
 Резюме. Когда читал лекцию по этологии на летней практике в Пущино, увидел вдруг, как неожиданно обнаруживается маскировочность окраски больших пёстрых дятлов — на вывешенных там фото 9й конкурса научной фотографии «Стихии науки», проводимого «Наукой и жизнью». А вот недавно маскировочность окраски дятлов была показана строго, и именно на кормовых деревьях — для трёх американских видов этих птиц. Дальше мы пересказываем работу об этом Monica L. Carlson et al. (2024).
Резюме. Когда читал лекцию по этологии на летней практике в Пущино, увидел вдруг, как неожиданно обнаруживается маскировочность окраски больших пёстрых дятлов — на вывешенных там фото 9й конкурса научной фотографии «Стихии науки», проводимого «Наукой и жизнью». А вот недавно маскировочность окраски дятлов была показана строго, и именно на кормовых деревьях — для трёх американских видов этих птиц. Дальше мы пересказываем работу об этом Monica L. Carlson et al. (2024).
Резюме авторское. Маскировочный характер окраски птиц часто предполагается, но лишь в немногих работах данное свойство измерено а) напрямую, б) на экологически значимых субстратах в) с «точки зрения» тех, кто стремится данных птиц обнаружить, например, хищников и т. д. приемников сигналов. Авторы изучали а)-в) для цветного узора [все же в основном чёрно-белого. Прим.публикатора] оперения трех видов дятлов, обычно обитающих в Нью-Джерси (США) — каролинского меланепреса Melanerpes carolinus, пушистоперого Dryobates pubescens и волосатого Leuconotopicus villosus дятлов: действительно ли на коре своих обычных кормовых деревьев они хуже видны хищным птицам, чем на случайно выбранных стволах в их среде обитания?
Сначала они изготовили стандартизированные фото коры деревьев, на стволах которых кормились данные виды, и фото контрольных субстратов — коры деревьев с проложенных там же трансект.
Далее на оба субстрата наложили стандартизированные фото самих дятлов (вид со спины) для оценки заметности их визуальных образов для зрения возможных хищников — воробьиной пустельги Falco sparverius и клинохвостого орла Aquila audax. Затем для каждой комбинации дятлов и субстратов количественно оценили 4 составляющие маскировочных свойств рисунка:
1) взаимное соответствие зернистости [в образах дятла и поверхности, на которой сидит, чем больше расхождение, тем птица заметней, и наоборот],
2) взаимное соответствие цветов окраски птицы и субстрата,
3) то же для яркости и
4) насколько расчленяюща окраска птиц.
1)-4) исследовали лишь для структуры рисунка, безотносительно к цвету его составляющих. Дальше определяли количественно, насколько точно окраска дятлов соответствует таковой «своего» и «контрольного» субстрата с точки зрения цветовых пятен «в глазах» названных хищных птиц. Выяснилось, что да: именно на обычных кормовых деревьях дятловая окраска оказывается маскировкой по параметрам 2), 3), и 4). Авторы предположили, что потребность в маскировке при кормлении [ответ на риск нападения хищника] вероятно [отчасти] формировала эволюцию дятловой окраски. Дятлы были хуже заметны именно на кормовых деревьях (более маскрировочная окраска по данным параметрам чем на наугад выбранных деревьях трансект). Это рождает предположение, что на эволюцию рисунка оперения дятлов влияла необходимость быть хуже видимыми именно при кормлении [Большая маскировочность окраски позволяет реже осматриваться при долблении, т. е. сократить непродуктивные траты времени при долблении и другом кормодобывании].
Введение
Содержание
- 1 Введение
- 2 Методы сбора данных о кормовых деревьев
- 3 Изображения дятлов
- 4 Обработка изображений
- 5 Восприятие хищников
- 6 Количественная оценка маскировочности окраски: 4 параметра
- 7 Выполненные анализы
- 7.1 Действительно ли дятлы хуже заметны хищнику на кормовых, чем на трансектных деревьях?
- 7.2 Различается ли маскировочность окраски изученных видов количественно и качественно?
- 7.3 Кому из хищников они менее заметны, пустельге или орлу (различающимся остротой зрения)?
- 7.4 Чем кормовые деревья отличаются от трансектных (т. е. случайно выбранных в лесу)?
- 8 Результаты
- 8.1 Криптическая окраска работает: дятлы, как правило, хуже видны на кормовых, нежели на обычных деревьях.
- 8.2 Дятлы, как правило, более криптичны «в глазах» хищника с менее острым зрением
- 8.3 Разные виды дятлов специализированы по характеру маскировочности окраски
- 8.4 Кормовые деревья значимо отличаются от деревьев трансект по таксономическому составу, жизненности и диаметру ствола (~возрасту)
- 8.5 Визуальные образы коры кормовых и трансектных деревьев различны
- 8.6 Криптичность дятлов на коре зависит в первую очередь от рода и размера/возраста дерева, а также от вида дятла
- 9 Обсуждение
Стремление избежать гибели от хищника, возможно, одно из сильнейших давлений отбора, формирующих фенотип животных. Маскировочная (криптическая) окраска — обычная стратегия уменьшения риска обнаружения и гибели. Она часто выражена в подборе фона или создании расчленяющего паттерна окраски, затрудняющих распознавание контуров жертвы (Cott, 1940; Cuthill et al., 2005; Stevens & Merilaita, 2009; Thayer, 1909). Почти всегда (кроме случая прозрачности, Johnsen, 2014), криптические свойства сильно зависят от визуальных характеристик фона, с одной стороны, и зрения того хищника, от которого маскируются, с другой (Endler, 1978).
Природные биотопы различаются цветом, узором и освещëнностью [поверхностей, на которых обитают животные]: незаметные в одних частях ареала (например, на коричневых пятнах подстилки из листьев), они бросаются в глаза в других (на ярко освещенной поверхности зеленых растений) (Robledo-Ospina et al., 2017; Stevens et al., 2015). Кроме того, маскировка «работает» лишь при взгляде с определенной дистанции (Barnett & Cuthill, 2014; Barnett et al., 2017a, 2017b, 2018a, 2018b; Rodríguez-Morales et al., 2018; Tullberg et al., 2005), при специфических позах потенциальных жертв (Kang et al., 2015; Stevens & Ruxton, 2019) или «в глазах» хищников с определенными особенностями зрения (Stevens, 2007; Stevens et al., 2017; Théry et al., 2005). Если коротко, корректное изучение маскировочной окраски животных требует изучить, как жертв видят хищники, обычно охотящиеся на них, причём на тех субстратах/в тех микростациях, где они кормятся, отдыхают, реализуют другие виды активности [а хищники их ищут и ловят].
Количественно охарактеризовать образ животного так, как он видится хищнику, очень долго было трудно (Stevens et al., 2007; Stoddard & Osorio, 2019), но сегодня доступны продвинутые инструменты для этого (Mason & Bowie, 2020; Stevens et al., 2007; Stoddard & Osorio, 2019; Troscianko & Stevens, 2015; van den Berg et al., 2020), проложившие путь ко все усложняющимся исследованиям маскировочной окраски животных. С их помощью работают с натурными изображениями или видеозаписями животных в естественной среде обитания.
Например, Кang et al. (2015) сняли бабочек на той коре, на которой они обычно отдыхают, и оценили видимость получившихся образов «с точки зрения» охотящихся на них птиц. Выяснилось, что положение тела жертвы [видимо, относительно складок корки] влияет на успех маскировки. Price et al. (2019) сравнили фото полиморфных европейских зелёных крабов Carcinus maenas в их предпочтительными микрообитаниях для анализа восприятия их образов хищными крабами. Здесь отдельные морфы используют разные стратегии маскировки в зависимости от природы субстрата.


Исследования маскировки in situ, включающие моделирование образа жертвы, возникающего «в глазах» вероятного хищника, были проведены и на некоторых видах птиц. Например, Gómez и др. (2018) анализировали образы гнезд морского зуйка Charadrius alexandrinus и шилоклювки Recurvirostra avosetta «в глазах» вероятных разорителей, выявив предпочтение их устраивать именно там [в тех микростациях], где яйца наименее заметны.

Кладка шилоклювки (с подброшенным яйцом ходулочника Himantropus himantropus, редкий случай факультативного гнездового паразитизма)
Troscianko et al. (2016b) по фото измеряли криптичность насиживающих козодоев, смолелировали, как птица видится вероятным хищникам. Выяснилось, что маскировочность образов насиживающей птицы положительно связана с выживаемостью кладок. Подобно двум этим работам, маскировочность птичьей окраски с учётом восприятия хищника изучали in situ в основном на гнездах и яйцах (Gömez et al., 2018; Langmore et al., 2009; Stevens et al., 2017; Stoddard et al., 2016; Troscianko et al., 2016) или на оперении наземногнездящихся птиц (Mason et al., 2023; Stevens et al., 2017; Troscianko et al., 2016b; Wilson-Aggarval et al., 2016). В первую очередь это связано с трудностями количественного анализа внешнего облика (все равно, с помощью ли калиброванных фотографий или спектрофотометрических данных) высоко подводных птиц на природных субстратах.
Одно из примечательных исключений — недавнее сравнение криптичности обликов взрослых обыкновенных пищух Certhia familiaris, пойманных паутинной сетью и обтянутых ею, на коре их гнездового дерева, ближайших деревьев и деревьев в более обширном пространстве вокруг, как они воспринимаются возможными хищниками (Nokelainen et al., 2022). Хотя не было найдено значимых различий между этими тремя ситуациями, выяснилась значимо большая криптичность оперения спины сравнительно с боками и брюхом, при меньшей заметности пищух для хищных млекопитающих, чем для птиц (Nokelainen et al., 2022). Для углубления понимания критичности окраски не-наземногнездящихся птиц авторы количественно определили её степень у 3х видов дятлов «в глазах» двух видов хищных птиц.
Отдельные наблюдения за кормящимися дятлами [или фото сидящих на стволах], позволяют предположить криптичность окраски их оперения (Bock & Larson, 1986; Short, 1982; Thayer, 1909). Больше того, широкомасштабный обзор видов семейства дятловых (Picidae) выявил взаимосвязь паттернов окраски оперения и среды обитания (Miller et al., 2019), что в принципе можно объяснить её маскировочными свойствами [Хотя вовсе не обязательно]. Наконец, поведение и экология многих дятлов требуют маскировочности окраски: большинство из них кормятся, лазая по стволам и ветвям деревьев, обследуют их, высматривая на поверхности, зондируя щели и/или раздалбливая корку в поисках артропод (Jackson et al., 2020; Jackson & Ouellet, 2020; Miller et al., 2020). Всё это времязатратно и требует отвлечения от осматриваний, снижающих риск неожиданного появления хищников вблизи птиц и их смертоносных атак.
Маскировочность обликов на подобных субстратах создаёт очевидные преимущества. Взрослых дятлов ловят хищные птицы, ястреба Accipiter sp. и сокола Falco sp. (Götmark & Post, 1996; Kettel et al., 2019; Ratcliffe, 2010; Rytkönen et al., 1998), создавая давление отбора на большую незаметность от этих весьма зорких хищников [это лишь предположение авторов, пока никем не доказанное — все эти хищники ловят дятлов нечасто]. Также показана способность дятлов, наряду с многими другими птицами, к сложному когнитивному поведению, особенно при кормодобывании (Cárdenas-Posada et al., 2023; Yi et al., 2014; Cárdenas-Posada et al., 2023; Yi et al., 2014).
Следовательно, вдобавок к развитию маскировочности окраски через элиминацию более заметных особей, отбор может поддерживать выбор птицами именно тех субстратов, на которых они незаметней всего. Авторы не различали оба пути развития маскировочности, но признают возможность и согласованное действие [и, главное, одновременное развитие] обоих. Они изучали, насколько маскировочна окраска трех североамериканских дятлов (каролинского меланепреса Melanerpes carolinus, пушистопëрого Dryobates pubescens и волосатого Leuconotopicus villosus дятлов) (рис.1) в их обычных местах кормодобывания «в глазах» двух видов хищных птиц.

Рисунок 1. Объекты исследования. Слева направо — каролинский меланепрес, пушистопёрый и волосатый дятлы. Показаны не в масштабе.
Исходя из предыдущих работ данного направления (Barbosa et al., 2008; Cheney et al., 2014; Chiao et al., 2009; Skrade& Dinsmore, 2013; Stevens et al., 2017; Stoddard et al., 2016; Troscianko et al., 2017; Troscianko et al., 2016b; Wilson-Aggarwal et al., 2016) анализировались фото птиц на коре в обычных местах кормления в сравнении с наугад взятыми деревьями. В количественной оценке степени критичности использовали вышеназванные параметры 1)-5). Они отражают соответствие между узором рисунка/»заливкой» его составляющих у фигуры (образа птицы) и узором/заливкой фона (субстрат). Предполагали, что чем сильней и разносторонней (по большему числу параметров) подобное соответствие, тем выше маскировочность окраски.
Действительно, в ряде работ-предшественниц (Skrade & Dinsmore, 2013; Troscianko et al., 2017; Troscianko et al., 2016b) показала связь между 1)-5) и выживаемостью жертв. Тем более данные поведенческих и нейробиологических экспериментов показали, что параметры 2), 3), 1) у птиц и других животных весьма значимы в ранней обработке воспринимаемого зрительным анализатором (Gaffney & Hodos, 2003; Ghim & Hodos, 2006; Lee et al., 1997; Mazer et al., 2002; Nguyen et al., 2004; Porciatti et al., 1989; Stoddard & Osorio, 2019; Van Hooser, 2007). Поэтому вполне вероятно, что они используются и зрением хищных птиц при распознавании образов потенциальной добычи [точней, обеспечивают нераспознавание её образа, маскируют его].
В дополнение к рабочей гипотезе о худшей заметности дятлов на их обычных кормовых деревьях, проверяли ещё одну — что деревья, выбранные для поиска пищи, отличаются от всех прочих в сторону меньшей заметности кормящихся там дятлов. Из предыдущих работ известно, что дятлы, как правило, кормятся в среднем на более крупных деревьях, имеющих, по крайней мере, некоторые признаки усыхания или стволовой гнили (Block, 1991; Conner, 1980; Conner et al., 1994; Dudley et al., 2012; Gunn & Hagan, 2000; Imbeau, 2002; Jackson, 1970, 1979; Kisiel, 1972; Nappi et al., 2015; Raley & Aubry, 2006; Williams, 1975). Оба параметра коррелируют с обилием потенциальных жертв дятлов (Conner et al., 1994; Jackson, 1979; Nappi et al., 2015). Поэтому авторы предположили, что дятлы предпочитают такого рода деревья, и именно здесь ниже заметность птиц данных видов.
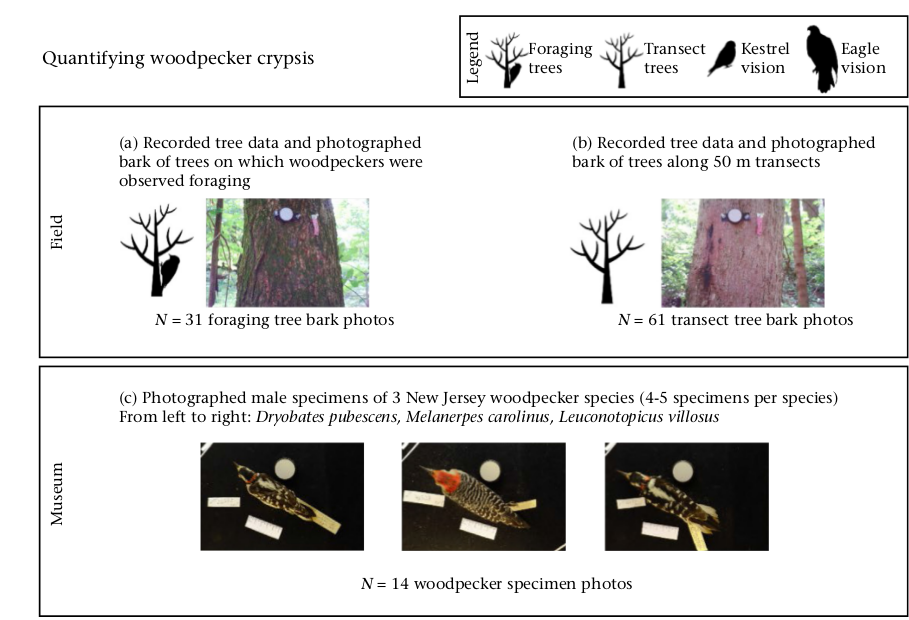

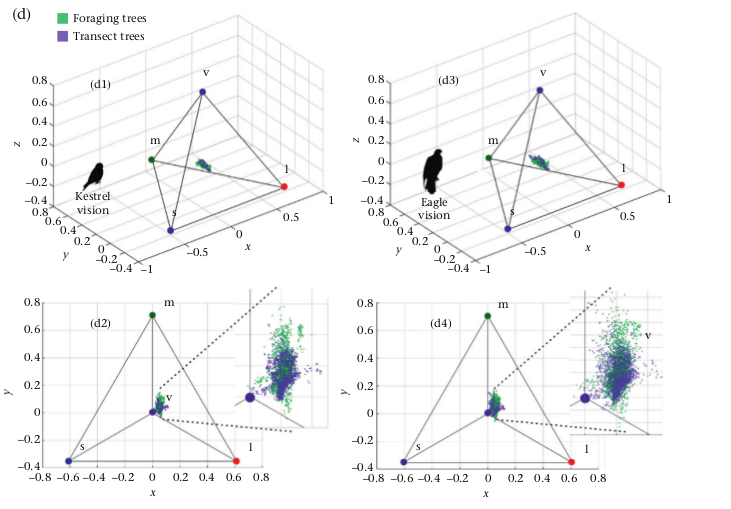
Рисунок 2. Краткий алгоритм исследования. Обозначения. Значки легенды слева направо: места кормления, просто деревья на трансекте сквозь тот же лес, как это видит пустельга, как это видит орёл. Последовательность действий: Верхний прямоугольник: Работа в поле. (а) Сбор данных о деревьях, на которых кормились дятлы, сфотографирование поверхности их коры, 31 фото. (b) сбор данных о деревьях вдоль 50-метровых трансектов в том же лесу, 61 фото. Средний прямоугольник — работа в музее. (с) Фотографирование тушек 3 названных видов дятлов Нью-джерси, по 4-5 экз. для каждого, самцов со спины, всего 14 фото. Нижний прямоугольник: визуальное моделирование и анализ данных.
(d). Получение образов 3х видов дятлов на своих обычных субстратах, как они видятся пустельге (в каждой паре изображений слева) и орлу (справа). (е). Количественное сопоставление образа птиц и субстрата по ахроматическим характеристикам — освещённости, контрасту и зернистости. (f). Численная оценка соответствий образов птиц и субстрата по цветам окраски («заливка» изображений), контуры тела птиц на субстрате показаны красным для первого и бурым — второго случая. (g). Оценка, насколько окраска образов дятлов, помещённых каждый на «свою» поверхность коры, является расчленяющей, с использованием фильтров Габора (Troscianko et al., 2017).
Методы сбора данных о кормовых деревьев
Краткий графический алгоритм методов исследования см. рис.2. Кормление дятлов наблюдали в июне-июле 2019 г. и в мае 2022 г. на территории Института углубленного изучения лесов и в [в расположенном с краю от него] резервате дикой природы им. Чарльза Роджерса, Принстон, Нью-Джерси, США. Кору в месте кормления удалось заснять у 18 каролинских, 7 волосатых, 3 пушистопëрых дятлов, 1 золотого Colaptes auratus и 1 хохлатой желны Dryocopus pileatus, всего 31 субстрат.
В конце концов авторы исключили золотого дятла с желной из исследований окраски, оставив места их кормления в базе данных. Дятлов считали кормящимися, если они осматривают субстрат, собирают членистоногих с поверхности коры и/или долбят его, но не когда барабанят на тех же стволах и сучках. Барабанная дробь — инструментальный сигнал, [функционально однотипный с песней воробьиных], обслуживает территориальность и/или привлечение партнера. Она хорошо отличима на слух от долбления при кормодобывании, поскольку громка, относительно быстра, имеет чаще всего равномерный темп (иногда ускоряющийся или замедляющийся, но всегда однотипный, это видовой сигнал). Напротив, долбление, связанное кормодобыванием, разнообразно и с неустойчивым темпом, [поскольку подчиняется особенностям субстрата], удары звучат тише и следуют медленней, менее регулярны (Short, 1982).
Авторы анализировали криптичность окраски только на кормовых деревьях, исключив те, на которых птицы барабанят или демонстрируют. Именно здесь дятлы снижают бдительность, направляя внимание на кору вместо осматриваний в поисках потенциальной опасности. Поэтому пресс хищничества и рожденное им давление отбора на более маскировочную окраску максимальны именно при кормлении. Напротив, когда дятлы барабанят, охраняя территории или ухаживая, им даже выгодно быть позаметней (Cárdenas-Posada & Fuxjager, 2022). Исследования на других видах птиц показали, что во время ухаживаний и другой социальной активности птицы держатся в местах, где особенно заметны (Uy & Endler, 2004), несмотря на большую возможность обнаружения хищником. Поэтому авторы исключили из анализа деревья, на которых дятлы барабанят, оставив лишь те, где кормятся.
Для каждого наблюдения (подробное объяснение, что что такое см. дополнение к методам) записывали координаты по GPS, затем характеризовали кормовое дерево, указывая его род (по возможности вид), обхват ствола на уровне груди (см), и жизненность по 4х балльной шкале (1 — живое, совсем не заметно мертвых ветвей, 2 — усыхающее, мертвых ветвей менее половины, 3 — почти усохшее (их большинство), 4 — сухостой, живые ветви отсутствуют. Затем камерой Sony A7, модифицированной для восприятия ультрафиолета, снимали поверхность коры на высоте ~1,2 м на протяжении ~40-60 см в длину. Камеру оснащали фильтрами, блокирующими или пропускающими УФ-излучение (Baader UV/IR 211 Cut/L-filter, 2 дюйма, также блокирующий ИК-диапазон vs Baader U-filter, 2 дюйма, CWL 350 нм). Это позволило получить изображение в видимом спектре (420-685 нм), и вслед за ним в ультрафиолете (320-380 нм).
Каждая фотография включала калибровочный образец 40% или 50% стандарта отражающей способности спектралона и масштабную линейку». [Т.е. попросту в экспозицию был включен кусочек пластика с известными характеристиками по коэффициенту отражения, рассеиванию света и спектральным характеристикам отражённого света. Используется для цветокалибровки и калибровки по яркости (экспокоррекция снимка). Фотографам более известно под названием «серая карта». Прим. Vinnie_sanders].
[При съемках субстратов] камеру устанавливали на штативе так, чтобы обеспечить равномерность освещения коры, как в тени, так и на солнце, во избежание неконтролируемого эффекта световых бликов. Делали по нескольку снимков с разной выдержкой, чтобы убедиться, что изображения не пере- либо недоэкспонированы (см. примеры изображений на см. рис.2а).
Также в июне-июле 2019 г., июле-августе 2020 г. описывали деревья, просто растущие на трансектах (их было 8 по 50 м каждый) там же, где и кормовые . Описывались все деревья, растущие в 2 м и менее от линии трансекта (дистанцию измеряли рулеткой, вытянутой под прямым углом к этой линии) и были крупней 30,48 см (12 дюймов) по окружности. Единицы этой выборки назвали «деревья трансект» чтобы отличить их от кормовых деревьев дятлов.
В таком случае некоторые из первых деревьев одновременно являются и вторыми. Их выборка включает кормовые деревья, а не противопоставляется им. Данные по ним, включая фото, собирались также, как по кормовым только не отмечали координаты каждого по GPS; вместо отмечали по GPS координаты начала и конца каждого трансекта.
Изображения дятлов
Взрослых самцов краснобрюхого (N=5), пушистоперого (N = 5) и волосатого дятлов (N= 4) засняли в коллекции Академии естественных наук Университета Дрекселя (список образцов см. приложение, таблица A1). Предшествующие работы такого рода показывают, что 4-5 экз. /вид вполне достаточно (Barreira et al., 2016; Carlson & Stoddard, 2024; Gomez & Th?ery, 2004; G€otmark & Hohlf€alt, 1995; Marcondes & Brumfield, 2019; Shultz & Burns, 2013). Больше того, качественно продемонстрировано, что по разным аспектам маскировочности окраски особи одного вида, как правило, более сходны друг с другом, чем с птицами других видов (приложение, рис. А4).
Хохлатую желну и золотого дятла не снимали по двум причинам: их было мало, меньше 3х, они слишком крупны для съемок на фоне коры тех стволов где кормились, не наложить соответствующие изображения. Однако фото их кормовых субстратов сохранены в наборе данных, поскольку на тех же деревьях живут потенциальные жертвы для видов, включённых в анализ (Maxson & Maxson, 1981). Тушки дятлов [для съемки] размещали на поглощающем свет черном фоне, окруженном рассеивателем света из политетрафторэтиленового пластика (PTFE), равномерно пропускающим и отражающим свет в УФ- и видимом спектрах.
Последний располагали так, чтобы весь приходящий свет либо прошел через него, либо отразился на образце, выровняв его освещенность. Источник света — купленная авторами лампа для рептилий (Exoterra Sunray reptile lamp, мощностью 50 Вт), размещенная на расстоянии ~45 см от образца. На каждом фото использовали серую карту и шкалу масштабирования.
Дятлов снимали цифровой камерой Nikon D7000 с объективом Nikkor 105 мм по методам из работ-предшествннниц (Stevens et al., 2007; Troscianko & Stevens, 2015). Камеру модифицировали для обнаружения УФ-излучения, удалив УФ-фильтр с датчика освещенности. Авторы последовательно использовали УФ- и ИК-фильтры (Baader UV/IR 211 Cut/ L-filter, 2-дюймовый) и УФ-фильтр (Baader U-filter, 2-дюймовый, CWL 350 нм) для получения изображения в видимом человеком спектре (420-685 нм), а затем снимали тот же образец в УФ- спектре (320-380 нм). Камеру установили на штативе на высоте ~1,15 м над образцом, для каждого вида дятлов делали несколько снимков с разной выдержкой, чтобы изображения не были переэкспонированы или недоэкспонированы (см. пример фотографий на рис. 2b).
Обработка изображений
Вся обработка изображений, количественная оценка маскировочности окраски, [и её численный] анализ выполнены в среде MATLAB (Natick, Массачусетс, США) версии 2017a, если не указано иное. Фотографии проверяли вручную на предмет переэкспонирования с помощью RawTherapee версии 5.8; фото коры или птиц с переэкспонированными пикселями исключены из анализа.
Каждая пара снимков (сделанных соответственно в видимом для человека и УФ-диапазонах) выровнена с помощью встроенной функции MAT-LAB. Затем на каждом фото измерили коэффициенты отражения разных пикселей и длину шкалы в пикселях. Далее фото нормализовали по значениям «своего» стандарта отражения и уменьшили до самой “уменьшенной” фотографии (имевшей минимум пикселей на мм из всех, Troscianko & Stevens, 2015,т. е. 8,486 пикселей на мм).
Для каждого фото коры вручную выбрали место, где помещать изображения дятлов, для гарантии, что образ дятла наложен на саму кору и не перекроется с фоном, стандартом отражения или шкалой. Каждое фото повёрнуто так, чтобы вертикально ориентировать переднезаднюю ось птицы относительно дерева, имитируя обычную посадку кормящихся дятлов.
Далее контур дятлов выделяли вручную, наложив на фон черную маску (с нулевым значением пикселя), и на основе таких силуэтов сделали два типа изображений для количественной оценки криптичности окраски птиц. Первый — изображение самой коры, ограниченное наложенным силуэтом птицы; его делали для каждой комбинации вид дятлов/его субстрат (рис.2ef) и сравнили с также обрезанным изображением дятлов для анализа соответствия фону (см. ниже количественную оценку криптичности).
Данная техника позволяет сравнить две области точно соответствующие друг другу по размеру и форме. Контроль размера и формы [сравниваемых] образцов значим, поскольку на измерения структуры рисунка (особенно зернистости), производимые с помощью двумерного быстрого преобразования Фурье (2D FFT) (см. количественную оценку криптичности ниже), влияют размер и форма таких рисунков.
Второй тип изображений создан наложением каждого обрезанного образа дятла на «свой» образ коры кормового дерева, чтобы анализировать его видимость хищнику на своём субстрате во всех комбинациях тех и других (рис.2d). Их использовали для оценок, насколько окраска птиц расчленяюща на «своём» субстрате (см. ниже количественную оценку криптичности). Изображения, где анализируемый образ животного наложен на субстрат, иногда называют “химерными”: ими пользовались в предыдущих исследованиях маскирововчных свойств окраски (Gömez et al., 2018).
Восприятие хищников
Для каждого из 3х типов образов, коры, дятла и химерного, реконстрировали как их видят «глаза» потенциального хищника. Использовали модель спектральной чувствительности его зрения (чувствительного к фиолетовому цвету, VS, как у ястребов и соколов). На ее основе значения пикселей, составляющих фото, преобразовывали в значения улавливания фотонов двухконусным фоторецептором в случае ахроматического анализа или в значения улавливания фотонов 4мя цветовыми рецепторами (длинноволновых LW, средневолновых MW, коротковолновых SW и чувствительных к фиолетовому VS) в случае хроматического анализа (Hart, 2002; Troscianko & Stevens, 2015). Авторы выбрали визуальную систему VS, присущую большинству хищнных птиц, охотящихся на взрослых дятлов (Odeen & Håstad, 2003). Увы, кривые спектральной чувствительности ястребов и соколов были недоступны, поэтому использовали таковые индийского павлина Pavo cristatus (Hart, 2002; Troscianko & Stevens, 2015).
Для ахроматического анализа использовали значения улавливания фотонов двойными колбочками, реагирующими на ахроматические аспекты сцены, вроде яркости (Jones & Osorio, 2004; Osorio et al., 1999; Osorio & Vorobyev, 2005; von Campenhausen & Kirschfeld, 1998). Для преобразования величин улавливания фотонов камерой в улавливание их каждым из типов фоторецепторов павлина использовали функцию отображения (Troscianko & Stevens, 2015), специально разработанную для того типа камеры и объектива, что использован авторами (Troscianko & Stevens, 2015).
Чтобы прикинуть детализацию, с которой хищник может рассматривать на коре кормящихся дятлов, каждое изображение преобразовывали, используя программу AcuityView (Caves & Johnsen, 2017), реализованную в MATLAB. Она преобразует вводимое изображение с учётом остроты зрения смотрящего и дистанции, на которой тот находится, а дальше преобразует изображение с помощью 2D FFT (быстрое двумерное преобразование Фурье) так что оно даёт наилучшее приближение к образу, воспринимаемому данным зрителем с данного расстояния (Caves & Johnsen, 2017).
Сокола — известные хищники дятлов [??? Непонятно, откуда авторы это взяли, это не так, но даже в той степени, в какой это так, речь не идёт о пустельгах], поэтому в первом моделировании восприятия хищником использовали данные об остроте зрения воробьиной пустельги. У неё острота зрения 1/40 (минимальный угол разрешения MRA). Авторы не нашли публикаций об остроте зрения ястребов р. Accipiter, и во втором моделировании восприятия дятлов хищниками использовали данные об остроте зрения клинохвостого орла Aquila audax (1/140 MRA) из Австралии (Reymond, 1985), принадлежащего к тому же семейству.
Дистанция от зрителя до объекта в обоих случаях 10 м. Выбор именно этого расстояния основан на том, что люди с их остротой зрения ~ 1/100 МРА) обнаруживали в лесу напечатанные на 3D-принтере модели пушистоперого дятла (самого мелкого из трёх) с ~14 м (Kawalec, 2021). Примеры изображений дятлов с поправкой на остроту зрения приведены в приложении, рис. А1.
Количественная оценка маскировочности окраски: 4 параметра
Для каждой комбинации вида дятла/кормового субстрата авторы определили 1) взаимное соответствие фигуры и фона и 2) насколько расчленяюща окраска у птиц. 1) измеряли 4мя способами: взаимное соответствие а) контрасту, б) яркости, в) зернистости и г) цвету.
А) показывает, насколько фигура соответствует фону с точки зрения диапазона отображаемых ахроматических оттенков. Если она полностью серая (контрастность низка) а фон в черно-белую полоску (контрастность высока), а) низко. Для его измерения авторы сравнили контрастность изображений дятлов vs коры по захвату светового потока двойными колбочками птиц [отвечающими за ахроматическое зрение] в каждом пикселе этого образа, с поправкой на остроту зрения хищника в каждом из случаев.
Как уже говорили, тестовые изображения птиц демонстрировали на чёрной поверхности (захват света 0); ненулевой захват света был только в пикселях, образующих фигуру, не фон (рис.2е). Тем же способом демонстрировались изображения коры, с которыми сравнивали образы птиц: их размеры и форма (значит, и число пикселей) в обоих случаях одинаковы (рис.2е). Контрастность рассчитывали как стандартное отклонение величин захватов светового потока в разных пикселях, образующих изображение (захвата двойными колбочками потенциального хищника, «глазами которого» рассматривается изображение, Cheney et al., 2014; Troscianko et al., 2016b). Контрастность образов птиц vs кормовых субстратов считали как абсолютную величину разности их величин, взятую со знаком минус (Troscianko et al., 2016b) по формуле (1)
Б) показывает степень соответствия фигуры и фона с точки зрения общей яркости, измеренной средней величиной яркости разных пикселей, образующих тот и другой (Luttrell et al., 2015). Скажем, черная (яркость низка) фигура на белом (высока) субстрате имеет низкое б), темно-серая (средняя яркость) на светло-сером (тоже средняя яркостью) — высокое (=большее соответствие). Численно яркость оценивается средним значением по всем пикселям изображения, поэтому черно-бело-полосатая фигура и серый фон могут идеально соответствовать друг другу по яркости, но различаться контрастностью.
Б) измерили также величиной светового потока, захватываемого двойными колбочками хищника, «чьими глазами» смотрят на образы дятла vs субстрата. Дальше сравнили измерения яркости по всём пикселям обоих изображений (с поправкой в обоих случаях на остроту зрения) как разницу сумм яркости всех пикселей, образующих изображение дятла, и пикселей, создающих образ коры, в обоих случаях деленное на число пикселей.
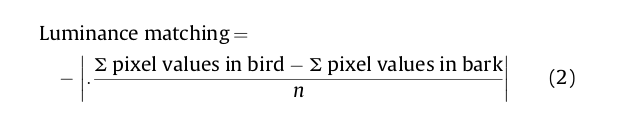
В) рассчитано как сопоставление зернистостей образа птицы vs образа фона, показывает насколько первый соответствует второму или контрастирует с ним. Скажем, при пятнах с горошину (зернистость повышена) объект сильней выделяется на фоне с пятнистостью величиной с яблоко (где зернистость понижена). Для измерения в) использовали 2D-FFT светового потока, захваченного двойными колбочками хищника, смотрящего на образ дятла, и сравнивали его с также преобразованным образом коры, с поправкой в обоих случаях на остроту зрения (Barbosa et al., 2008; Chiao et al., 2009; Godfrey et al., 1987; Troscianko et al., 2016b). Использовали те же изображения, что в анализах а) и б), лишь добавили к ним заполнение, пиксели со значением 0, чтобы вписать оба сопоставляемых образа в одинаковые квадраты.
С помощью 2D-FFT исходное изображение сначала преобразовали в образ, представляющий величины преобразования Фурье (его называли FFT-образ; каждый его пиксель представляет собой синусоидальную функцию пикселей исходного изображения, имеющую определенную частоту и ориентацию, Rzeszotarski et al., 1983; рассмотрено в Stoddard & Osorio, 2019). Частота синусоиды кодируется расстоянием каждого пикселя от центрального пикселя FFT-образа, а ориентация — его угловым расположением в пространстве, представляющем кору, и измерили объем, получившийся из перекрывания между этими гиперпространствами (Stoddard & Stevens, 2011), используя функцию R «voloverlap» в пакете R «pavo» (Maia et al., 2019) (рис. 2f). Соответствие зернистости рассчитали по формуле (3):

[Смысл этой процедуры пояснил разбирающийся в таких вещах т. Vinnie Sanders:
Мне не ясна конкретная математика вычислений (особенно, как получали стандартное отклонение значений пикселей для изображений энергетического спектра после полосового фильтра). Но по смыслу — каждое заранее подготовленное для сравнения изображение пары окраса дятла и рисунок коры дерева несколько раз обработали полосовым фильтром на быстром преобразовании Фурье (присутствует краткое, в общих чертах, описание алгоритма работы фильтра). Размерность полосы фильтру задавали от 2 пикселей с шагом степень двойки до максимально возможного для изображения.
Я не знаю, какие именно преобразования в матлабе они делали, но, по-моему, результат был аналогичный обычным полосовым фильтрам в графических редакторах — когда на изображении остаются лишь элементы соответствующей размерности (заданной параметром фильтру в пикселях), отображённые светлым на сером (50% яркости) фоне. В обработке изображений используется для создания масок, позволяющих выделить элементы определённого размера: например, чтобы выполнить повышение резкости только для волос. Или только для пор кожи — подчёркивая её текстуру, не затрагивая остальные элементы изображения, иной размерности.
Для полученных наборов изображений рассчитали стандартное отклонение значений пикселей. Из рассчитанных стандартных отклонений значений пикселей для преобразованных с помощью полосового фильтра изображений дятлов, вычли соответствующее по параметру полосового фильтра стандартное отклонение значений пикселей для пропущенных через полосовый фильтр изображений коры. Полученные разницы просуммировали по модулю для всех значений параметра полосы фильтра и результат умножили на -1. Если я правильно понимаю, при идеальном совпадении сравниваемых рисунков полученный параметр был бы равен 0].
Г) Соответствие цветностей — характеристика, показывающая, как точно фигура соответствует фону по цвету. Для измерения (Г) авторы случайно выбрали по 500 точек в образах дятла vs коры с поправкой на остроту зрения. Затем рассчитали, как это выглядит «в глазах» хищника, определив длины волн фотонов, уловленных его колбочками (4 классов — длинно- L, средне- М, коротко- S и фиолетовыми VS) при смотрении на эти точки, и разместив значения уловленного в 4хмерном пространстве воспринимаемых им цветов (тетраэдр, каждая вершина которого «представляет» один из цветов — и, соответственно, колобочек (Endler & Mielke, 2005; Goldsmith, 1990; Stoddard & Prum, 2008) (рис. 2f).
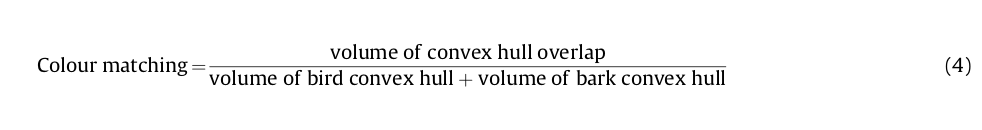 Г) получали как меру пересечения этих гиперобъёмов для образов дятлов vs образы коры. Пперед исследованием те и другие также заключали в квадраты, как описано выше.
Г) получали как меру пересечения этих гиперобъёмов для образов дятлов vs образы коры. Пперед исследованием те и другие также заключали в квадраты, как описано выше.
В измерениях характеристик А)-В) 0 указывает на идеальное соответствие фигуры и фона (т. е. максимум маскировочности), тогда как всё меньшие (отрицательные) — на худшее совпадение. В измерении Г) наоборот, большие значения = лучшее соответствие (и большая маскирововочность).
Д) — степень дизруптивности окраски — показывает, насколько видимость фигуры понижена (и сама фигура = цель хищника менее заметна) из-за взаимодействия между её собственным узором и узором фона, действительно ли эти узоры близки настолько, что затрудняют выделение образов дятлов? Д) измеряли по алгоритму, использованному в Troscianko et al. (2017): он применил фильтры Габора для количественной оценки степени, в которой узор внутри фигуры препятствует её выделению на определённом фоне (или, енаоборот, помогает последнему).
Фильтры Габора эффективны для выделения и количественной оценки сложных узоров, «изображений со структурной избыточностью, имеющих квазипериодическую структуру. К ним относятся дактилоскопические изображения, изображения кристаллограмм и интерферрограмм. В живой природе подобный окрас нередко встречается у зебр, различных кошачьих (тигры, рыси, дымчатые коты), птиц (тетерева), рыб (лепоринус полосатый) и прочих представителей флоры и фауны» (Daugman, 1988; Gabor, 1946; Stoddard & Osorio, 2019). Хотя есть и другие способы оценки дизруптивности окраски, авторы использовали этот (GabRat), поскольку Troscianko et al. (2017) показали, что оценки, полученные с его помощью — лучший предиктор времени, проходящего до обнаружения виртуальной цели в соответствующих задачах, и большая мера дизруптивности коррелирует с более долгим поиском (т.е.худшей видимостью) этих целей.
Для определения Г) использовали изображения птиц, наложенные на кору, а также изображения-контуры птиц, состовшие только из одного пикселя. Для моделирования их видимости «глазами» пустельги и орла мы использовали фильтры Габора с пространственной частотой 10 циклов/градус, сообразно пикам контрастной чувствительности (PCS), измеренной у воробьиной пустельги (Hirsch, 1982) и клинохвостого орла (Reymond & Wolfe, 1981). Для каждого образа дятла на коре Г) оценивали как среднее всех значений расчленяющего эффекта в каждом из пикселей (Troscianko et al., 2017), по формуле (5), где n — количество точек в контуре:
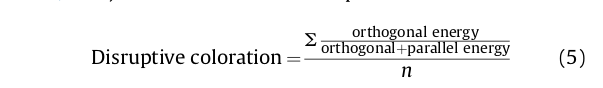
Чем выше Г) (более расчленяющий характер окраски), тем менее заметной на коре птица должна быть в принципе.
Выполненные анализы
Действительно ли дятлы хуже заметны хищнику на кормовых, чем на трансектных деревьях?
Авторы рассчитали средние значения каждого из 5 параметров маскировочности окраски А)-Д) сперва для каждого вида (по всем особям выборки), а затем и в среднем для дятлов, усреднив значения по 3м видам. Они это делали и для определения маскирововочности окраски «дятла вообще», и из-за небольшого размера выборок по отдельным видам (а так он возрастает). Поскольку величина выборок для кормовых и трансектных деревьев различна, в их сравнении использовали Welch’s t-tests (вариант t-критерия Стьюдена для независимых выборок).
Различается ли маскировочность окраски изученных видов количественно и качественно?
Чтобы исследовать межвидовые различия дятлов по маскировочности окраски «в глазах» хищника, авторы сравнили у них 5 аспектов последней А)-Д) на кормовых и трансектных деревьях с помощью t-теста Уэлча. Затем выполнили однофакторный дисперсионный анализ, где действующим фактором были виды дятлов, а откликом — 5 вышеназванных аспектов маскировочности окраски, чтобы определить, насколько они детерминированы видом дятла.
Затем оценили значимость различий между видовыми значениями параметров А)-Д), используя ранговый коэффициент Тьюки для определения значимости различий между групповыми средними, вариант множественных сравнений (только для кормовых деревьев). Каждый из этих анализов выполнен дважды, для случаев пустельги и орла.
Кому из хищников они менее заметны, пустельге или орлу (различающимся остротой зрения)?
Для ответа на этот вопрос авторы сравнили по t-критерию Уэлча характеристики маскировочности А)-Д), усредненные сперва внутри каждого вида, а затем и по всем 3м видам, как они видятся пустельге и орлу. Здесь использовали лишь данные по кормовым деревьям.
Чем кормовые деревья отличаются от трансектных (т. е. случайно выбранных в лесу)?
Авторы сравнили распределения кормовых и трансектных деревьев по составу родов, баллам жизненности и возрасту (~размеру) с использованием критерия χ2; по окружности ствола на уровне груди их сравнили с использованием t-критерия Уэлча. Авторы также сравнивали образы коры кормовых vs трансектных деревьев в «глазах» пустельги и орла по степени контрастности, яркости и цветности, чтобы лучше понять, как эти характеристики фона могут быть связаны с данными о маскировочности окраски птиц (обратите внимание, что сам по себе этот анализ не имеет отношения к изучению маскировочности окраски птиц).
Для измерения контраста и яркости коры «в глазах» хищника использовали данные по захвату фотонов разной длины волны колбочкками VS. Первый оценивали стандартным отклонением значений для пикселей, составляющих образец коры, яркость — средним значением для тех же пикселей. Сравнение деревьев по контрасту и яркости проводили по t-критерию Уэлча.
Для сравнения коры разных деревьев по цветности случайным образом выбрали 500 точек в каждом из образцов, и дальше анализировали их размещение в тетраэдрическом цветовом пространстве также, как в случае окраски птиц. Все изображения коры были одного размера и формы, поэтому содержали одно число пикселей.
Дальше исследовали влияние названных характеристик дерева на криптичность окраски кормящихся на нём дятлов. Использовали дисперсионный анализ (ANOVA), где действующими факторами были род дерева, его жизненность, окружность на высоте груди и вид дятла, а откликами — каждая из 5 составляющих маскировочности окраски (А)-Г).
Результаты
Криптическая окраска работает: дятлы, как правило, хуже видны на кормовых, нежели на обычных деревьях.
Выявлено, что «в глазах пустельги» все 3 вида дятлов в среднем значимо менее заметны на кормовых деревьях (N=31), чем на трансектных (N=61). Эта криптичность достигнута за счёт а) значимо более расчленяющей окраски (среднее значение данного параметра±SD: 0,32±0,03 на коре кормовых деревьев и 0,30 ± 0,03 на трансектных; t90 = 3,11, P = 0,002, d1 = 0,68, 95% ДИ [0,24; 1,12]; рис. 3а1) и б) большего соответствия субстрату по яркости (среднее±SD: 27,85±19,43 для кормовых деревьев и 39,63±21,57 для трансектных; t90 =2,56, P = 0,012, d=0,56,95%ДИ[0,12-0,99]; Рис.3a3).
Не выявлено значимых различий между заметностью птиц «в глазах пустельги» на кормовых vs трансектных деревьях по а) соответствию контрастностей образов дятла и коры (среднее значение параметра±SD для первых 12,72±4,69; для вторых 13,97±4,17; t90=1,30, P = 0,196, d=0,29, 95% ДИ [0,15, 0,72]; рис.3a2), б) по соответствию зернистостей (4.99e03±2.77e03 для первого, 5.40e03±2.51e03 для второго, t90=0,70, P=0,4803, d=0,15, 95% ДИ [0,28-0,58]; рис.3a4), в) по соответствию цветности (среднее±SD на коре кормовых деревьев 0,03±0,02 vs 0,02±0,02 на трансектных; t90=1,88, P=0,064, d = 0,41, 95%ДИ [0,02-0,84]; Рис. 3a5).
«В глазах» клинохвостого орла окраска дятлов существенно более маскировочна на кормовых деревьях сравнительно с трансектными (рис.3b, N =31 и 61), по параметрам а) расчленяющей окраски (среднее±SD: 0,33±0,03 vs 0,32±0,03 соответственно; t90=2,65, P= 0,009, d=0,58, 95% ДИ [0,14; 1,02]; рис.3b1), б) соответствия яркостей (29,45±20,59 vs 42,61± 22,75 соответственно; t90=2,71, P=0,008, d=0,59, 95% ДИ [0,15; 1,03]; рис.3b3) и в) соответствия цветностей (0,07±0,03 vs 0,05±0,04; t90=2,28, P=0,025, d=0,50 по шкале Коэна, 95% ДИ [0,06, 0.93]; рис.3b5).
Не выявлено значимой разницы между образом дятлов на обоих субстратах по двум параметрам: а) соответствие контрастностей (16,67±6,68 vs 18,85±6,56; t90=-1,53, P=0,130, d= 0,33, 95% ДИ [0,10-0,77]; рис.3b2) и б) соответствие зернистостей (7.52e03 ± 3.56e03 vs 7.39e03 ±2.35e03; t90=0,20, P=0,843, d=0,04, 95% ДИ [0,47- 0,39]; рис.3b4). Все значимые эффекты были средними. Полученный результат остался значимым и после анализа False discovery rate (FDR) — прогноза частоты ложноположительных результатов (Benjamini & Hochberg, 1995).

Рисунок 3. Сравнение криптичности образов 3х видов дятлов (среднее по 3 видам: каролинский, пушистоперый и волосатый) на коре кормовых деревьях vs трансектных «в глазах» (а) пустельги (b) орла и (c) при остроте зрения пустельги (она ниже) и орла (выше). Параметры сравнения сверху вниз: расчленяющая окраска; сопоставление контрастностей, освещенностей, зернистостей, цветностей. *P<0,05; **P<0,01; ***P<0,001.
Дятлы, как правило, более криптичны «в глазах» хищника с менее острым зрением
Окраска дятлов более маскировочна «в глазах» пустельги, чем орла, у первой острота зрения ниже (рис. 3B, сравнение только по кормовым деревьям). Разлчия значимы по параметрам а) соответствия контрастностей (среднее±SD для пустельги vs клинохвостого орла = 2,72±4,69 vs 16,67±6,68; t60= 2,69, P=0,009, d=0,68, 95% ДИ [0,17; 1,18]; рис. 3С2) и б) соответствию зернистостей (-4.99e03 ± 2.77e03 vs 7.52e03 ± 3.56e03; t60 =3,11, P=0,003, d=0,78, 95% ДИ [0,27; 1,29]; рис. 3с4). Однако по соотношению цветностей наоборот, дятлы значимо хуже заметны хищнику с более острым зрением, т. е. орлу (средние±SD 0,03 ± 0,02 для пустельги vs 0,07±0,03 для орла; t60 =5,32, P< 0,001, d=1,34, 95% ДИ [0,78- 1,88]; рис. 3С5).
Этого авторы не ожидали и подробно анализируют в обсуждении. Криптичность образов дятлов не отличалась «в глазах» пустельги и орла ни по соотвестствию контрастностей (среднее±SD: 0,32±0,03 и 0,33±0,03; t60=1,17, P=0,245, d=0,29, 95% ДИ [0,20- 0,79]; рис.3с1), ни по соответствию яркостей (27,85±19,43 vs 29,45±20,59; t60±0,31, P=0,754, d=0,08, 95% ДИ [0,41- 0,57]; рис.3с3). Те же данные, взвешенные по остроте зрения потенциального хищника и разбитые по видам дятлов приведены в приложении, рис. А2.
Разные виды дятлов специализированы по характеру маскировочности окраски
Межвидовое сравнение разных аспектов маскировочности окраски (только на кормовых деревьях) выявило, что по некоторым из пяти окраска одних видов намного криптичнее, чем других (рис.4а-с). Пушистоперые дятлы обладают значимо более расчленяющей окраской, чем каролинские «в глазах» и орла (0,351 vs 0,297; P<0,001; рис.4а), и пустельги (0,342 vs 0,291; Р < 0,001; Приложение, рис. А3). Окраска волосатого дятла также значимо более расчленяюща, чем каролинского «в глазах» и орла (0,351 vs 0,297; P<0,001; рис.4а), и пустельги (0,340 vs 0,291; P < 0,001; Приложение, рис. А3). Напротив, маскировочность окраски краснобрюхого дятла значимо лучше чем у волосатого соответствует коре кормовых деревьев по яркости коредеревьев, чем волосатые дятлы «в глазах» и орла (22,763 vs 38,821; Р = 0,011; рис. 4c) и пустельги (22,051 vs 36,540, P = 0,017; Приложение, рис. A3).
[Если бы авторы изучили больше видов и пёстрых дятлов, и меланепресов, можно было б проверить естественное предположение, то первые специализируются на расчленяющем эффекте, вторые же маскируются в первую очередь за счёт пятен бликования].
Не обнаружено значимых различий между видами по таким аспектам маскировочности, как сопоставление контрастностей, цветностей или зернистостей ни «в глазах» орла (рис. 4, е), ни «в глазах » пустельги (приложение, рис. A3). В том числе соответствие контрастностей сильно варьировало внутри вида (приложение, рис. А4), видимо из-за разного качества кожи тушек, подготовленных для исследования (например, видимость белых сегментов перьев может влиять на показатели контрастности). Эта внутривидовая изменчивость, возможно, способствовала невыявлению значимой разницы между видами по ряду параметров маскировочности.

Рисунок 4. Разный характер криптичности трех видов дятлов (столбцы слева направо M.carolinus, D.pubescens, L.villosus) в «глазах» орла, а-е — её составляющие, см. рис. 3. Звездочки — значимые различия по тесту Тьюки: *P<0,05; **P<0,01; ***P<0,001. Использованы лишь кормовые деревья, см.Приложение, рис. A3b. Данные по криптичности тех же видов «в глазах» пустельги аналогичны.
Кормовые деревья значимо отличаются от деревьев трансект по таксономическому составу, жизненности и диаметру ствола (~возрасту)
Кормовые деревья и деревья трансект, использованные в исследовании, значимо различны по трем параметрам (рис.5с). В среднем они принадлежат к разным родам (χ27=46,57, N=92, P < 0,001; рис.5c1): среди кормовых больше ясеней (Fraxinus sp.) и дубов (Quercus sp.), среди трансектных — кленов (Acer sp.), буков (Fagus sp.), ликвидамбаров смолоносных Liquidambar styraciflua и ниссы лесной Nyssa sylvatica (рис.5с1).
Значимо различалось и распределение деревьев между 4мя категориями жизненности (1 живые, 2 усыхающие, 3 преимущественно усохшие, 4 сухие), χ23=24,96, N=92, P<0,001; рис. 5c2. На кормовых деревьях значимо больше сухих ветвей; они также значимо толще, с большей окружностью ствола (среднее±SD=152±65,67 см vs 93±50,98 см, t90=4,80, P<0,001; N=31 и 61, рис.5с3). Следовательно, кормовые деревья постарше наугад взятых в том же лесу; дятлы кормятся на деревьях не случайным образом, сообразно обилию разнотипных стволов в лесу, но следуют видовым предпочтениям, связанным в том числе с визуальными характеристиками коры и степенью маскировочности образа птицы на ней.
Визуальные образы коры кормовых и трансектных деревьев различны
У вторых выше яркость коры (N=61 и 31) как «в глазах» пустельги (среднее±SD, 109,74±25,13 у кормовых vs 124,64±25,02 у трансектных деревьев, t90±2,70, P=0,008, d=0,59, 95% ДИ [0,15- 1,03]; рис. 5а1), так и «в глазах» орла (110,59±25,24 vs 126,11±25,64; t90±2,76, P =0,007, d=0,60, 95% ДИ [0,16; 1,04]; рис.5b1). Напротив, соотношение контрастностей выше у коры кормовых, чем трансектных деревьев. «В глазах» пустельги эти различия близки к значимым (20,82 ±7,90 vs 18,08±5,32; t90=1,97, P=0,052, d=0,43, 95% ДИ [0,004, 0,86]; рис.5а2) и едва значимы в «глазах» орла (76±9,64 vs 23,24±6,91; t90=2,02, P=0,047, d=0,44, 95% ДИ [0,006- 0,873]; рис. 5b2).
В тетраэдре основных цветов цветового зрения птиц гиперобъёмы восприятия коры кормовых и трансектных деревьев показывают сильное перекрытие. Однако есть и значимые различия, и «в глазах» пустельги (рис.5d1, ed2), и орла (рис.5d3, ed4). В целом для обоих хищников кора кормовых деревьев имеет больший зеленоватый оттенок, чем у трансектных. Точки, соответствующие первой, на графике «сдвинуты» к средним значениям длин волн, рис.5d2, d4). Облака точек, соответствующие на графиках этим образам, в случае пустельги (рис. 5d2) заняли несколько меньший объем и размещены концентрированней, чем в случае орла (рис. 5d4).

Рисунок 5. Сравнение визуальных образов коры кормовых vs трансектных деревьев по соответствиям освещённости и контрастности в «глазах» (а) пустельги и (б) орла. (c) Сравнение кормовых и трансектных деревьев по таксономическим родам, жизненности и окружности на высоте груди (CBH). (d) Облака, показывающие положение в 4хмерном пространстве базовых цветов цветового зрения птиц для кормовых (зеленые точки) и трансектных деревьев (фиолетовые) «в глазах» орла и пустельги. *P<0,05; **P<0,01.
 Криптичность дятлов на коре зависит в первую очередь от рода и размера/возраста дерева, а также от вида дятла
Криптичность дятлов на коре зависит в первую очередь от рода и размера/возраста дерева, а также от вида дятла
Дисперсионный анализ ANOVA показал значимое влияние рода дерева и вида дятла на все 5 показателей криптичности образа жертвы «в глазах» орла (табл.1, также см. доп информацию в приложении, табл.). Влияние окружности ствола значимо для всех параметров, кроме соответствия контрастностей; жизненность дерева влияло лишь на один аспект криптичности (соответствие цветности, табл.1). Данные по образам жертвы «в глазах» пустельги очень сходны, и качественно (набор влияний) , и количественно (сила каждого значимого), см. Приложение, табл. A2).

Таблица 1. Значимые влияния (выделены полужирным) на криптичность окраски дятлов «в глазах орда», выявленные дисперсионным анализом ANOVA. Столбцы слева направо — составляющие криптичности в том же порядке, что на рис., строки сверху вниз — охват/возраст дерева; его жизненность; род дерева; вид дятла.
Обсуждение
Эволюция маскировочности окраски дятлов
Авторы показали, что на кормовых деревьях дятлы выглядят лучше замаскированными, чем на трансектных (т.е. случайном подмножестве стволов в их среде обитания) по параметрам расчленяющего эффекта окраски, соответствия яркостей и цветностей [с фоном]. Эти данные поддерживают точку зрения, что риск гибели от пернатых хищников сыграл определенную роль в эволюции оперения всех трех видов дятлов. Они же то дополняют другие исследования, показывающие, что в масштабах микрообитания животные (и/или их яйца) лучше замаскированы именно на используемых ими субстратах, в сравнении с неиспользуемыми (Endler, 1984; Kang et al., 2015; Stevens et al., 2014, 2015, 2017; Troscianko et al., 2016b).

В ряде исследований показан активный выбор субстратов, дающих лучшую маскировку, как у бабочек (Kang et al., 2012; Kettlewell, 1955; Sargent, 1966), пауков (Fernández Campón, 2014; Heiling, 2005), креветок (Duarte et al., 2018; Green et al., 2019), крабов (Nokelainen et al., 2017; Stevens et al., 2014), рыб (Kjernsmo & Merilaita, 2012; Smithers etal.,2018), ящериц (Marshall & Stevens, 2014), черепах (Nafus et al., 2015) и птиц (Bailey et al., 2015; Gómez et al., 2018; Lovell et al., 2013; Stevens et al., 2017). Однако в большинстве таких случаев в микростациях, дающих максимум маскировки, животные отдыхают или размножаются, а не кормятся. Так делают пауки-паутинники (Parawixia bistriata) на листьях, наиболее точно соответствующие окраске их тела (Fernández Campáon, 2014); японские перепела Coturnix japonica гнездятся на субстрате, на котором их яйца наименее заметны (Lovell et al., 2013).
Однако это не исключает и объяснения полученных данных, альтернативного выбору дятлами кормовых микростаций по максимуму «предлагаемой» ими криптичности внешнего облика птиц. Возможно, дятлы попросту выбирают места, наиболее богатые потенциальными жертвами (инстинктивно или под действием лбучения), ориентируясь на особенности ствола и коры, сигнализирующие о таких «кормных местах» (их цвет, шероховатость корки, обилие усохших ветвей и пр.).
Дятлы, чье оперение лучше подходит для таких предпочтительных мест поиска корма, будут «автоматически» менее заметны для хищников, с меньшей вероятностью подвергнутся нападению и т. д. Всё это создаёт давление отбора в пользу особей с большей криптичностью оперения именно на кормовых деревьях. В поддержку этого объяснения приводятся разнообразные данные об их избирательности в выборе кормовых микростаций и кормовых деревьев (Conner, 1980; Conner et al., 1994; Dudley et al., 2012; Gunn & Hagan, 2000; Imbeau, 2002; Jackson, 1970; Kisiel, 1972; Nappi et al, 2015; Williams, 1975), также как данные о корреляции обилия членистоногих с вышеназванными характеристиками деревьев и их коры (Conner et al., 1994; Jackson, 1979; Nappi et al., 2015).
Так, потенциальный корм дятлов обильнее на (и в) более крупных деревьев большинства видов, избираемых для кормления, вследствие большей складчатости и глубокой бороздчатости корки (т.е. более развитая морщинистая текстура обеспечивает членистоногим больше мест для укрытия, размножения и зимовки (Jackson, 1979). Аналогичным образом, мертвые ветви вероятней всего имеют мягкую, разлагающуюся древесину, одновременно привлекательную для членистоногих и удобную для кормодобывания дятлов (Conner et al., 1994).
Род дерева также возможный сигнал обилия пищи: некоторые таксоны из них легче поражаются вредителями леса вследствие более мягкой древесины (Conner et al., 1994), или большее развитие морщинистой корки даёт членистоногим и их яйцам лучший приют (Jackson, 1979) и/или здесь отсутствуют соединений, отпугивающие членистоногих (Phillips & Croteau,1999; Raffa & Berryman, 1983). Возможна и корреляция цвета коры некоторых деревьев с обилием членистоногих, когда, например, бабочки для отдыха выбирают фон, на котором они наименее заметны для хищников (Kang et al., 2012, 2015; Kettlewell, 1955; Sargent, 1966); такой («маскирующий») цвет коры может дать дятлам признаки обилия членистоногих.
В целом произведенное исследование подтвердило связь между численностью членистоногих, криптичностью субстрата для дятлов и выбора последнего для поиска пищи, поскольку окружность дерева (~возраст), таксономический состав, жизненное состояние и яркость коры значимо различались у кормовых и трансектных деревьев. В том числе кормовые деревья а) крупнее (~ старше), б) несут больше сухих веток, в) у них темнее кора по сравнению с трансектными. Кроме того, окружность ствола и род дерева — сильные предикторы нескольких аспектов маскировочности окраски дятлов.
Специализация [разных видов] в стратегиях маскировки
В работе получен ряд доказательств межвидовых различий в характере маскировки [авторский вывод про «специализацию» здесь преждевремен и не обоснован ничем, это только различия]. Пушистопëрый и волосатый дятлы отличаются большим развитием расчленяющего эффекта окраски, чем каролинский; он, в свою очередь, точней соответствует субстрату по яркости. Все это может быть следствием разной экологии видов. Возможно, разные виды дятлов подвержены нападению хищников с разной дистанции и/или атакуются хищниками с разной остротой зрения, что может влиять на предпочтительные способы маскировки их на субстрате (Forsman & Appelqvist, 1998). Для более тщательного изучения этой возможности будущие исследования должны включать больший диапазон дистанций для наблюдения и/или вариантов остроты зрения разных хищников, чем было использовано авторами.
Однако возможны и объяснения, отличные от экологических. Особенности маскировочности окраски того иди иного вида могут следовать из ограничений на эволюцию, сложившихся в разных филогенетических ветвях дятловых или просто отражают факт наличия у оперения других функций, кроме маскировки. Так, краснобрюхие дятлы принадлежат к кладе [бывший род Centurus, ныне группа видов внутри рода Melanepres], члены которой сходны по мелкой поперечной полосатости на спине [чëрно-белой], но населяют разные местообитания и даже ландшафты (Navarro-Sigüenza et al., 2017).
Если этот паттерн полосатости эволюционно консервативен, то простейший способ роста маскировочности окраски состоит в вариировании яркости, достижимой более точной «настройкой» интенсивности черных и белых полос [данного паттерна]. Консервативность паттерна, возможно, установилась в связи с задачами коммуникации, меж- или внутривидовой, тогда как функция маскировки у оперения вторична [и регулируется отбором так, чтобы не затрагивать основное].
Влияние остроты зрения хищника на эффективность маскировки
Выявлена лучшая маскировочность образов дятлов «в глазах» хищника с меньшей остротой зрения (1/40 МРА) по параметрам соответствий контрастности и зернистости визуального образа (по сравнению с хищником с большей остротой зрения, 1/140 МРА). Одно из возможных объяснений совпадения показателей детализации — меньшая острота зрения снижает детализацию, сглаживая паттерны с высокой зернистостью. Если визуальные образы птиц и коры различаются в первую очередь этим (скажем, рисунок оперения мелкий, а шероховатостей коры крупный) то хищнику с меньшей остротой зрения это трудней распознать.
Также снижение остроты зрения понижает контрастность мелкого рисунка: частая черно-белая полосатость выглядит однотонно-серой (=менее контрастной) при меньшей остроте зрения или с меньшей дистанции, чем при высокой остроте. При низкой контрастности фона (коры), черно-белое оперение с мелким рисунком видится более контрастным смотрящему с меньшей остротой зрения.
Неожиданно выяснилась большая криптичность образа дятла по соотношению цветности «в глазах» хищника с большей остротой зрения. Возможно, это вызвано спецификой цветовой заливки воспринимаемой картины: мелкомасштабным узором самого объекта, но крупномасштабным узором фона, на котором от видится зрителю, или наоборот. Меньшая острота зрения размывает цвета, образующие первый, но не второй. Например, мелкий красно-желтый узор при низкой остроте зрения «размоется» у смотрящего до оранжевого, но крупный красно-желтый узор фона узор воспримется как он есть.
Возможно, здесь связь с феноменом, описанным у людей: значимые визуальные объекты, вроде фотографий известных людей) успешней распознают именно при низкой остроте зрения. Тогда, как и при взгляде с большей дистанции, большая зернистость затруднит выделение отличительных характеристик объекта. Когда же узоры высокой степени детализации (мелкомасштабное «кружево») больше не различаются, идентифицировать легче (Harmon & Julesz, 1973).
Резюмируя: авторы [косвенно подтвердили] распространённое мнение, что рисунок оперения дятлов помогает уменьшить риск нападения хищников (Thayer, 1909). [Все исследованные виды и правда] по целому ряду параметров, составляющих маскировочность, хуже заметны именно на коре кормовых деревьев сравнительно с трансектными тех же местообитаний.
[Прямым доказательством было бы непосредственное обнаружение меньшего риска гибели в когтях хищника при кормлении или отдыхе (вообще нахождении на тех же деревьях), чем в каких-то иных ситуациях. Если да (разница риска гибели значима), то наличный паттерн окраски этих видов действительно продукт отбора на меньшую заметность для хищников. Если нет, маскировочные свойства окраски — побочное следствие каких-то других причин, включая рассмотренные авторами выше].
Выполненная работа расширяет понимание роли маскировочных свойств в эволюции оперения, дополняя фундаментальные исследования критичности окраски наземноннездящихся птиц (Mason et al., 2023; Stevens et al., 2017; Troscianko et al., 2016b; Wilson-Aggarwal et al., 2016). Однако авторы признают и её ограниченность. Во-первых, как и во всех наблюдательных исследованиях маскировочных свойств окраски in vivo, возможен непреднамеренный пропуск именно тех соучеников кормления дятлов [и/или на тех деревьях], когда маскировка была наиболее эффективна [с другой стороны, это мажорирование только усиливает выводы автора о маскировочных свойствах, если между выявленными и пропущенными случаями разница только в степени, но не в качестве], и эти субстраты не попадали в выборку данных о кормовых деревьях.
Во-вторых, в работе использовали тушки только самцов. В дальнейших исследованиях надо включить и самок (авторы ожидают у них ещё большей криптичности окраски) из-за отсутствия красного оперения на голове. Минус работы и небольшой объем выборки; нужны дополнительные исследования на разных участках в пределах видовых ареалов, чтобы определить насколько всеобщи сделанные выводы. Кроме того, работа не касается прямого вопроса о действенности маскировки: достаточно ли велики наблюдаемые различия в её составляющих между видами, чтобы влиять на уровень хищничества и выживаемость птиц? Для этого требуются дополнительные исследования охот хищных птиц на дятлов и количественная оценка их выживаемости.
В предшествующих исследованиях взаимоотношений хищник-жертва (Skrade & Dinsmore, 2013; Troscianko et al., 2017; Troscianko et al., 2016b) было и правда показано, что различия в тех аспектах криптичности, что изучали у дятлов — предикторы шансов на выживание жертв, поэтому [здесь типологическая экстраполяция] могут также влиять и на выживание дятлов. Наконец, авторские результаты получены исходя из лучших на сегодня методах анализа паттернов окраски и моделей их визуального восприятия хищными птицами, но надо признать что исследования тех и других находятся только в начале пути (Stoddard & Osorio, 2019).
При всех этих ограничениях, выполненное исследование — ещё один шаг в изучении маскировочности окраски на субстратах, экологически значимых для жертв, и с моделированием их у визуального восприятия хищниками в рамках естественной истории тех и других.
Примечание
1Коэффициент d Коэна — величина эффекта, значимость соответствующих различий между средними (0,2 или менее — малые, около 0,5 — средние, 0,8 и выше значительные, Сohen, 1988).
Рекомендуем прочесть
 Сколько дятлов лесу нужно?
Сколько дятлов лесу нужно? «Сортировка» вместо отбора в контексте урбанизации вида
«Сортировка» вместо отбора в контексте урбанизации вида Проблемы и риски выдалбливания дупел у дятлов
Проблемы и риски выдалбливания дупел у дятлов Вольфганг Шлейдт. Коммуникация животных. Поиски понимания сложности поведения
Вольфганг Шлейдт. Коммуникация животных. Поиски понимания сложности поведения Зачем большим пёстрым дятлам всю зиму охранять территории?
Зачем большим пёстрым дятлам всю зиму охранять территории? Болит ли голова у дятла?
Болит ли голова у дятла? Важность повторного использования дятлами дупел
Важность повторного использования дятлами дупел Расшифровка сигналов агрессии у коростелей
Расшифровка сигналов агрессии у коростелей
