у одного вида, но из популяций разных городов с разным стажем урбанизации. Быстрее всего на селекцию урбанизацией отвечает SERT, по нему можно гомологические ряды у птиц-урбанистов строить. SERT — это ген серотониновой стресс-лимитации, он помогает легче переносить любой стресс (т.н. серотониновая эйфория). Ген дофаминового рецептора DRD4, кстати, консервативен у птиц и млекопитающих.
В нашем институте есть модель беляевских крыс, селектированных, как и лисы, на агрессивное и толерантное поведение по отношению к человеку. И есть другая модель — неселектированных мышей с индивидуальным опытом побед и с опытом поражений. Грубо говоря, мышей попарно ссаживают сильного со слабым и дают им подраться 5 минут. Затем к тому же сильному сажают другого слабого, а к тому же слабому – другого сильного и так до тех пор, пока не сформируется устойчивый, но ненаследуемый комплекс поведения победителя – наглый задиристый, легковозбудимый мышь, охотно лезущий в драку и депрессивный мышь-проигравший, пассивно избегающий драки, неофобный, словом, пуганая ворона куста боится.
Итак две модели поведения – одна наследственная, другая – модификационная. Обе связаны со стрессом. По ним у нас в институте сделаны чип-секи, выявляющие дифференциально экспрессирующиеся гены в мозгу – соответственно у беляевских крыс дифференциально экспрессирующиеся у наследственно агрессивных к человеку vs наследственно толерантных или у мышей дифференциально экспрессирующиеся у особей с опытом «накопления побед» vs «накопления поражений».
Мы решили поискать гены SERT, DRD4, ADCYAP1 среди генов этих чип-секов. Оказалось, что среди топ-10 и топ-20 дифференциально экспрессирующихся генов беляевских крыс из этой тройки генов вообще нет! Ожидали чего угодно, но не этого. Cреди дифференциально экспрессирующихся генов у мышей выявляется из первой тройки лишь ген SERT из топ-20, но он – не показатель, он будет облегчать любой стресс, его экспрессия повышена и у проигравших, и у победителей по сравнению с контролем, так что неудивительно, что отбор ловит его и у птиц-урбанистов.
Поэтому удивительно, что у беляевских крыс отбор его не поймал, хотя у птиц-урбанистов он его ловит, а норма реакции экспрессии, как показывает эксперимент на мышах, у него широкая, его уровень легко можно поднять и надолго оставить высоким – словом, всё это благоприятствует генокопированию. Зато в обоих моделях вместо DRD4 очень хорошо дифференциально экспрессируется DRD2.
Это контринтуитивно – различия в спектрах SNP по DRD4 выявляются у больших синиц уже на 4 поколении селекции, правда там селекция шла на поисковое поведение. Но беляевские агрессоры и доместиканты ведь тоже сильно отличаются по поисковому поведению и у них-то селекция идет гораздо дольше. История с SERT также контринтуитивна. Как все это объяснить – мы не знаем. Единственно, что могу сказать от себя, что это еще один факт, показывающий, что Беляев открыл яркий, красивый, интересный, но все-таки очень специфичный и редкий в природе отбор, почему прямо экстраполировать результаты Беляева на природную эволюцию… нельзя. Год назад я думал прямо противоположно, но потом меня смутило во-первых, что в гомологических рядах Вавилова редкие варианты признака доминантны в центрах происхождения, а в гомологических рядах Беляева – рецессивны или полудоминантны.
Как правило, выживают посредством доминантных генов и вопреки всякой скрытой изменчивости (генам полудоминантным и рецессивным). Если скрытая изменчивость дает полезный фенотип – отбор сделает все, чтобы она перестала быть скрытой. Во-вторых (и это главное) гомологические ряды Вавилова равно выявляются и у доместицированных, и у диких популяций /видов/родов, а гомологические ряды Беляева – только у доместикантов. И это не зависит от таксономического статуса – похожи будут лиса, собака, корова и лошадь, но у диких видов псовых признаков беляевских собак и лис нет, если не считать таким признаком банальную разницу в размерах (волк и кустарниковая собака; дог и чихуахуа – но тут классическое селекционистское объяснение просто в глаза бьет)…»
Заинтересовавшись, есть ли пересечение пересечение генов, вовлечённых в урбанизацию «диких» видов птиц, с генами, связанными с доместикацией собственно беляевских лис, мы специально посмотрели по соответствующей статье и не нашли.
«Если они есть у птиц и лис, то в неидентифицированных областях типа ENSCAFG00000032444, которые не с чем сравнить у птиц и людей. Общая картина у лис, видимо, такая же, как у крыс – 146 дифэкспрессирующихся генов у доместицированных и диких лисов отвечают за общую активность гормональных и нейротрансмиттерных систем и за регуляцию их онтогенеза, но не за решение конкретных задач, в отличие от генов, вовлечённых в урбанизацию.
Так как DRD4 оказался связанным с приоритезацией внимания и объемом рабочей памяти, интересно посмотреть гены у людей, могущие решать сходные задачи: влияющие на объем рабочей памяти, на категоризацию, на языковую беглость (когда нужно назвать за ограниченное время термины одной категории, например, животные, профессии и т.д.), на решение задач вопреки отвлекающим действиям. Заметим, что все они находятся вне применяемых в ИЦиГ методик тестирования на агрессивность/доместикатность. Пересечений с «лисьим» списком пока не найдено (из просмотренных 20 генов нет ни одного ни в «лисьем», ни в «крысином» списках). Интересно ещё исследовать гены, связанные с пением у птиц, так как рецепторы DRD отметились и там, а у людей они вовлечены в распознавании нюансировки речи».
См. соответствующие тезисы в трудах BGRS’SB 2018 и V. Suslov. Ontology of homologous series. BGRS/SB-2018. The 11th International Conference//Integrative Bioinformatics and Systems Biology (WIBSB-2018) First Sino-Russian Workshop. 22–23 August, 2018. Abstracts. Novosibirsk, ICG SB RAS, 2018. P. 61. DOI 10.18699/WIBSB-2018-46).
Разница в стрессорах, к которым приспосабливаются: неопределённость vs страх
Нужно отметить, что стрессор, к которому приспосабливаются, при урбанизации совсем другой, чем при доместикации. Во втором случае это сам человек и связанные с ним вещи, постройки и пр., звероводам важно, чтобы лисы в вольере на него не бросались и от него не шарахались на проволоку, т.ч. это отбор дружелюбных к людям и толерантных к производимым ими изменениям остановки. В городе такому отбору подвергаются в лучшем случае «традиционные синантропы» — домовый воробей, галка, городская и деревенская ласточки, пустельга, сипуха и пр., но именно они наиболее уязвимы из-за динамичности современного города, его постоянных перестроек, идущих одновременно в разных масштабах. Это подрывает постоянные перестройки кварталов связность обычных мест гнездования, сокращает обычно используемые корма, притом что птицы не успевают отыскивать новые, появляющиеся во вполне достаточном количестве и пр. Они сокращают численность, уходят из «ядер» агломераций в регион на микроурбанотерритории и т. д.
А виды-синурбанисты, живущие на куда меньшем, сильней изолированном и более подвижном «архипелаге» природных территорий или их близких производных, остаются устойчивы, наращивают численность и кто быстрее, кто медленней, расширяют ареал центробежно, проникая всё дальше в застройку, используя всё меньшие фрагменты древесно-кустарниковой растительности и/или луговых/болотных участков (в зависимости от видовой специфики). И, если урбанизация пошла — они вполне благополучны. Если что не так, она остановится; мне известны всего 2 случая, когда уже возникшая городская популяция исчезла — рябинник Turdus pilaris в Кёнигсберге в 1930е (проник в 1900-х) и сорока в Донецке в 90е (проникла в 1970е, см.тут, с.324-325).
Откуда такая разница? Главный стрессор, к котором приспосабливаются виды-синурбанисты, в современном городе другой, чем у доместицированных видов и у видов-синантропов в доиндустриальную эпоху. Это изменчивое и нестабильное «кружево» местообитаний. Какой обычный городской вид ни возьми, даже сизаря, чёрного дрозда, вяхиря, большую синицу с лазоревкой, не говоря уж о видах, больше привязанных к городским лесам и водоёмам, не гнездящихся на застроенных территориях. Везде их местообитания не сплошные, а представляют собой «архипелаг».
В отличие от подобного «архипелага» негородских популяций тех же урбанизированных видов дроздов, воробьёв, юнко и т. д. видов, изученных в плане урбанизации и особенно гормональных сдвигов по ходу этого процесса. «Острова» городских местообитаний исключительно нестабильны: старые сокращаются, делятся, меняют конфигурацию, появляются новые в неожиданных и непредсказуемых для птиц местах. В этих условиях выигрывают особи, которые своевременно уходят с первых и переходят на непредсказуемые местообитания. При этом местообитания, наиболее входные с природными аналогами часто оказываются «экологический ловушками». Концентрирующиеся там и жёстко следующие видовому стереотипу особи с высокой вероятностью губят потомство или гибнут сами. А успешное размножение достигается в более изменённых местообитаниях, требующих выработки нетипичных кормовых методов, нетрадиционных мест гнездования и других приспособлений.

В Великобритании успешно урбанизировались сапсаны, реинтродуцированные в 1989 г. красные коршуны…
Всё это создаёт средовой стресс, воздействующий на популяцию. Это популяции «требуется» перераспределять население по «островам» меняющегося размера и «качества» каждый раз заново. Так изменяется «конструкция» пространственно – этологической структуры популяции в одну и ту же сторону у всех урбанизированных видов, даже неродственных и экологически различных, причём не только в городе, но и прямо со «стартовых позиций» урбанизации, проходящих по кольцу городских лесов. Буквально в последние годы это случилось у московских красноголовых нырков, и ряда других видов, дотоле не показывающих такой склонности (возможно в связи с истреблением последних сходных с природными местообитаний в городе, почему им пришлось осваивать сильно изменённые местообитания и меняться самим в поведении, местах гнездования или способах кормления, чего ранее избегали). Т.е. больше всего птиц «беспокоит» риск не найти подходящих местообитаний в пункте, выбранном для следующей стадии годового цикла. Или риск, найдя природное местообитание, не встретить партнёра, потерять кладку и выводок, т.е.оказаться в «ловушке».
Конфигурация «пятен» меняется постоянно, быстро и непредсказуемо для особей, т.ч. значительную часть времени приходится их искать по всему городу пригодные сейчас для жизни, а найдя – в любой момент быть готовыми «пропустить ход» в связи с быстрым изменением найденного участка. Это создаёт стресс, «изнашиваюший» особей, приводящий к повышению заболеваемости, несмотря на большую продолжительность жизни и возрастающую устойчивость к «островному эффекту». Это в точности тот же средовой стресс, который создается неопределённостью паводков для хомяков (спасающихся на незатапливаемых возвышенностях, расположение и доступность которых тоже непредсказуемы для зверька непредсказуема).

…и завезённые попугаи Крамера Psittacula krameri
Рекомендуем прочесть
 Урбанизация «диких» видов птиц/млекопитающих как доместикация домашних
Урбанизация «диких» видов птиц/млекопитающих как доместикация домашних Урбанизация как основной фактор негативного влияния на местообитания диких животных Московского региона
Урбанизация как основной фактор негативного влияния на местообитания диких животных Московского региона Смелость города берёт
Смелость города берёт «Город как заказник»: (пока?) упускаемая возможность
«Город как заказник»: (пока?) упускаемая возможность Модели урбанизации «диких» видов птиц
Модели урбанизации «диких» видов птиц Некоторые пути сохранения биосферы при урбанизации
Некоторые пути сохранения биосферы при урбанизации Парфорсная эволюция как механизм быстрой адаптации
Парфорсная эволюция как механизм быстрой адаптации Методология изучения рекреационного воздействия на городские леса и подходы к проектированию рекреационных объектов
Методология изучения рекреационного воздействия на городские леса и подходы к проектированию рекреационных объектов
