

Изменения гормонального состояния при урбанизации у птиц
Содержание
Прочитал интересный обзор Frances Bonier (2012) про изменения эндокринной системы в урбанизированных популяциях птиц (или у «диких» видов, оказывающихся в городе, вынужденных в нём существовать), который перескажу. Там приводятся данные также по рептилиям с млекопитающими, но сильно более отрывочные, чем по птицам, что не так интересно.
Как это ни удивительно (пишет автор во Введении), когда мы используем мелкомасштабную шкалу географических сравнений, то плотность человеческого населения и уровень биоразнообразия позитивно коррелируют друг с другом. Что говорит, с одной стороны, о том, что население нашего вида умело выбирает для поселения наиболее благоприятные места и в них увеличивается всего шибче. С другой – что теснимому нашими городами с постройками биоразнообразию ещё есть куда прятаться, а нам – есть чего охранять, не всё этими самыми городами вытеснено. В том числе охранять путём содействия возвратной урбанизации, тем более что для редких и уязвимых видов в староосвоенных регионах – не только в ближних пригородах «урбанистических ядер», но и подале — это единственный шанс уцелеть.
Также повышенный уровень стресса в городе связан не столько с конкретными вредными/пугающими факторами воздействия, сколько с общей неопределённостью ситуацией, нестабильностью и изменчивостью горсреды и, соответственно, существования особей в группировках. В том числе потому, что воздействия, направленно меняющие, скажем, суточные ритмы, фотопериодизм и сезонность размножения у птиц, вроде подавляющего воздействия городского освещения на выработку мелатонина, отвечающего за эти процессы, одновременно стрессированность не повышают. Тем более что за вычетом этой неопределённости/неустойчивости все прочие изменения условий существования в городе скорей к лучшему (для всех птиц, отдельные особи которых могут жить и размножаться в урбоместообитаниях). Корма там больше, хищников и специализированных конкурентов меньше, климат мягче, вообще абиотические условия благоприятней, почему продуктивность урбанизированных популяций выше, чем таковых в рекреационных лесах ближних пригородов – хотя демография городских популяций такова, что сильно большая часть особей отказывается от репродукции и/или практикует отложенное размножение.
То есть индивидам «дикого» вида в городе жить в принципе лучше и веселей, чем в естественных местообитаниях. Проблема в том, что надо жить популяциями – а вот «собрать» в нестабильных мозаиках городского ландшафта популяционную структуру, чтобы могла устойчиво воспроизводиться в череде поколений несмотря на три вида «помех» этому процессу – средовую стохастичность вокруг группировок, демографическую стохастичность – в самих группировках, и неопределённость, связанную с изменениями персонального состава соседних особей – задача вовсе не тривиальная.
Поэтому у большинства видов успешное размножение отдельных особей в городе или отдельных поселений не приводит к образованию популяций, к колонизации подходящих биотопов, расположенных буквально через улицу, по соседству, именно потому, что не у всех получается «поднять» устойчивость пространственно-этологической структуры популяций до уровня, превышающего нестабильность с изменчивостью и гетерогенностью городской среды. Понятно, что это стрессирует особей, поскольку устойчиво жить и размножаться они (а тем более их потомки) могут только в устойчивой популяционной структуре. Пока же она не сформирована, продукты успешного размножения городских особей уходят из города, пополняя собой «дикие» популяции, поскольку им надо встраиваться в устойчивую структуру, а в городе её ещё нет.
Поэтому стрессированность особей в городской популяции, находящейся в процессе формирования, пока идёт интенсивный обмен особей с пригородом, и значительная часть популяции пополняется притоком оттуда, и отток туда тоже значителен – она повышена. А вот когда урбанизация «дикого» вида завершилась, возникшая популяция демографически изолируется от пригородных почти полностью – и тогда уровень стресса снижается. Виды птиц по отношению к городу западные специалисты сейчас делят на urban avoider, urban adapter, and urban exploiter – соответственно, исчезающие из городов, прозябающие там (на уровне численности, меньшем, чем в естественных биотопах) и процветающие, умеющие эксплуатировать плюсы городских экосистем.
Возвратная урбанизация, о которой я писал ранее – это процесс, переводящий виды первой и второй категории в третью, запускается пребыванием региональной популяции вида «под прессом» растущего города в течение некоторого лаг-периода, если только за это время она вообще не вымрет.
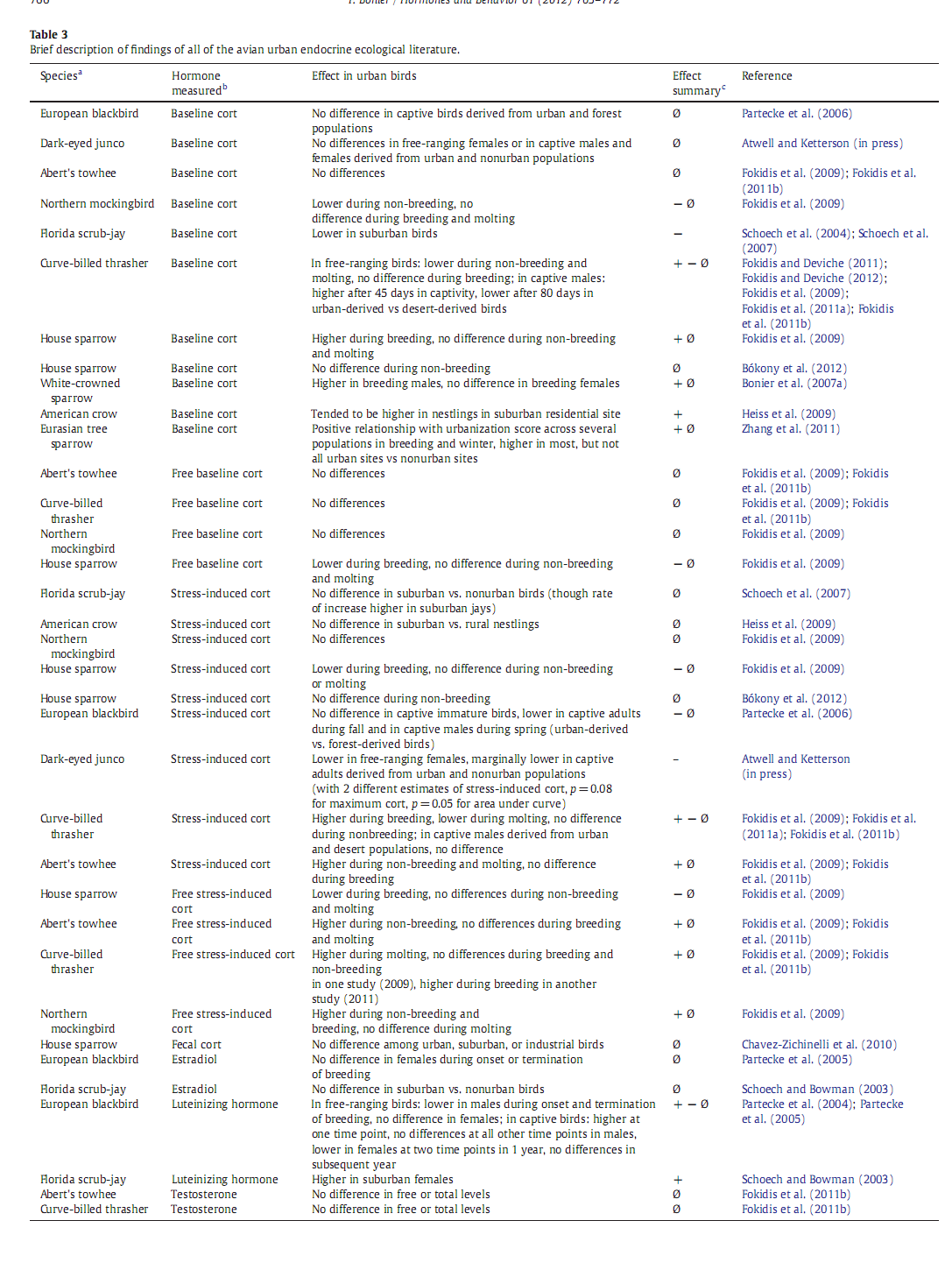
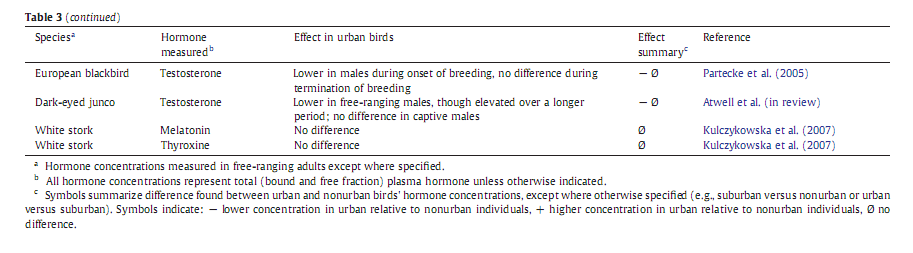
У позвоночных, в первую очередь у птиц, есть две главные «оси» управления эндокринной системой – гипоталамо – гипофизарно – надпочечниковая и гипоталамо-гипофизарно – гонадная (НРА и НРG). Они ответственны за поддержание энергетического баланса в условиях меняющихся потребностей метаболизма и обеспечения репродукции. В обоих «осях» зафиксированы изменения, связанные с урбанизацией. В большинстве исследований на этот счёт меряли концентрации гормонов – конечных продуктов первой и второй оси: гормона стресса кортикостерона и половых стероидов, тестостерона с эстрадиолом соответственно.
До сих пор исследовали либо, как особи разных видов в городе
а) отвечают на новые условия и/или выдерживают их, что связано в основном с НРА и
б) как меняется репродукция городских популяций, что связано в основном с НРG.
Сравнивали либо их уровни в городской популяции с негородскими для конкретного вида, либо (реже) смотрели их изменения для одного вида в разных городах по размеру, положению с сети городов, или (совсем редко) для разных видов птиц, вселяющихся в один и тот же город. Где предсказуемо выяснилось, что эндокринный ответ на урбанизацию у разных видов оказывается радикально различным, включая экологически близких и родственных. См.таблицу 2.
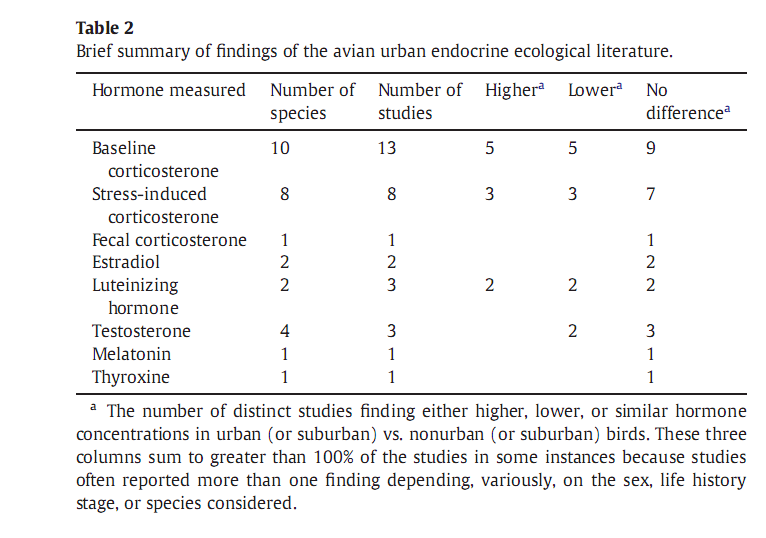 Действительно ли аллостатическая нагрузка меняется в процессе урбанизации?
Действительно ли аллостатическая нагрузка меняется в процессе урбанизации?
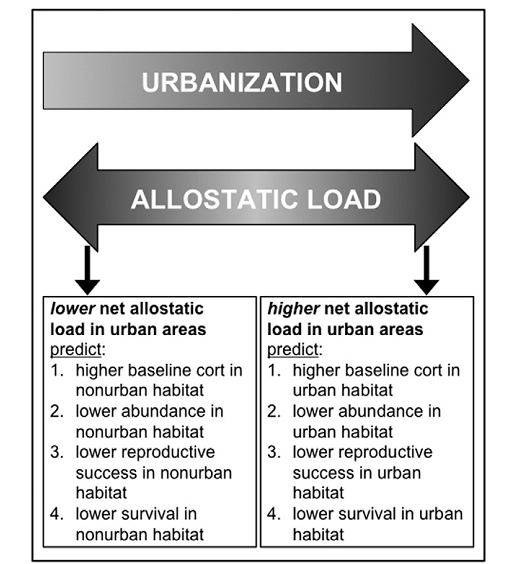
Аллостатическая нагрузка — это результат избыточной реакции на стресс, ведущий к преждевременному изнашиванию нашего организма. В городских популяциях разных видов она бывает выше, ниже или такой же. В первом случае численность и жизнеспособность городских популяций ниже чем в аналогичных местообитаниях вне города (также как репродуктивный успех с выживаемостью), во втором – наоборот (рис.1).
 В ряде случаев эндокринный ответ на факторы, обычно связанные с урбанизацией, исследовали за пределами городского контекста. Скажем, в выборке гнёзд воробьиной пустельги Falco sparverius и беловенечной зонотрихии Zonotrichia leucophrys изучали влияние разных факторов беспокойства, связанных с местом расположения гнезда на уровень кортизола – базовый и индуцированный стрессом. У птенцов первого он не менялся в зависимости от разных видов беспокойства (скорость движения и шум машин на автотрассе неподалёку, доля застройки в ландшафте вокруг). У птенцов второго, наоборот, обе концентрации были выше в гнёздах, расположенных рядом с загруженной трассой, чем в гнёздах, расположенных в отдалении.
В ряде случаев эндокринный ответ на факторы, обычно связанные с урбанизацией, исследовали за пределами городского контекста. Скажем, в выборке гнёзд воробьиной пустельги Falco sparverius и беловенечной зонотрихии Zonotrichia leucophrys изучали влияние разных факторов беспокойства, связанных с местом расположения гнезда на уровень кортизола – базовый и индуцированный стрессом. У птенцов первого он не менялся в зависимости от разных видов беспокойства (скорость движения и шум машин на автотрассе неподалёку, доля застройки в ландшафте вокруг). У птенцов второго, наоборот, обе концентрации были выше в гнёздах, расположенных рядом с загруженной трассой, чем в гнёздах, расположенных в отдалении.
 Fokidis с коллегами изучил изменения концентрации кортизола у птиц г.Феникс, а) вселившихся туда из пустыни вокруг и б) домового воробья –уже синантропного вида и расселяющегося по урболандшафту. Пятнистый кривоклювый пересмешник Toxostoma curvirostre – пустынный вид, образовавший процветающую урбопопуляцию – показывает значимо меньший уровень глюкокортикоидов в своей «природной» пустыне во внегнездовое время, которое, видимо, самый напряжённый сезон, в гнездовое же концентрации не отличаются. Важно подчеркнуть, что «негородские» домовые воробьи в данном случае населяли агроландшафт, тогда как прочие виды – пустыню, то есть существенно более экстремальный ландшафт. И осталось неясным, насколько влияли эти различия, а насколько – различия между городом и негородом.
Fokidis с коллегами изучил изменения концентрации кортизола у птиц г.Феникс, а) вселившихся туда из пустыни вокруг и б) домового воробья –уже синантропного вида и расселяющегося по урболандшафту. Пятнистый кривоклювый пересмешник Toxostoma curvirostre – пустынный вид, образовавший процветающую урбопопуляцию – показывает значимо меньший уровень глюкокортикоидов в своей «природной» пустыне во внегнездовое время, которое, видимо, самый напряжённый сезон, в гнездовое же концентрации не отличаются. Важно подчеркнуть, что «негородские» домовые воробьи в данном случае населяли агроландшафт, тогда как прочие виды – пустыню, то есть существенно более экстремальный ландшафт. И осталось неясным, насколько влияли эти различия, а насколько – различия между городом и негородом.
Есть только два исследования, в которых концентрации глякокортикоидов в городских популяциях сравнивались с негородскими для нескольких городов, различающихся степенью урбанизации. Это работа Zhang et al. (2011) по полевым воробьям из 7 разных мест внутри Пекина, существенно различающихся степенью урбанизации, разделённых дистанцией >3 км, и работа автора обзора с коллегами по Z.leucophrys из трёх 3-х городов на западном побережье США. В обоих случаях зафиксировано повышение концентрации глюкокортикостероидов относительно негородских популяций, тем большее, чем выше степень урбанизации, во втором случае – у самцов, но не у самок.
Напротив, у домовых воробьёв, исследованных зимой в разных точках «городского градиента» вокруг Будапешта, уровень глюкокортикоидов не варьировал в зависимости от степени урбанизации. Далее, популяции флоридской кустарниковой сойки Aphelocoma caerulescens из пригородов (suburbs) Флориды показывают меньший уровень глюкокортикоидов, чем особи из негородских популяций. Schoech et al., 2004, 2007 показывает, что это может быть связано с большей доступностью корма в пригородах. Эта работа уникальна, поскольку в отличие от других исследований гормонального отклика на урбанизацию авторы не только выдвинули предположение, какой средовой фактор за оклик ответствен, но и проверили его в опыте.
Оказывается, подстановка кормушек и гарантированность корма снижает уровень глюкокортикоидов в негородских популяциях, но всё-таки не до такого низкого уровня, как у птиц, живущих в пригороде. При этом дача сойкам дополнительного корма вполне отражалась на росте жировых запасах, содержании белка и пр., т.е. корм нормально усваивался. Видимо, важен ещё какой-то фактор.
Дальше рассматривается вопрос: если урбосреда вызывает «хронические» изменения гормонального состояния, то «острое» воздействие меняющихся условий при урбанизации вызывает такие же изменения НРА или нет? Было показано, что экспресс-изменения доступности корма и прямое воздействие средовых пертурбаций от непогоды, до неожиданных препятствий, особенно в случае столкновений с ними, шумы, и пр. сильно влияют на НРА-ось, что существенно в контексте урбанизации.
В большинстве сформированных городских популяциях особи формируют толерантность к тем стрессорам, на которые «дикие» популяции отвечают неофобией и тревожно-оборонительной реакцией. У них ниже дистанция вспугивания, ниже неофобия, больше смелость, в смысле готовность исследовать потенциально опасную, сложную и тем самым завлекательную ситуацию, включая взаимодействие и игру с составляющими её предметами компонентами, вместо того чтобы отступать от греха подальше. У «городских» птиц больше инноваций в плане необычных методов кормления, способов гнездования и пр. черт поведения с экологией, а также в поведении сильней выделена индивидуальность, притом что особи в популяции не более разнокачественны (скажем, в отношении приверженности альтеративным стратегиям поведения, или распределения по альтернативным психотипам, «координатное пространство» которых общее для всех позвоночных). Некоторые из этих поведенческих изменений точно связаны с тем, что в урбанизация «диких» видов меняет характер андренокортикальной реакции на стресс.
Отличается ли ответ городских птиц на острые стрессирующие воздействия?
Восемь работ сравнивают величины стресс-индуцированного уровня глюкокортикоидов у городских vs негородских популяций, согласно стандартному протоколу таких опытов (стресс причиняется поимкой и взятием в руки). И опять, никакого определённого паттерна найти не удалось (табл.2, 3). Я думаю, это связано с тем, что у них нет нормального критерия отличения «пассивных урбанистов», просто обитающих на «островах» своих привычных биотопов в городе, от успешно идущей урбанизации вида. Поскольку этот критерий должен строиться на сформированности популяционной структуры, устойчиво воспроизводящейся в мозаике городских местообитаний вопреки разнообразным стрессам и шумам, а не на успехе размножения/выживания индивидов, но я не видел западных работ в этой парадигме вообще.
Только две работы включают выдвижение и проверку гипотезы, как наблюдаемые гормональные изменения участвуют в адаптации вида к урбосреде, как возникли и зачем нужны. Jesce Partecke с соавторами исследовал гормональное состояние «городских» чёрных дроздов, давно уже сформировавших городские популяции, демографически обособленные от «лесных». Cейчас показаны и регулярные генетические различия, которые в работах самого Партекке найти не удалось. Птиц отлавливали в городских и «лесных» популяциях и выдерживали в вольере в саду, после чего исследовали стресс-индуцированные изменения уровня глюкокортикоидов. Предсказуемо, у городских особей он оказался ниже в «трудное время», осенью и весной.
Исключительно интересны исследования городской популяции тёмноглазого юнко, существующей в г.Сан-Диего совсем недолго – с 1983 года (все прочие популяции вида населяют горные сосновые леса, см. видео с диверсификацией юнко в Северной Америке, 7.02-7.17).
Там выявлены аналогичные сдвиги стресс-индуцированного подъёма глюкокортикоидов в связи с урбанизацией, что у дроздов. У свободноживущих «городских» самок он значимо ниже, чем у негородских; та же самая разница зафиксирована у самок, отловленных и выдерживаемых в одних и тех же условиях в вольере. Гормональные изменения скоррелированы с изменениями поведения. Смелость и развитость исследовательского поведения равно отрицательно коррелируют со стресс-индуцированным подъёмом глюкокорикоидов что у городских, что у негородских по происхождению птиц, выращиваемых в вольере, как и уменьшение дистанции вспугивания у свободноживущих птиц в городе. И у городских птиц в процессе урбанизации, с одной стороны, уменьшался подъём глюкокорикоидов в ответ на стресс, с другой, увеличивалась смелость, развитость исследовательского поведения и пр. Подытоживая эти данные, можно сказать, что смелость и чувствительность НРА к средовым стрессорам сопряженно меняются в ходе урбанизации вида, и концентрация глюкокортикоидов выступает важным посредником в этом процессе.
Также как с базовым уровнем глюкокортикоидов, работы Fokidis с соавт. по стресс-индуцированному уровню не имеют аналогов в литературе, настолько много всего и тщательно сделано. У городских пересмешников Toxostoma curvirostre уровень стресс-индуцированных кортикоидов варьирует в зависимости от стадии годового цикла. По сравнению с конспецификами, живущими в пустыне, он оказывается выше во время размножения, ниже во время линьки и сходен во внегнездовое время. Птицы из тех и других популяций, отловленные и выращенные в вольере, показывают сходный уровень стресс-индуцированных глюкокортикоидов.

Toxostoma curvirostre
Авторы исследовали также чувствительность и мощность НРА «городских» и «негородских» птиц, как эндокринной системы. Свободноживущие самцы сильней увеличивают уровень глюкокортикоидов в ответ на инъекцию высоких доз вазотоцин-аргинина и как высоких, так и низких доз адренокортикотропного гормона (АВТ), чем самцы из негородских популяций. Напротив, на инъекцию адренокортикотропного гормона (АКТГ) пересмешники из городских и пустынных популяций отвечают сходным образом. Данные этих полевых опытов показывают, что больший уровень глюкокортикоидов у городских птиц в период размножения может быть связан возрастающей чувствительностью системы: гипофиза – к АВТ, а надпочечников к АКТГ. Интересно, что городские и пустынные птицы равно снижают уровень глюкокортикоидов в ответ на инъекцию дексаметазона, синтетического антагониста рецепторов АКТГ.
Отличается ли НРG у городских птиц?
Урбанизация «диких» видов специфическим образом влияет на демографию городских популяций, которые почти всегда переходят к большей оседлости, большей доле неразмножающихся птиц или птиц с отложенным размножением на фоне увеличения среднеожидаемой продолжительности жизни в городской популяции, так что репродуктивный выход скорей увеличивается. А вот что происходит всегда – птицы начинают размножаться раньше, чем вне города, на фоне снижения сезонности размножения (стремящегося к большей круглогодичности). Что, ясное дело, не может не влиять на гормональный контроль размножения и/или опосредоваться изменениями в нём.
Однако данных об изменених в НРG в связи с урбанизацией ещё меньше, чем в случае с глюкокортикоидами, и какой-либо чёткий паттерн тут также не выявлен. Два исследования показывают сходный уровень эстрадиола у городских чёрных дроздов и «пригородных» флоридских кустарниковых соек с таковым во внегородских популяциях этих видов. У городских чёрных дроздов по сравнению с «лесными» ниже уровень тестостерона, но лишь в период активного функционирования гонад, а в период их регрессии зимой сходен. В недавно возникшей популяции юнко в Сан-Диего репродуктивный период экстремально расширился в сравнении с природными популяциями, и повышенный уровень тестостерона теперь поддерживается дольше. Однако и среднеустойчивый уровень этого гормона, и подъём, вызванный инъекцией гонадотропинов, у городских птиц оказываются ниже, чем в природных популяциях. Напротив, при клеточном содержании птицы, взятые из тех и других популяций, показывают сходные уровни тестостерона, т.е. различия в уровне наблюдаемые в поле – это пластичная и обратимая реакция на урбосреду, а не устойчивый сдвиг микроэволюционного характера. У тауи Аберта Pipilo Aberti и коричневого кривоклювого пересмешника концентрация тестостерона у размножающихся самцов была сходной в городских и природных популяциях.
Наконец, анализируя содержание лютеинизирующего гормона (ЛГ), мы видим, как отклик на урбанизацию зависит от пола. Свободноживущие самцы «городских» чёрных дроздов под Мюнхеном показывают меньший уровень ЛГ, чем «лесные», тогда как у самок уровень ЛГ близок в обоих случаях. Но когда птенцов из обоих популяций выращивают искусственно, наблюдается очень различный паттерн содержания ЛГ. «Городские» самцы показывали повышенный уровень ЛГ в начале периода быстрого развития гонад и сходный – на всех прочих этапах годового цикла, что повторилось и на следующий год. У самок дроздов уровень ЛГ «городских» птиц в этих условиях был ниже в 2 сезона, и сравнимым во все остальные, что повторилось и на следующий год. Т.е. при искусственном выращивании птиц из «городских» и «негородских» популяций прежде устойчивые различия стираются или делаются менее определёнными, т.е. они скорей связаны с пластичной реакцией на особенности урбосреды, чем обусловлены наследственными изменениями (в отличие от содержания глюкокортикоидов). Самки флоридской кустарниковой сойки из пригородов имели повышенную концентрацию ЛГ в сравнении с товарками из природных популяций.
На основании этих данных автор обзора выдвигает гипотезу, что именно реакция эндокринной системы делит виды, проникающие в урбосреду, на 3 вышеназванные группы с разной успешностью (Urban-Endocrine Hypothesis).
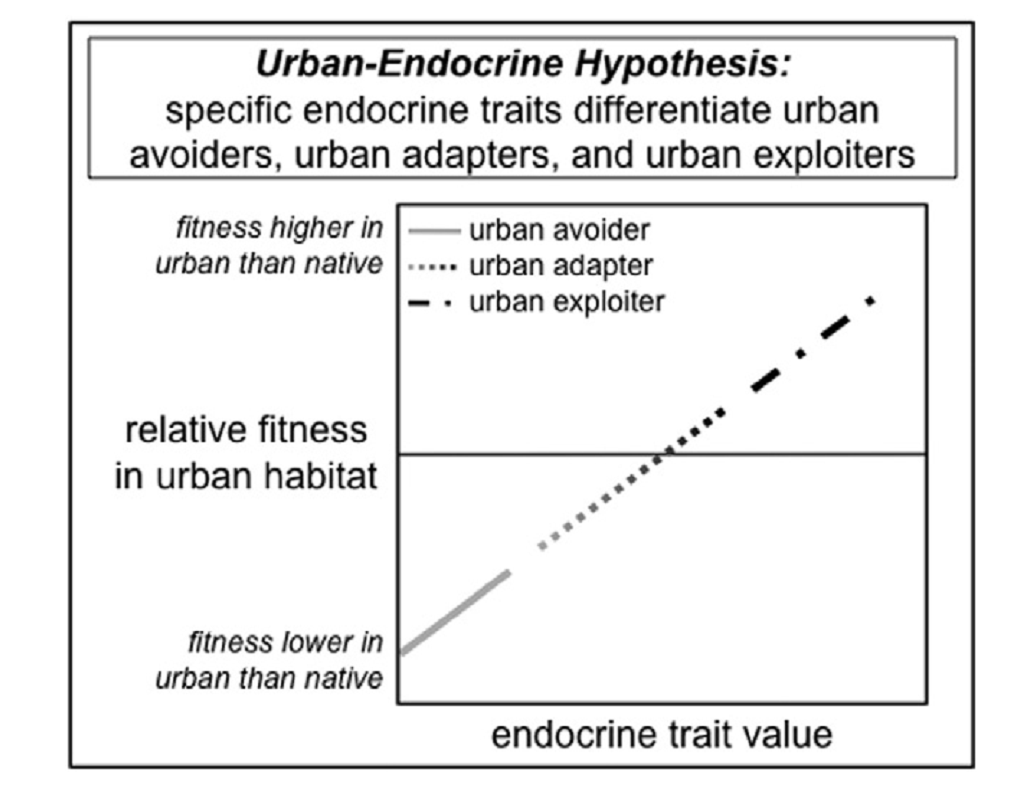
См.рисунок с предсказаниями Urban-Endocrine Hypothesis – что именно отклик эндокринной системы на специфику урбосреды «разводит» проникающие в город виды на пассивных урбанистов, терпящих урбосреду и эксплуатирующие урбосреду. Endocrine traits – характеристики эндокринной системы (НРА и НРG осей), такие как концентрация гормонов отклика, чувствительность системы, взаимодействие гормонов и поведения. Ось Х – признаки отклика эндокринной системы и одновременно её пластичность, позволяющая реализовать данный отклик в ответ на изменение условий (межиндивидуальная, наследуемая, или внутрииндивдуальная, модификационная). Ось У – итоговая приспособленность при жизни в городе, достигаемая «ценой» того «напряжения сил», которое откладывается по оси Х. Виды, расположенные «над» линией регрессии, оказываются преадаптированы к городу, быстро осваивают города и преуспевают в них, расположенные «под» — в лучшем случае «терпят», когда их местообитания «захвачены» растущим городом. Соответственно, микроэволюционные процессы, связанные с урбанизацией «диких» видов, состоят в переводе видовой популяции из второй категории в первую. Во многих случаях это случается не в самом городе, а на «архипелаге» местообитаний вида в регионе, испытывающей «давление» средового стресса, связанного с их последовательной фрагментацией растущим «урбанизированным ядром» (возвратная урбанизация).
По всей видимости, здесь особо значима НРА, реакция которой на острый стресс, связанный с существованием вида в урболандшафте, ответственна за сопряжённые изменения поведения в сторону большей толерантности к стрессорам, с одной стороны, и большей готовности к исследованию новизны, нежели бегства от неё – с другой. Первое утверждает автор обзора, второе ж добавил я сам на основании данных по доместикации, где в точности те же изменения поведения лисиц, норок и пр. связаны с теми же изменениями в НРА.
Урбанизация как доместикация
P.S. Судя по всему, сформированные урбанизированные популяции отличаются от «диких» ровно тем же, чем доместицированные лисицы, норки, крысы и пр. от соответствующих диких форм: резко ослабленной стресс-реакцией, особенно на новизну, повышенным «любопытством» с готовностью исследовать объекты, обычно пугающие — людей, технику, постоянные перестройки урболандшафта в пределах участка обитания. Опосредуется это гормональными изменениями, в виде общего ослабления стресс-реакции при «лучшем дозировании» её по реальной опасности. Так, «отбор лисиц на доместикацию замедляет темпы развития реакции страха и существенно отодвигает сроки ее первых проявлений. Действительно, у неселекционируемых лисиц уровень исследовательского поведения резко падает в возрасте 45 дней – именно тогда впервые проявляется реакция страха.
У ручных же этого не происходит даже в возрасте 4 месяцев. Интригует то, что падение исследовательской активности у щенков скоррелировано с подъемом в постнатальном онтогенезе уровня глюкокортикоидов в крови (Plyusnina et al., 1991). Вполне возможно, что замедление темпов развития нейрофизиологического субстрата реакции страха – это только одно из проявлений регуляторного действия генов, вовлекаемых в отбор на доместикацию».
В случае доместикации лис, крыс, норок и других зверей, исследованной в рамках школы Д.К.Беляева, отбор вёлся в двух противоположных направлениях – на большее миролюбие с толерантностью по отношению к человеку, и наоборот, на повышенную агрессивность. В обоих случаях отбор был быстр и успешен; что внутреннее состояние первых и вторых лис (крыс и пр.) сместилось именно в этом направлении, видно по появлению в вокализации новых звуковых сигналов, которых не было раньше.
Лучшими маркёрами того, что отбор ручных и/или агрессивных особей действительно меняет базовый уровень соответствующих мотивационных переменных [1] выступают изменения вокального репертуара, именно появление новых сигналов. Скажем, для ручных лисиц это кудахтанье/шумное дыхание, для агрессивных – фырканье/кашель.
Как это описано для сигналов-стимулов, данные инновации репертуара выражают новое внутреннее состояние животного вовне по правилам связи определённых характеристик вокализации с определёнными внутренними состояниями, присущим всем видам этого класса. Аналогичные изменения вокализации доместицированных форм в сравнении с дикими зафиксированы для крыс, морских свинок и других форм. В отличие например, от экспрессии генов мозга, где сравнение домашних собак с волками, домашних свиней с кабанами и домашних морских свинок с дикими не позволяет выявить какую-то одну группу генов, меняющих характер экспрессии во всех доместикационных событиях, все они наособицу.
Обратите внимание, что отбор действует не на «агрессивность вообще», а очень по разному в разном контексте. «Отбор на усиление агрессивности [крыс] по отношению к человеку не оказал существенного влияния на проявление межсамцовой агрессии. Необходимо отметить, что присутствие интрудера из агрессивной линии усиливало агрессивные ответы резидентов из ручной линии. Это указывает на то, что отбор на доместикацию серых крыс вызвал существенное ослабление межсамцовой агрессии и не вызвал изменений в основном агонистическом репертуаре (Plyusnina et al.,2011). Подтверждением сохранения высокой способности к проявлению агрессии у ручных крыс служат и данные по материнской агрессии. Ручные и агрессивные самки не отличались по уровню материнской агрессии и превосходили по основным ее показателям неселекционируемых самок. Можно предполагать, что длительное разведение ручных и агрессивных крыс в неволе привело к уменьшению порога материнской защитной агрессии (Konoshenko, Plyusnina, 2012) (тут, с.152).
То есть не только у человека, но и социальных животных специфика поведения больше определяется значимыми сигналами «извне» (включая произведённые/производимые им самим), чем влечениями «изнутри»).
Важно подчеркнуть, что миролюбивые лисы, норки, крысы и пр. отнюдь не стали менее агрессивны «вообще». Просто они, как и умные люди в сравнении с не очень, опытные боксёры в сравнении с неопытными драчунами, лучше классифицируют проблемные ситуации, куда попали, отличая должные от недолжных (для проявления соответствующего поведения), и точней дозируют необходимое количество усилий по сложности задачи. Соответственно, их агрессия и оборонительное поведение не растрачиваются впустую, вроде бегства от ухаживающего человека или атаки, если он подойдёт ближе, к нему они дружелюбны или равнодушно-толерантны. А вот на реальный вызов, скажем связанный с внутривидовой агрессией, реально затрагивающей статус животного, они (особенно «мирные» крысы) дают сильный и точной ответ, не уступающий таковым «агрессивной» линии и часто превосходящий «дикую».
Происходит такой рост адекватности в связи с гормональными изменениями, сопровождающими доместикацию. Во всех линиях уменьшается стрессируемость особей ситуациями, воспринимаемыми как проблемные, от приближения людей до агрессии другой особи, уменьшается концентрация главного гормона стресса, кортизола, высвобождаемого при этом. Происходит это благодаря изменениям в одной из 2-х главных эндокринных осей организма — гипоталамо-гипофизарно-надпочечниковой системы (вторая гипоталамо-гипофизарно-гонадная).
«При отборе лисиц на доместикацию происходили генетические изменения регуляторных систем онтогенеза, прежде всего – ослабление функциональной активности гипоталамо-гипофизарной надпочечниковой системы (ГГНС) (Трут и др., 1972; Оськина и др., 2008). Особенно важно отметить, что эффекты отбора по поведению на ГГНС прослеживаются на уровне не только фенотипических параметров активности этой системы, но также экспрессии ключевых генов системы, кодирующих кортикотропин-релизинг фактор, проопиомеланокортин и рецептор глюкокортикоидов (Gulevich et al., 2004; Оськина и др., 2008; Гербек и др., 2010).
Доместикация вовлекает в сферу своего действия не только гормональные, но и нейромедиаторные системы онтогенеза (Naumenko, Belyaev, 1980). В связи с проблемой доместикации особого внимания заслуживает серотониновая система мозга, участвующая в ингибировании агрессивного поведения. У доместицированных лисиц были выявлены более высокие уровень серотонина в мозге и активность ключевого фермента его синтеза (Popova, 2006). Интересно, что роль именно серотониновой медиаторной системы была неоднократно продемонстрирована в регуляции эмбриогенеза (Бузников, 1987; Cote et al., 2007).
Принципиальное значение имеет тот факт, что трансформация поведения лисиц в сторону домашнего в ходе эксперимента сопровождалась возникновением ряда фенотипических новшеств, которые имеются у собак и у других домашних животных. Эти новшества затрагивали, прежде всего, такой признак, как окраска мехового покрова: возникновение специфически локализованных депигментированных пятен («Star» фенотип) и светло-бурых подпаленных участков («Mottling» фенотип). Характерными морфологическими маркерами доместицируемых лисиц, так же, как и других домашних животных, являются вислоухость, «лаечный» (свернутый в кольцо или полукольцо) хвост, укорочение и расширение лицевого черепа; у отдельных животных наблюдалось также укорочение хвоста и лап (Trut, 1999; Трут, 2007; Trut et al., 2009)«. Аналогичные изменения гормонального статуса фиксируются при доместикации у птиц, например, у бенгальской амадины в сравнении с дикой острохвостой.
Дальше анализ поведенческих признаков, выполненный в цитированной выше работе, показывает, что у диких, агрессивных и мирных лис разнообразие соответствующего поведения (выступающего откликом на отбор) описывается с помощью двух главных компонент. Координаты по первой описывают, условно говоря, знак реакции на проблемную ситуацию, вроде приближения человека и иной потенциальной угрозы. Координаты по второй относятся к уровню активности — насколько животное бурно проявляет эту реакцию?
А лёгко «выстреливающая» стресс реакция с повышенным содержанием кортизола – неприятная штука. Конечно, сие обеспечивает мобилизацию при стрессе, но за счёт плохих телесных и когнитивных эффектов. Блокирует обучение, способствует «выученной беспомощности», всякие внутренние проблемы вызывает и пр. Поэтому уменьшение концентрации кортизола и ослабление индуцированной стресс-реакции при доместикации крайне способствует продлению периода обучения и всем плюсам, описанным выше.
Надо сказать, что не только у позвоночных, но и у людей устойчивый стресс и подъём уровня кортизола — характерный симптом на вынужденность существования в неблагоприятной среде, опасной, подавляющей, где человек постоянно сталкивается с пренебрежением, преследованием, нападениями и пр. Как удар плетью или кулаком оставляет следы на коже, пренебрежение к бедным, матерям-одиночкам или неграм в США, и т.д. при капитализме создаёт постоянный стресс для всех названных категорий жертв системы, который они не могут контролировать. Что ведёт и к разрушительным последствиям для телесной физиологии и/или душевной жизни, что к угнетению когнитивных способностей.
Подъём базового уровня кортизола с общим облегчением стресс-реакции равно у людей и животных тормозит обучение, сокращает «детский» чувствительный период, когда индивид учится легче всего и многому может выучиться, наиболее любознателен и открыт миру и пр. Так, подъём кортизола в крови представителей угнётённых групп ведёт к повышенным рискам рождения детей с низким и аномально низким весом новорожденного (<1500 г и <1000 г.). «Количество случаев рождения детей с низким и аномально низким весом в США снизилось за последние десятилетия, но остаётся ещё очень высоким [2]. В 1996 г. 7,4% новорожденных родились с весом ниже 2500 г – почти 290000 детей разных национальностей (Guyer et al., 1997). Около 19% из этих детей весили менее 1500 г.
В США случаи низкого веса значительно более распространены среди чернокожих, чем среди белых или латиноамериканцев. В 1996 году эти уровни составляли соответственно 13,0%, 6,3% и 6,2%. Интересно, что различия между белыми и чернокожими отсутствуют на Кубе (Hogue, Hargraves, 1993)».
Хелен Би, 2004. Развитие ребёнка. 9-е изд. СПб.: Питер. С.132.
И действительно, у сложившихся урбанизированных популяций птиц в сравнении с «дикими» наблюдаются этого же рода изменения. Общий уровень стресса устойчиво снижен, вместе с содержанием глюкокортикоидов, и с неофобией, в реакции на сложную среду и потенциально-опасные ситуации преобладает смелость с исследованием, а не отступление. Как показано на городской популяции юнко, существующей в Сан-Диего с 1983 (все прочие популяции – «дикие», существующие в горных сосновых лесах), эти различия сохраняются у особей тех и других популяций, выращенных в одинаковых вольерных условиях, также как сопряжение меньшей стрессируемости со смелостью и исследованием нового.
Однако в период формирования городских популяций уровень глюкокортикоидов скорей повышен пропорционально урбанизованности территории, осваиваемой «диким» видом, также как степень стрессируемости новыми условиями (обзор). Важно подчеркнуть, что у птиц общая стрессируемость и связанный с ней базовый уровень глюкокортикоидов – наследственный признак, устойчиво различающийся, скажем, у разных подвидов Melospiza georgiana при искусственном выращивании их представителей в одних и тех же условиях.
Иными словами, растущие города, скорее всего, «не отбирают» наиболее «смелые» и/или «умные» виды региональной орнитофауны, но их особи «делаются» таковыми в самом процессе урбанизации. Почему этот последний носит микроэволюционный характер, несмотря на неопределённость данных о морфологической дивергенции формирующихся городских популяций от исходных негородских.
Действительно, проверка идеи, что в процессе урбанизации диких видов млекопитающих преимущество получают «более умные» особи, поэтому
а) размер черепа в городских популяциях должен быть больше, чем в сельских,
б) размер черепа в городских популяциях должен расти со временем
— не подтвердила её. Смотрели черепа 10 видов мелких млекопитающих по коллекционным данным за последние 100 лет. Оказалось, что а) выполняется только для двух видов из этих 10, б) не выполняется вовсе — более того, у 2-х видов размер черепа в городских популяциях уменьшался. Далее, сельские популяции всех насекомоядных видов также показывают увеличение размеров черепа. Последнее, думаю, происходит в ответ на стресс, следующий из фрагментации их местообитаний разрастающимся «городским ядром», в то время как в городе завершившееся освоение урбосреды позволяет «миниатюризировать мозги», беря не размерами, а совершенством когнитивной конструкции. Опять же, как при доместикации.
Иными словами, «больший ум» городских популяций, их готовность исследовать сложную среду, играть с незнакомыми предметами, подробно описанные в книге Н.Н.Мешковой и Е.Ю.Федорович (1996) – побочное следствие снятия блока, связанного с высокой и лёгкой стрессируемостью, а не отбора на повышенные когнитивные способности per se. И действительно, когда мышей селектировали на бОльшие «когнитивные способности» — рост вероятности решения экстраполяционного теста, исходно не отличающейся от 0.5, сперва это не получилось из-за резкого возрастания роста тревожности в тестовой обстановке у отбираемых. Поэтому стали отбирать одновременно на лучшее решение теста и меньшую тревожность. Первое получилось не очень – тестовые результаты сперва поднялись над уровнем 0,5, но в 6-8м поколениях отбора снова упали – а второе получилось отлично. Причём в отличие от изменений агрессии, описанных выше, тревожность понизилась в широком спектре ситуаций — вместе с неофобией и в разных контекстах – отношение к новой пище, тест открытого поля (тут, с.46, 155). Видимо, именно этим объясняется подъём вероятности решений теста за пределы 0,5 в первых поколениях отбора, а не улучшение когнитивных способностей как таковых.
Из этой картины урбанизации «диких» видов следует один интересный момент. Смелость вкупе с готовностью исследовать незнакомую и потенциально опасную ситуацию вместо того, чтобы отступать от неё, развивая пассивно-оборонительные реакции — важная характеристика «лучших» особей в природных популяциях. Приверженцы конкурентной стратегии [3], они поддерживаются отбором не сами по себе (иначе б заполонили популяцию), а для устойчивого воспроизводства пространственно-этологической структуры последней. Там их поведение играет роль индуктора и организатора, управляющего активностью приверженцев противоположной патиентной стратегии. Благодаря ему паттерны активности «худших» особей «помещаются» в пространственные и временнЫе лакуны между активность «лучших», так что те и другие образуют скоординированную «карусель» или «хор». Одновременно каждая из альтернатив специализируется к условиям собственной «лакуны», которая одновременно и коммуникативная ниша. Что делает их ещё более комплементарными друг другу в плане состава поведения и взаимозависимыми в функциональных связях, укрепляет социум и пр. См. описание коммуникативной сети между самцами разного «качества» (или видов, занимающих разное место в межвидовой иерархии) у певчих птиц в работах P.McGregor с соавторами или В.В.Иваницкого.
То есть специфика процесса урбанизации «диких» видов состоит не в отборе наиболее умных особей в формирующейся городской популяции (или видов с более развитым мозгом/пластичным поведением в городскую среду), как часто считается. Нет, главное содержание урбанизации – микроэволюционное изменение, в ходе которого с основной массы «худших» особей, приверженных патиентной стратегии, снимается блок на развитие смелости, уменьшается неофобия, увеличивается если не способность к обучению, то готовность учиться и пр.
Точно также как в процессе доместикации, это происходит за счёт снижения реакции стресса, вызванного неопределённостью и новизной, с увеличением способности дозировать её сообразно реальной стрессогенности событий и объектов, с которыми столкнулось животное, без мощной добавки в связи с неофобией. Одновременно сокращается до минимума пассивно-оборонительная реакция в отношении просто новых и неизвестных объектов, падает дистанция вспугивания в отношении безоружных людей и медленно работающей техники, оставаясь значительной, скажем, для вооружённых людей или быстро приближающихся машин. В ряде случаев поведенческие изменения, уменьшающие беспокойство людьми, техникой, или гибель от неё, когда оказываются недостаточны, «тянут» за собой изменения в морфологии, как у белолобых ласточек Petrochelidon pyrrhonota.
Всё это не отменяет разделения на «лучших» и «худших» особей в ходе конкуренции уже в городской популяции, только идёт оно по другим основаниям, нежели в популяциях городских. В городе «лучшие» особи более управляемы средой и менее зависимы от социальных отношений в группировках, «худшие» — наоборот. Легко видеть, что в негородских популяциях картина противоположная. Первые тем легче оставляют данное поселение и переселяются в другие с целью увеличить репродуктивный успех (что обычно и удаётся). Напротив, вторые, максимально привязаны к данной группировке и данному составу соседей, будучи консервативны как территориально, так и в отношении уже поддерживаемых социальных связей.
Примечания
[1] У спокойной особи в состоянии готовности действовать, а не только что в ситуации, контекстуально подталкивающей к дружелюбию и/или агрессии.
[2] Из-за максимального % детей растущих в бедности из развитых стран, что есть плата за блюдение чистоты либерального капитализма, с минимумом социального государства.
[3] Про противоположность конкурентной и патиентной жизненных стратегий особей, как альтернативных форм реагирования на проблемные ситуации, в которых они «втягиваются» ввиду конкуренции друг с другом и/или воздействия среды, см.тут.








{kind=link}