

Продолжение темы красивых теорий, отвергнутых из-за сопротивления некрасивых фактов
«Когда биологи говорят о происхождении видов, то само собой подразумевается, что те виды, которым выпало жить бок о бок, будут разительно отличаться друг от друга. Конкуренция заставляет искать альтернативные способы приспособляемости к среде. В противном случае, если виды живут в сходных экологических нишах, обладают близкими признаками и непосредственно контактируют, один из них окажется более приспособленным, а второй просто вымрет. Это как будто подтверждается многочисленными данными, однако зоологи из Оксфорда (Великобритания) полагают, что данные неверно интерпретируются. В журнале Nature Джозеф Тобиас (Joseph Tobias) и его коллеги сравнивают между собой более 90% видов печниковых (Furnariidae), одного из самых разнообразных птичьих семейств. При этом исследователи сопоставляли их не по всем признакам, а по некоторым — вроде строения клюва, ног и характера песен. Морфологический и вокальный сравнительный анализ подкреплялся генетическим. Чтобы собрать данные по 350 родам, пришлось обратиться не только к полевым исследованиям, но и к музейным материалам, а чтобы проанализировать массу генетических сведений, зоологи объединились с коллегами из США и Швеции.
Различия и сходства между видами соотносили с их удалённостью друг от друга [на самом деле – с временем дивергенции от общего предка, посчитанным по молекулярным филогениям, чтобы отделить сестринские виды от видов-двойников, которые очень похожи друг на друга, иногда до неотличимости – но менее родственны друг другу, чем обычные сестринские. Прим.публикатора].
См.схему родства между разными родами печников, демонстрирующая заодно разницу в строении клювов у разных групп (рис. из статьи).
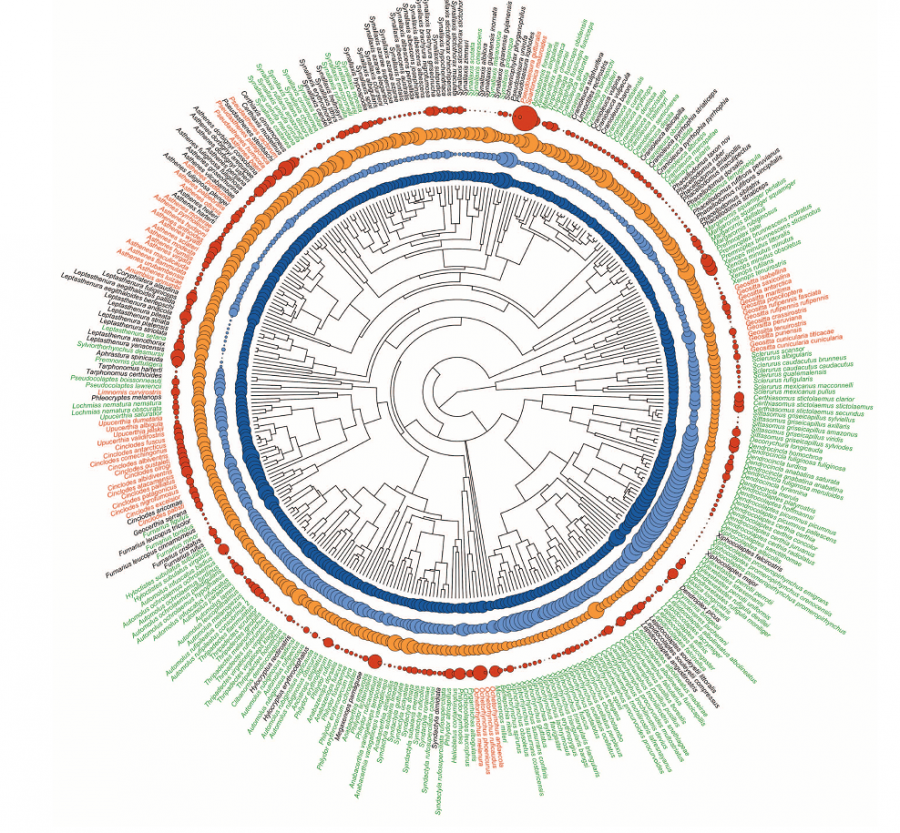 В итоге был сделан вывод, что по анатомическим признакам виды, живущие рядом, отличаются не больше, чем живущие «раздельно». Что же до вокальных особенностей, то голоса непосредственных соседей оказались даже более схожи, чем у видов, друг с другом не встречающихся. Последнее авторов работы весьма смутило, так как считается, что виды, живущие бок о бок, будут изо всех сил стараться петь по-разному.
В итоге был сделан вывод, что по анатомическим признакам виды, живущие рядом, отличаются не больше, чем живущие «раздельно». Что же до вокальных особенностей, то голоса непосредственных соседей оказались даже более схожи, чем у видов, друг с другом не встречающихся. Последнее авторов работы весьма смутило, так как считается, что виды, живущие бок о бок, будут изо всех сил стараться петь по-разному.
Форма клюва у печниковых может быть весьма разной, от строго прямой до сильно изогнутой, однако сильные модификации тут возникают лишь в том случае, если вид жил в изоляции и с себе подобными не контактировал. В случае же с песнями взаимный контакт, похоже, вообще приводит к обратному результату: песни начинают сближаться. См.рисунок.


Всё это, повторим, настолько идёт вразрез с господствующей точкой зрения, что возникает вопрос, почему до сих пор все считали (и считают) иначе.
Исследователи объясняют это тем, что до сих пор никто не обращал внимания на сравнительный возраст видов. То есть, например, два родственных вида могут жить рядом, но при этом один из них будет намного старше другого. В таком случае временна́я изоляция успеет внести в «старший» вид серьёзные изменения.
Генетический анализ позволил учёным сравнить виды одного возраста, что и поменяло картину сходств и различий. Хотя были и исключения, в целом, по словам авторов работы, закономерность получается именно такая: близкое соседство вовсе не подстёгивает гонку эволюционных различий, в отличие от жизни без соседей-родственников.
Как это объяснить, зоологи пока не знают, ограничиваясь лишь простым описанием того, что увидели при сравнении видов. Однако можно предположить, что, к примеру, сходство в вокальных сигналах должно сослужить близкородственным видам хорошую службу в обороне территории от чужаков. Ведь территорию часто приходится защищать не только от нарушителей из близкородственного вида, но и от многих других видов, и в этом случае общий сигнал тревоги способен объединить силы.
Впрочем, это лишь предположение, требующее научной проверки; кроме того, обнаружившуюся странность в сходстве-несходстве родственных видов хорошо бы испытать на других группах животных и с другими признаками: мало ли, может, эти выводы действительны только для печниковых и только для песен, ног и клювов этих птиц».
Источник compulenta
P.S. Это отличная невязка с «общим местом» из учебников теории эволюции[1], что дифференциация видов по эконишам «делается» конкуренцией в зоне симпатрии, а дифференциация видов совпадает с процессом расхождения по нишам или по крайней мере с ним существенно связана. Отсюда вывод, что дивергенция близких видов по морфологии, экологическим предпочтениям и пр. признакам «делается» преимущественно конкуренцией в зоне симпатрии, которая «разводит по нишам», а не филетической эволюцией, обосабливающей ту и другую форму, когда они были изолятами в аллопатрии.
Однако чем дальше, тем хуже эта концепция соответствует фактам, полученным в разных областях, но согласно оказывающим, что и обособленность видов, и их дивергенция по признакам происходят. Во-первых, обособление видов процесс «автоматический», связанный с накоплением побочных последствий изоляции в периферийных изолятах. Он не ускоряется конкуренцией с появившейся близкой формой (поскольку идея «усиления», reinforcement, и смещения признаков не работает).
А вот сорван быть может — из-за гибридизации, формирующей устойчивую гибридную зону, через которую постоянно идёт «обмен» генами, притом что не происходит каких-то попыток «заткнуть течь» и «защитить генофонд» отбором на совершенствование изолирующих механизмов, смещение признаков в зоне симпатрии и пр. То есть максимум дивергенции близких видов связан с давностью их обособления как репродуктивных и коммуникативных общностей, с раздельной эволюцией в условиях аллопатрии, зачастую далёких географически. А не с взаимодействиями в зоне симпатрии, будь то конкуренция и следующая из неё дифференциация ниш, или отбор на совершенствование изолирующих механизмов и следующее из него «усиление» со «смещением признаков».
Далее, виды, воспринимаемые как «близкие» из-за совместного обитания в обычных местах наблюдений, вроде травяной и остромордой лягушки, белобровика и рябинника, обосабливаются на деле географически далеко друг от друга, в условиях аллопатрии, их видовые адаптации отражают условия очень разных местообитаний. Если всё прошло хорошо, они долго пробыли в изоляции, при вторичном контакте им ничего не грозит — «слияния» через гибридизацию не происходит (или постоянной «течи» генов через устойчивую гибридную зону). Соответственно, тогда в зоне симпатрии им ничего не мешает развивать одни и те же (вторичные) адаптации для приспособления к одним биотопам. Тем более они — таки родственны, и это родство «проявится» в параллельности приспособлений, при сохранении разности присосабливающихся структур.
Косяк нынешнего варианта эволюционной теории в том, что он больше значения придаёт адаптивной гибкости, чем филогенетической косности, а она значима даже в самых «мелких» масштабах микроэволюционных процессов не менее первой.
Данные по печникам подтверждают мысль Лео Суреновича Степаняна («Надвиды и виды-двойники в авифауне СССР», М.: Наука, 1983), что в зоне симпатрии близких видов изменения в сторону большего сходства встречаются не реже, чем расхождение в признаках. Далее, во вторичном контакте близких форм чаще и «усиления», и «смещения признаков» фиксируется прямо противоположная ситуация, именно сближение признаков форм по направлению к зоне контакта, часто до такой степени, что затрудняет определение.

Эти случаи обычны настолько, что Л.С.Степанян (1983: 22-23, 27-33) специально для них предложил термин «возвратный изоморфизм». Забавный момент состоит в том, что такое сближение случается чаще, чем «усиление» именно в ситуации «неполностью разделившихся форм» (видов in statu nascendi), когда, согласно модели Добжанского, должен идти противоположный процесс «усиления». Скажем, вот примеры из авифауны Забайкалья.
Примером возвратного изоморфизма может быть пара горных вьюрков Leucosticte acrtoa и L.brandti. Первый вид географически более изменив, чем второй. Симпатричны они на Алтае: там обитает раса L.b.margariticea, наиболее трудноотличимая от местных L.a.arctoa. Другой пример – белоголовый сип Gyps fulvus и кумай G.hymalayensis. Второй вид узкоареален, монотипичен; морфологически он ближе всего к широкораспространному первому в зоне симпатрии. Причём в большинстве случаев здесь речь о гибридизации не идёт.
Третий – обыкновенная Сerthia familiaris и короткопалая пищухи C.brachyfactyla. Данные виды-двойники хорошо различаются песней, внешней морфологией – сильно хуже (несколько признаков, хорошо видимых только в руках). На Кавказе короткопалая живёт на Западном и Северо-Западном Кавказе. В таких же местообитаниях (влажные субтропические леса) Восточного Кавказа, вроде Талыша, её нет, но там живёт исключительно сходный с ней подвид обыкновенной persica – настолько, что крупные орнитологи неверно определяли добытые экземпляры.
Т.е. необычайное сходство симпатрических видов-двойников объясняется как вторичное явление, произошедшее уже после после перекрывания их ареалов. Такие двойники сохраняют высокий уровень генотипических отличий, сформировавшихся ранее в условиях аллопатрии и сильной географической разнесённости мест происхождения. Это также работает на млекопитающих – скажем, полёвках рода Terricola.
Как достигается это особо сильное сходство в случаях возвратного изоморфизма – не знаю; есть подозрение, что механизм, каким бы ни был, общий со случаями исключительно сильного межвидового сходства, о котором уже писал в связи с гиперболами Виллиса. Иногда мы, видимо, застаём процесс возникновения этого вторичного сходства. Скажем, сибирский Lanius cristatus и рыжехвостый жуланы L.isabellinus исходно достаточно сильно различаются, что видно при сопоставлении их «старых» подвидов. А вот молодая раса второй формы L.i.phoenicuroides вырабатывает признаки, сближающие её с L.c.cristatus, причём в условиях аллопатрии этих форм.
И заключительное соображение. По мере развития эволюционной теории мы видим, что её принципы имеют тенденцию подтверждаться. Скажем, что виды определяются не различиями, а обособленностью, и регуляторные механизмы в популяционных системах эту обособленность не просто поддерживают, а нацелены отбором именно на это. Или что обособление видов и их дифференциация по нишам – разные процессы, первый рождается из взаимодействий частей – географических популяций внутри популяционной системы материнского вида как целого.
Второй же определяется нишевой структурой сообщества, предзаданной вновь возникающим видам и пр.
А вот концепции относительно конкретных способов реализации этих принципов, господствовавшие во второй половине ХХ века, сейчас «падают». Это и «круги рас» с изолированными концевыми звеньями, и «усиление» с «совершенствованием изолирующих механизмов», и «правило Бейтмана» со следующей отсюда асимметричной моделью полового отбора, и исключительная ролью конкуренции в дивергенции близких видов и их дифференциации по нишам, и много чего ещё. Когда это обновление достигнет критической массы, откроется путь к новому синтезу – иной формулировке принципов, способной решить существующие противоречия СТЭ. Которые никуда не деваются.
Рекомендуем прочесть
 Теории биологической эволюции с позиций современного развития системной биологии
Теории биологической эволюции с позиций современного развития системной биологии Палеонтология и парадигмы современного естествознания
Палеонтология и парадигмы современного естествознания Перестройка в экологии: от описания видимого к пониманию скрытого
Перестройка в экологии: от описания видимого к пониманию скрытого Птицы и муравьи не мешают друг другу есть гусениц
Птицы и муравьи не мешают друг другу есть гусениц Холодный май — это потепление
Холодный май — это потепление Гиперболы Виллиса: попытка интерпретации
Гиперболы Виллиса: попытка интерпретации Обманщики чередуют ложь с правдой, чтобы им продолжали верить
Обманщики чередуют ложь с правдой, чтобы им продолжали верить Сезонные миграции копытных определяются культурой, а не генами
Сезонные миграции копытных определяются культурой, а не генами
