
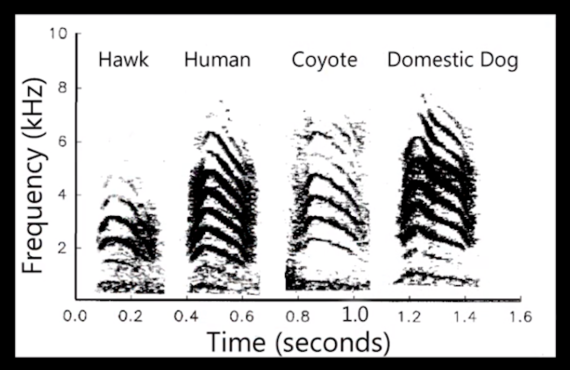
Sound-frequency analysis of praire dog alarm calls, differentiated according to predator
Ещё про «психологическую дуэль» между жертвой и хищником
В январе 1995 года в Йеллоустонский национальный парк вернулись волки, уничтоженные почти за 70 лет до этого в результате сверхусердной борьбы с расплодившимися хищниками. В течение двух следующих зим в парк было выпущено 31 животное, пойманное в Канаде и снабжённое радиоворотничком, чтобы объездчики знали о местоположении волков.
Но что станется с главной добычей волков — оленями-вапити? Все эти десятилетия они усиленно размножались, причиняя немалый вред деревьям парка. Что будет теперь, когда заклятый враг вернулся?
Всё стало ясно уже на второй год. Там, куда волки ещё не добрались, оленухи паслись безмятежно, а вокруг весело скакали телята. «Сцена из диснеевского фильма», — вспоминает эколог Джон Лондре из Университета штата Нью-Йорк в Осуиго (США). В тех же уголках парка, где волки уже освоились, ситуация была совсем другой: телята жались к беспокойным мамам. «То были две страны: в одной царил мир, в другой бушевала война», — говорит учёный.
В этот момент Дж.Лондре понял: волки не просто убивают оленей, они меняют их поведение, не пошевелив ни одной лапой. Само их наличие (запах, который донёс ветер, или след на влажной земле) порождает у жертвы дурное предчувствие. Окружающая местность отныне оценивается в соответствии с риском быть съеденным. Для описания этого эффекта он выдумал термин «пейзаж страха»: «горы» там, где риск высок, и «долины» относительной безопасности, в которых можно слегка расслабиться.

Это простое открытие перевернуло экологию, в которой до той поры считалось, что хищники влияют на объект охоты лишь непосредственным убийством. Кроме того, был брошен вызов распространённому представлению, что большинство животных испытывают страх лишь короткое время — в виде внезапного приступа паники или в ходе погони, тогда как продолжительный психологический стресс — удел людей и других приматов. По словам Дж.Лондре, в традиционных экологических моделях отношения хищника и жертвы рассматривались как игра в шары: если один шар ударит по другому, тот считается выбывшим из игры. Попытка добавить психологический аспект казалась в то время неоправданной антропоморфизацией и была предана анафеме.
Но времена меняются. Сегодня экологи применяют понятие пейзажа страха для изучения не только волков и вапити, но и акул и дюгоней, пауков и кузнечиков. Снова и снова подтверждается догадка о том, что самое большое влияние на поведение жертвы оказывает не убийство, а устрашение. Это сказывается на том, как потенциальная добыча питается, размножается и воспитывает молодняк. Эффект распространяется по всей экосистеме, воздействуя на местную флору и даже на круговорот питательных веществ в почве. Хищнику достаточно быть страшным
Дж.Лондре не был первым, кто осознал экологическую роль страха. Ещё в 1970-х годах исследования показали, что хищники вынуждают жертву выстраивать дорогостоящую защиту — например, мигрировать в менее плодородные области и плохо питаться из-за того, что приходится постоянно быть начеку. Но большинство экспериментов на эту тему обладали невеликим масштабом и малой продолжительностью. Кроме того, почти не рассматривались последствия этого ужаса в долгосрочной перспективе.
Поэтому йеллоустонский пример можно считать первым. Перед тем как там вновь появились волки, экологи совершенно правильно предсказали размеры популяции хищников и то, сколько оленей они убьют. Но поскольку считалось, что волки воздействуют на вапити, лишь убивая их, численность популяции последних оказалась сильно переоценённой, вспоминает Скотт Крил из Университета штата Монтана.
Наблюдения, выполненные С.Крилом в 2002–2006 гг., показали, что в присутствии волков олени пребывали настороже более чем вдвое дольше обычного. Они также уходили с полей в леса, предпочитая безопасность изобилию травы. В результате они успевали съесть примерно на четверть меньше, чем раньше, что привело к падению рождаемости. С.Крил уверен, что дело тут в нехватке сил, а не в нападениях волков, ибо хищники очень редко бросаются на телят. Измерение уровня прогестерона (гормона, пик которого приходится на беременность) в навозе полутора тысяч самок показало, что он сильно снизился в тех областях, где жили волки.
В безмятежные времена йеллоустонская популяция вапити насчитывала 19 тыс. особей, а сейчас едва превышает 6 тыс. Там, где олени проиграли, парк выиграл. В 2010 году Уильям Рипл из Университета штата Орегон сообщал, что волки позволили осине, иве и трёхгранному тополю вернуться в прежние границы. Выживало больше молодых деревьев, ведь напуганные вапити стали реже обгладывать их нижние ветви, а высота старых увеличилась вдвое и даже втрое.
Чем выше дерево, тем больше строительного материала для бобра, и популяция последних выросла с одной колонии в 1996-м до дюжины в 2009-м. Бобровые плотины создали великолепные условия для птиц, земноводных, рыбы и др. А всё из-за того, что волки очень страшные…
Эта разновидность цепной реакции — трофический (пищевой) каскад — хорошо знакома экологам, но опять и снова она понималась раньше как результат нападения хищника на жертву. Некоторые специалисты и сейчас подвергают сомнению существование пейзажа страха, объясняя перемены игрой климата. Но аналогичные изменения происходят не только в Йеллоустоне. Например, пышные луга морской травы на мелководье залива Шарк у западных берегов Австралии обязаны своим существованием именно хищникам. В отсутствие тигровых акул там пасутся дюгони, которые с сентября по май прячутся от врага в глубоких водах, а когда возвращаются, объедают лишь самые верхние листья, потому что приходится всё время держать голову поднятой для лучшего обзора окрестностей. Одного присутствия акул оказалось достаточно не только для того, чтобы изменить численность дюгоней, но и для того, чтобы удержать их от уничтожения морской травы.
Но наблюдений скептикам недостаточно — им подавай эксперименты. Лиана Занетте из Университета Западного Онтарио (Канада) придумала, как заставить животных почувствовать угрозу, но при этом не позволить хищникам никого убивать. Сначала она досконально изучила чириканье подвергшегося нападению певчего воробья Melospiza melodia на островах Галф у западного побережья Канады. Затем она защитила гнёзда электрифицированным ограждением от енотов и проволочной сеткой от ворон и сов. Убедившись в том, что птенцов никто не тронет, она включила запись звуков, производимых хищниками.
Результаты, опубликованные в 2011 году, превзошли все ожидания. Птички, регулярно хватавшиеся за сердце, выращивали в год на 40% птенцов меньше по сравнению с теми, которым проигрывали звуки безобидных тварей. Они откладывали меньше яиц, а те, что удавалось отложить, были легче обычного, и из них реже вылуплялись птенцы — отчасти из-за того, что пугливые самки хуже их высиживали. А вылупившиеся птенцы чаще погибали от голода, потому что родители с трясущимися поджилками приносили меньше еды в гнёзда.
В прошлом году Дрор Хаулена и Освальд Шмиц из Йельского университета (США) показали, что каскад, порождённый страхом, может идти ещё дальше. Они выращивали кузнечиков в больших клетках на свежем воздухе, после чего в половину из них запускали пауков. Хищники не могли убить жертву, потому что им предусмотрительно склеивали ротовой аппарат, но кузнечики об этом не знали. В присутствии пауков скорость обмена веществ бедняг подскакивала на 40%, повышая их потребность в энергии. В результате кузнечики ели больше золотарника, богатого углеводами, в ущерб травам, насыщенным белком. Белок необходим для роста и размножения, но насекомые предпочитали пожертвовать этими радостями ради быстрого пополнения энергозапасов. Это привело к изменению химического состава организма, повысив отношение углерода к азоту в их телах на 4%. Из таких кузнечиков после смерти получалось плохое удобрение. Не сделав ровным счётом ничего, пауки повлияли на круговорот питательных веществ в почве!».
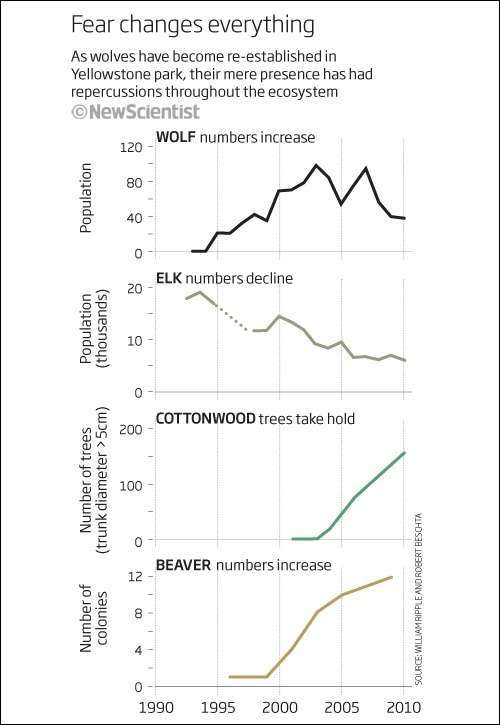
Изменение численности волков в Йеллоустоне отразилось не только на популяции вапити, но и на количестве толстых тополей и колоний бобров.
Биоэкологам давно известно, что виды, расположенные на разных концах пищевой цепи, влияют друг на друга. Но никогда ещё эта связь не оказывалась настолько сложной. Согласно традиционной точке зрения, выживание животных зависит только от того, насколько хорошо им удаётся избегать близкого знакомства с хищниками и насколько удачно они пообедают сами, то есть от взаимодействия с непосредственными соседями по пищевой цепи. Почему травоядные не уничтожают пастбища целиком? — Потому что хищники не позволяют популяции достичь катастрофических размеров. А новая теория говорит о том, что само присутствие хищников вынуждает животных покидать открытые пространства. «Мир остаётся зелёным благодаря страху», — подчёркивает г-н Лондре.
Пейзаж страха помогает не только тем, кто изучает природу, но и тем, кто заботится о её сохранности. Например, Джоэл Браун из Иллинойсского университета (США) ищет ирбисов по реакции на них потенциальной добычи. Нахуры и гималайские тары чуют ирбиса лучше любого зоолога. Они сбиваются в кучу и больше смотрят по сторонам, чем пасутся. Оценка численности ирбисов, полученная на основании таких наблюдений, затем проверяется по навозу и следам. Пожалуй, это первый проект, которому не надо смотреть на животных, чтобы их посчитать.
Похожим образом Бёрт Котлер из Университета Бен-Гурион (Израиль) установил, что нубийскому горному козлу в национальном парке Эйн-Авдат очень сильно мешают туристы. На выходных, когда люди собираются на склонах ущелья, животные пугаются, потому что обзор оказывается перегорожен, и они спасаются бегством, не покормившись как следует.
Подобные исследования позволяют экологам вносить коррективы в планы по защите животных. Например, на юго-запад США планируют вновь выпустить толсторогов, но за время их отсутствия растительность там стала высокой и теперь с лёгкостью скрывает подкрадывающихся хищников. «Они выпустят толсторогов, пумы их перебьют, и будет решено заняться контролем численности пум», — опасается Дж.Лондре. По его мнению, надо просто выкосить кустарник и создать коридоры низкой растительности, соединяющие горные цепи, чтобы пумы не всегда могли поймать добычу.
Дж.Лондре полагает, что будущее экологии — за работой с подобными метафизическими ландшафтами, которые для животных вполне реальны. Надо научиться понимать, где пугливое существо видит «гору», знаменующую собой высокий риск смерти, а где «долину».
Подготовлено по материалам NewScientist.
Дм.Целиков. Напуганные до смерти: о влиянии страха на экосистему
Вот так потихоньку оказывается, что все экологические отношения (вроде «хищник-жертва», внутри- и межвидовой конкуренции и пр.) в экосистеме предваряются информационными.
Так, разные виды птиц видят ультрафиолет, и используют это свойство по-разному. Там, где насыщенный синий/голубой цвет входят в брачные «украшения» самцов, по интенсивности ультрафиолета оценивается «качество» партнёра, как это происходит у лазоревок, волнистых попугаев или шотландских куропаток (у них «светятся» красные брови). Другой момент, более важный для нашей темы — ультрафиолетом «светятся» мочевые метки млекопитающих, что используется хищниками для выбора предпочтительного места охоты. Последнее было показано для обыкновенной пустельги и канюка-зимняка.
А мирные птицы, вроде уток, на территории, где хотят загнездиться, оценивают риск разорения гнёзд по интенсивности мочевых меток хищника. В работе Eichholz M.W. et al. (2012) показано, что утки определяют присутствие мочевых меток хищных млекопитающих (по отражению в ультрафиолетовых лучах или запаху) и избегают гнездиться в местах высокой плотности меток, даже если биотоп в целом вполне пригоден. Авторы использовали в опытах мочу лисицы, нанося метки на опытном участке, в контроле использовали просто воду. Два года подряд на первом участке гнездилось значимо меньше уток, чем на втором (суммарно 97 против 143 гнезд).
В опытах J.T.Forstman et al. (2013) разные виды мелких воробьиных чётко избегали гнездования на участках, обработанных экскрементами тех видов хищных млекопитающих, кто разоряет гнёзда. В результате численность соответствующих видов на опытном участке была сильно снижена в сравнении с контролем, при вполне подходящих местообитаниях. Число видов, поселившихся на опасном участке, было также понижено.
По всей видимости, в этом, как во всех перечисленных выше случаях, птицы заранее оценивают риск хищничества по «сигнальному полю» следов активности соответствующих видов и производят опережающее отражение, уходя из данного района. То же самое получается, если подходящее местообитание маркировать не запаховыми, а акустическими «метками» высокого риска хищничества – потенциальные жертвы его избегают (см. опыты с кукшей).

Кукша Perisoreus infaustus
Помимо «отрицательной» маркировки местообитания сигналами пребывания и активности хищников, при выборе местообитания птицы активно используют и «положительную» — социальную информацию, исходящую от сородичей, о том что им здесь хорошо, они нормально живут и вовсю социально взаимодействуют (особенно поют), нечасто прерываясь на оборонительное поведение, отслеживание потенциально опасных ситуаций и тревожную вокализацию[1]. Этологи называют это вторичными сигналами, «производимыми» намеренно (рекламирование территории, патрулирование границ и пр.) или нет (следы полноценной жизнедеятельности, вроде свежих шишек в дятловых «кузницах» в процессе взаимодействия с социальными компаньонами в группировках.
Их характер и концентрация определяет большую или меньшую притягательность данного поселения для активных вселенцев, старающихся занять территории (в сравнении с притягательностью соседних группировок, также доступных для птиц из «подвижного резерва популяции»). Тем самым вторичные сигналы, формируя сигнальное поле группировок (как отдельных локальных, так и всей сети поселений вида на более или менее обширной территории) выступают «семафорами и маяками», эффект которых связывает различные поселения в общую сеть потоком нерезидентов и/или переселенцев, одновременно делая соответствующий поток не стохастическим, а направленным.
Так возникают «профили притягательности» разных участков для поселения новых особей, противоположные «профилям страха» (см.пример для колоний степной пустельги). «Крутизна» соответствующих градиентов определяет силу и интенсивность потока переселенцев между поселениями вида в соответствующей местности[2]. Что обнаруживается на самых разных объектах и разными методами – у моих ли больших пёстрых дятлов, у колониальных ли видов птиц С.П.Харитоновым, у мелких мышевидных грызунов Н.А.Щипановым и пр., о чём и разговор ниже.
Важно отметить, что высокая эффективность вторичных сигналов как «аттракторов» выбора местообитания показана для подавляющего большинства видов птиц, в самых разных работах, в целом их «притягательная сила» значимо выше, чем эффект собственно местообитания.
Так, на Гавайских островах R.Podolsky использовал вторичные сигналы, чтобы способствовать формированию небольших колоний темноспинных альбатросов Diomedea immutabilis, формирование которых затруднено из-за фактора беспокойства, и привыканию к последнему. Использовали визуальные (плоские и объёмные модели альбатросов в позах ухаживания и отдыха), и записи их вокализаций, транслируемые через громкоговорители. Всего было 81 модель и 12 громкоговорителей, распределённых так, что давало им равную возможность привлечения. Регистрировали число пролетающих альбатросов, которых удалось осадить неподалёку от точек привлечения, на какой дистанции, и чем занялись севшие птицы (начнут ли ухаживать и пр.). На контрольном участке без специального привлечения село лишь 1.8% птиц из пролетающих, только визуальные стимулы привлекли 5.2% птиц, вместе с акустическими – уже 8.2%. Объёмные модели были привлекательней плоских, парные – более одиночных, модели в позе ухаживания – более, чем в позе отдыха. Из 607 приземлившихся альбатросов 55% сели ближе 3 м к аттракторам, 66% из 97 ухаживаний также были ближе 3 м.

Это можно использовать при управлении соответствующими видами птиц — скажем, концентрируя население разных видов в наиболее подходящих участках, если те изолированы внутри труднопроходимого для них ландшафта, например, города, или концентрации населения исчезающих видов на «отдельных «островах» ООПТ. Скажем, обзор 24 современных исследований использования вторичных сигналов для привлечения конспецификов показал, что успех был достигнут в 20 случаях. Например, искусственное проигрывание песни успешно использовали для осаживания мигрантов исчезающего вида овсянок Ammodramus maritimus mirabilis во Флориде именно на охраняемых участках травянистых местообитаний в приморских маршах.

См.аналогичные данные по близкому виду овсянок, или по ещё одному исчезающему виду – виреону Vireo atricapilla. Там проигрыванием видоспецифических вокализаций на 7 экспериментальных участках с подходящими местообитаниями осадили 73 птицы в 2001 и 75 в 2002 г., многие из которых образовали пары и загнездились. На стимуляцию в этом и других опытах успешней всего реагировали второгодки, чем более старшие возраста, они же в основном и гнездились в соответствующих местах (сравни их преимущественное участие в процессах урбанизации). Птицы, привлечённые в 2001 г., и их потомки, гнездились в соответствующем месте и в следующем, хотя стимуляцию уже не проводили. Иными словами, однократное привлечение вида вторичными сигналами может создать жизнеспособное поселение в новом месте.

Черноголовый виреон
Больше того, сейчас понятно, что территориальные виды певчих воробьиных и, шире, лесных дендрофильных птиц, реагируют на вторичные сигналы не хуже колониальных видов. Раньше считалось, что только они подвержены «эффекту Дарлинга» — росту колонии за счёт привлечения вселенцев скоплением активно кричащих и взаимодействующих птиц своего вида больше, чем собственно местообитанием, его кормовыми возможностями и пр. Сейчас понятно, что территориальные виды, оптимальные местообитания которых сплошные и протяжённые, а не точечные, как у большинства колониальных, подвержены эффекту Дарлинга в большей степени, чем вторые. Оно и понятно – также как и колониальным видам, им психологический комфорт и достаточная социальная плотность для успешной коммуникации столь же важны, как и качественные местообитания. Если первое сконцентрировано в немногих точках – формирующихся поселениях вида, то второе есть много где, в то время как оптимальные в кормовом и защитном отношении места размещения колоний сильно ограничены и тоже точечны. Понятно, что лесным территориальным видам в этих случаях надо в первую очередь искать первое, а не второе а, значит, сильней ориентироваться на вторичные сигналы, чем колониальным.
Иными словами, раньше думали, что животные просто «прячут» гнёзда в тех микростациях и таким способом, как требует соответствующий инстинкт, а дальше уж как кривая вывезет (тем более что «инстинкт слеп», а отбор «отберёт лучших прятальщиков»).
Но нет, они активно используют оба типа сигналов в собственных интересах – вторичные сигналы от жизнедеятельности и особенно социальной активности конспецификов для «притяжения», «метки» повышенного риска хищничества для избегания соответствующих местообитаний. Кормовая их привлекательность оценивается независимо и накладывается на эту оценку. В соответствии с «профилем притяжения» и «профилем страха» структурируется сеть поселений вида весной, в период распределения территорий, так, чтобы летом, в существенно иных условиях, риск разорения гнёзд находящихся на этих участках, был наименьшим для всей системы, а возможность обеспечения пищей 5-9 птенцов — наибольшей.
Т.е. они не только ориентируются на долговременный выигрыш, жертвуя краткосрочным[3], но и координируют свои реакции на локальные проблемы так, что соответствующий ответ
а) включает изменения на территории, существенно большей, чем проблемная область (в данном случае участки с повышенным риском хищничества),
б) оптимизирует ситуацию на этой существенно большей территории не столько для каждой отдельной особи, сколько для системы в целом.
Сходный пример описывается в обзорных работах Н.А.Щипанова (2000, 2003), Н.А.Щипанова и А.В.Купцова (2004) для видов мелких млекопитающих с хорошо развитой системой популяционной компенсации после истребления населения вида на ограниченном участке, созданного какой-либо локальной катастрофой. Локальная проблема вызывает движение на существенно большей территории, при этом активизируется особая часть популяции — нерезидентные особи из «резерва», в норме поддерживающие устойчивый поток переселенцев между группировками. Они заселяют по новой «зачищенную» территорию, причём в потоке переселенцев особо активизировано размножение (скажем, присутствуют уже беременные самки). Это позволяет восстановить численность вида в кратчайшие сроки, причём происходящая компенсация вполне точная — численность восстанавливается на уровне чуть выше прежнего. Всё вышеописанное показано как для «диких» видов мелких млекопитающих в природных и сельскохозяйственных ландшафтах, так и для синантропных видов грызунов в урболандшафтах.
Прежде всего, эти «профили страха» и «профили притягательности» суть формы популяционной саморегуляции[4]. Они устанавливают социально-обусловленную анизотропность пространства, значимую для резидентов и активных вселенцев, как внутри группировки (предпочитаемый центр и «худшая периферия»), так и между ними, на местности, занятой данной популяцией («лучшие» и «худшие» группировки).
«Широко известно, что разные части колонии имеют неодинаковую привлекательность для размножающихся особей (Coulson, White, 1960; Сoulson, 1971; Burger, Shisler, 1980). Наиболее предпочитаемая часть колонии называется ее биологическим центром. Менее предпочитаемые области колонии называют периферией. В ряде случаев вводится еще подразделение “перицентрия” или “субцентр” – области, лежащие возле центра (Линг, 1965; Coulson, 1985). Выделяется также “край” колонии не в геометрическом, а в функциональном смысле.
Первые прилетающие с зимовки озерные чайки садятся в наиболее предпочитаемый район колонии – ее биологический центр. Затем в течение 2–3 первых дней они занимают всю площадь будущей колонии, хотя плотность их заселения невелика. В дальнейшем плотность гнездования растет по мере образования микроколоний и заселения новых птиц в пустые места между микроколониями. Буквально с первых дней прилета, несмотря на избыток свободных мест, озерные чайки пытаются вторгнуться в уже занятые области колонии. Наблюдения за произвольно выбранными летающими чайками показали, что доля вторгающихся птиц среди летающих чаек возрастает от края колонии к ее центру (Харитонов, 1983). Первые гнезда всегда появлялись в центре колонии (Харитонов, 2006).
Озерные чайки имеют тенденцию время от времени менять положение своей гнездовой территории в колонии. Только единичные особи гнездятся в том же районе колонии, где они заняли свою первую в данном сезоне территорию. В результате переселений озерные чайки перед гнездованием оказываются в более предпочитаемых районах колонии. В течение одного сезона от30 до 71% чаек с разных сплавин пытается переселиться в более предпочитаемые области колонии, реально переселяется 12–36% птиц. Чем дальше от центра расположена сплавина, тем большее число чаек с нее пытается переселиться в более предпочитаемую область и действительно туда переселяется (Харитонов, 1983а). Кроме центростремительных, есть менее интенсивные центробежные переселения, то есть переселения от центра к периферии. От сезона к сезону чайки переселяются на большие расстояния, чем в течение одного сезона (Харитонов, 1993). Переселения внутри сезона и между сезонами являются продолжением одно другого, образуя непрерывную цепочку переселений. В среднем в течение сезона в колонии озерных чаек оз. Киево центросремительный поток переселенцев был в 2.5 раза интенсивнее центробежного (Харитонов, 1983а; Харитонов, 1993).
Молодые птицы имеют больше шансов вторгнуться в менее предпочитаемые районы колонии (край и периферия), где конкуренция меньше. С возрастом птицы становятся более конкурентоспособными и многие из них переселяются в центр. Из-за того, что в колонии озерных чаек центростремительный поток сильнее центробежного, следует, что с возрастом все большее число чаек переселяется ближе к центру, и в центре колонии гнездится больше птиц старших возрастов.
История колонии оз.Киево показывает, что предпочитаемые места в колонии играют в жизни колониальных птиц большую роль. На этом озере в 1983–1987 гг. падение численности вместе с центростремительными переселениями вызвали перераспределение чаек по озеру. Начиная с 1983 г., из-за начавшегося разрушения большого плавающего острова, который являлся центром колонии, каждый сезон разорялись тысячи гнезд. В то же время, успех размножения на береговых сплавинах был значительно выше – там ни разу не было отмечено большого разорения гнезд. Однако, вопреки ожиданию, это не вызвало массового переселения чаек на береговые сплавины. На фоне уменьшения численности колонии и освобождения мест в центре для многих чаек центр оказался более привлекательным, чем стабильное и более успешное место на береговых сплавинах. Площадь колонии стала сокращаться от периферии к центру до полного изчезновения колонии. Этот пример показывает, что озерным чайкам “психологический комфорт” при нахождении в центре колонии важнее успеха размножения (Харитонов, 1993). В таком странном с точки зрения естественного отбора явлении автор видит стремление птиц сохранить имеющиеся связи в колонии, то есть стремление прежде всего “сохранить систему”. По данной причине и возникает примат системности над адаптивностью.

Черношейные поганки с птенцами
Подобное явление отмечено и у черношейных поганок (Podiceps nigricollis). В смешанной с озерными чайками колонии многие поганки стремятся загнездиться в центре колонии озерных чаек, но именно в центре своей колонии озерные чайки сильнее всего беспокоят поганок, создавая помехи их гнездованию. В результате кладка черношейных поганок может быть достоверно меньше в центре, то есть в той части колонии, куда стремится большинство поганок (Чухарева, Харитонов, 2009). Подобных случаев описано довольно много (Панов и др., 1985; Burger, 1982). В этом плане примечательным выглядит характер распределения особей на гнездовании в одной из локальных популяций малых крачек Восточного Сиваша. В данной популяции больше половины особей (судя по ряду признаков – наиболее “качественных” особей) из года в год упорно гнездятся в месте, где успех размножения равен нулю из-за постоянно возникающих нагонных ветров, неизменно разрушающих колонию (Kharitonov, 1999).
У тонкоклювых кайр в Тихоокеанском регионе (где состояние биотопа гнездования не столь стабильно, как в северной Атлантике) на базарах также имеются наиболее предпочитаемые места, куда стремится значительно больше птиц, чем в другие части базара. Интересно, что при этом в колонии может быть много незанятых карнизов. Однако ищущие место особи предпочитают уже заселенные (Kharitonov, 2000).
Есть виды, например, черноголовый хохотун, макроструктура колонии которых непостоянна. Оказалось, что центральная часть колонии, т.е. место, куда заселялись первые птицы, у этого вида сильно меняется по годам. Так, в 1989 г. центр составлял 0.28 часть большой колонии черноголовых хохотунов, а в 1990 г. – 0.45. В 1991 г. в колонии было два центра, расположенных на некотором расстоянии друг от друга. Данные наблюдения и статистические тесты показали, что центр колонии у черноголовых хохотунов по привлекательности эквивалентен другим частям колонии (Kharitonov, 1999а). Подобную макроструктуру колонии с отсутствием постоянных предпочитаемых мест имеют также морской голубок, и, скорее всего, пестроносая крачка и черноголовая чайка. Равнозначность всех мест в таких колониях в какой-то мере “оправдана” тем, что представители этих видов селятся сразу с большой плотностью, и в такие колонии нельзя вторгнуться между уже имеющимися парами, можно заселиться только с краю. Это отличает данные виды от основной массы других колониальных птиц, чьи колонии имеют постоянный градиент предпочитаемости в течение ряда лет.
После классических работ Джона Коулсона (Coulson, 1968, 1971; Coulson, White, 1960), эффект “центр–край” уже может считаться общеизвестным свойством структуры колоний околоводных птиц. В развитие концепции неодинаковой предпочитаемости разных районов колонии мы показали, что макроструктура колонии может быть не только статической, но и динамической.
Примером статической макроструктуры являются многие колонии моевок в Великобритании, где сегрегация особей происходит при первом гнездовании. Затем на протяжении всей жизни птицы гнездятся в центральной или периферийной части колонии (Coulson, Dixon, 1979). У озерных чаек и тонкоклювых кайр в тихоокеанском регионе колонии имеют динамическую макроструктуру: птицы могут менять место гнездования в течение жизни, переселяясь из менее предпочитаемой в более предпочитаемую часть колонии.
Результаты проведенных исследований указывают, по крайней мере, на три критерия, которые позволяют определить разные по предпочитаемости части колонии. Первым критерием является последовательность посадки самых первых прилетевших весной на место гнездования птиц, которые появляются в центральных частях колонии. Второй критерий – в более предпочитаемые части колонии пытается вторгнуться большее количество птиц, чем в другие части колонии. Третья особенность состоит в сильном центростремительном потоке переселенцев внутри колонии, однако это касается только колоний с динамической макроструктрой. Все три критерия были выявлены на примере озерных чаек (Харитонов, 1983, 1993). Эти закономерности отмечаются и у многих других колониальных околоводных видов птиц. Даже такие консервативные (в отношении места гнездования) птицы, как тонкоклювые и толстоклювые кайры, могут сменить место при заселении птичьего базара. У белых гусей тоже обнаружены предпочитаемые места, существующие из года в год (Сыроечковский, 1981)…
…Ряд наблюдений указывает, что отдельные колонии в группах близлежащих колоний могут быть связаны между собой неслучайным образом. Основную работу по данному вопросу с индивидуально помеченными озерными чайками проводили в колониях поймы р. Москвы в 1983–1986 гг. в районе пос.Виноградово, в 75 км к юго-востоку от Москвы. Многие колонии озерных чаек формируются здесь во время весеннего разлива. В пойме р.Москвы мы различали постоянные колонии, существующие много лет, и временные, существующие 1–2 года (Харитонов, 1998).
Оказалось, что если рассматривать всю совокупность колоний Виноградовской поймы, то в пределах всей этой совокупности отдельные колонии имеют сходную градацию по трем признакам, определяющим макроструктуру колонии.
Так, колонии озерных чаек в пределах участка поймы образуют надколониальную систему, которая функционирует как одна большая колония. Макроструктура надколониальной системы соответствует макроструктуре колонии. В пределах такой надколониальной системы есть более и менее предпочитаемые колонии. При смене места гнездования озерные чайки стремятся переселиться в более предпочитаемые колонии поймы.
Центральная колония поймы (не геометрический, а биологический центр системы) была наиболее привлекательна для чаек. Первые прилетевшие в пойму чайки появлялись именно в этой колонии. Здесь отмечен наиболее высокий уровень вторжений заселяющихся птиц. Как в отдельной колонии, в системе колоний обнаружен центростремительный поток переселенцев. Относительное число меченых чаек (процент от числа всех встреченных меченых чаек в сезоне) в центральной колонии поймы с годами рос. Не только внутри колонии, но и в пределах группы колоний, расположенных в одной местности, имеется четкий центростремительный поток переселенцев, почти в три раза более интенсивный, чем от центра (Харитонов, 1998).
Ряд полученных данных свидетельствовал о том, что надколониальная система Виноградовской поймы, в свою очередь, является частью более обширной надколониальной системы (Харитонов, 1998). Объединение колоний в надколониальную систему происходит по иерархическому принципу.
Сходство характеристик колонии и надколониальной системы дает возможность заключить, что принципиальной разницы между колонией и надколониальной системой нет. Сходство характеристик колонии и группы колоний позволяет говорить о том, что понятие “колония” принципиально неточно. Колонией можно назвать любую пространственно обособленную группировку гнезд, внутри которой могут присутствовать и группировки более низкого ранга. Эти группировки тоже можно называть колониями. Например, отдельные сплавины, заселенные чайками на оз. Киево, мы вправе называть “частями колонии” или считать их отдельными колониями…
Из принципиальной неточности определения колонии следует очень важный методологический момент: мы можем рассматривать любую концентрацию гнезд как колонию, не принимая во внимание наличие группировок гнезд внутри этой концентрации.
Четырехлетние исследования (1989–1992 гг.) с помеченными крылометками птицами проведены и в группе колоний черноголовых хохотунов на Восточном Сиваше. У этого вида переселения из колонии в колонию от сезона к сезону указывают на случайный выбор места гнезда, то есть черноголовые хохотуны не имеют предпочитаемой колонии внутри надколониальной системы, и все колонии для них эквивалентны. В течение одного сезона размножения или в разные сезоны в колониях черноголового хохотуна нет направленных переселений не только на уровне отдельной колонии, но и на уровне надколониальной системы. Отсутствие постоянного градиента предпочитаемости – это тоже свойство пространственных группировок гнезд разного ранга. Поэтому, такая же неточность понятия колонии имеет место и у ряда облигатно-колониальных (термин по: Зубакин, 1976) видов, чьим представителем является черноголовый хохотун. Для видов, у которых разные части колонии эквивалентны по своим свойствам, такая же эквивалентность наблюдается и у надколониальных систем (Kharitonov, 1999b)».
С.П.Харитонов, 2011. Пространственно-этологическая структура колоний околоводных птиц// Зоол.журнал. Т.90. №7. С.846-860.
Таким образом, предварение непосредственных взаимодействий сигнально-информационными, благодаря которым особь может
а) оптимизировать своё поведение относительно состояния всей (социальной, популяционной) системы, а не только ближайшего окружения, т.е. использовать «в своих интересах» реализованное в системе дальнодействие сигналов и событий;
б) сделать это с опережением, до того как «кулак судьбы откроет глаза», так что он «открывает» их лишь неточно реагирующим на сигналы или «плохо продуцирующим» их особям[5],
— касается не только «собственно коммуникации» — взаимодействий между конспецификами, конкурирующими за социальный ресурс, вроде самок, территорий и статусов в иерархии, где действия особей в отношении друг друга опосредованы специально выделенными формами поведения – демонстрациями с прямой непосредственной сигнальной функцией. Как видно из вышесказанного, всё это верно для взаимоотношений «хищник-жертва», а также для конкуренции за гнездовые дупла и, шире, за гнездовые местообитания и пр., так что предварение непосредственных взаимодействий сигналами тотально.
Что отсюда следует? Во-первых, ещё до того, как популяция жертв начнёт реально нести потери от хищников, они начинают их бояться, и соответственно менять поведение — или чтобы выйти из-под воздействия страха, или как-то с ним жить. А это позволяет и хищникам и жертвам осуществлять опережающее отражение, и по его результатам «сортироваться» на выбравших одну тактику приспособления к данному экологическому фактору или другую. Причём «сортировка» идёт до того, как отбор в виде хватания хищником и разорения гнёзд реально начнёт действовать, последний лишь «закрепляет» её результаты, элиминируя хуже реагирующих на соответствующе сигналы или продуцирующие их недостаточно точно, с «искажениями» относительно значимой ситуации или воспроизводимой формы сигнала. То же самое верно в отношении элементов сигнального поля, устанавливающих «профили привлекательности», только знак (-) меняется на (+).
Благодаря этому информационный обмен позволяет «рассортировать» особей разного качества и с потенциями развития разного (или противоположного) поведения так, что каждая из них с высокой вероятностью попадёт в среду, наиболее благоприятную для развития её собственных качеств и потенций[6]. Это происходит до всякого отбора и помимо его; отбор носит лишь очищающий характер, закрепляя результаты подобной сортировки и устраняя её огрехи — и тем самым совершенствуя механизмы сортировки, они же механизмы коммуникации и популяционной саморегуляции.
На разных видах показано, что подобная «сортировка» особей по потенциям развития поведения, реально происходит и производится благодаря градиентам «привлечения», основанным на сигнальном поле, и отталкивания от «профилей страха». Благодаря такой сортировке, составляющей важную часть и обязательный элемент внутрипопуляционной регуляции, особи с разными потенциями, носители альтернативных стратегий с высокой вероятностью попадают в среду, наиболее подходящую для развития именно их поведения (внутреннего состояния, морфологии и пр.).
Почти всегда сортировка идёт вдоль градиента[7], на котором в одну сторону растёт уровень социального стресса (напряжённость конкуренции между индивидами за символические субституции ресурсов), а в другую – уровень средового стресса (степень экстремальности среды которой всякая особь противостоит независимо от прочих).
Понятно, что соответствующий градиент в равной мере протягивается как между разными поселениями одной местности (от «лучших» биотопов или группировок к «худшим»), так и между соседними географическими популяциями, отличающимися положением на градиенте «центр-периферия» ареала. Так, «внутри ареала вида фиксируется градиент «центр – периферия», вдоль которого и происходит главный процесс сортировки особей, дифференцирующий местные популяции на этом градиенте настолько, что они даже становятся подвидами. От центра к периферии популяции падает уровень социального стресса (значит, конкурентность в условиях повышенной плотности делается менее значимой для итоговой приспособленности особей), но растет уровень энвиронментального стресса (и терпимость к нему дает все больший вклад в приспособленность). Соответственно, особи, придерживающиеся более конкурентной жизненной стратегии, при перемещениях и переселениях «отсортировываются» в более центральные популяции, придерживающиеся более патиентной – в более периферийные (рисунок 1А).
Этот механизм работает, способствуя все большей дифференциации между смежными популяциями на градиенте не вопреки, а благодаря устойчивому обмену особями между ними, и в том случае, если в каждой из местных популяций степень патиентности-конкурентности жизненных стратегий распределена колоколообразно соответственно местным условиями (рисунок 1Б).
Далее, работа Dias et al. (1998) показывает, что представление популяционной динамики вида «источниковыми» и «стоковыми» популяциями, связанными асимметричным переселением, для ряда видов уже подкреплено эмпирически. Переселения особей и воспроизводство локальных группировок здесь организовано так, что максимизируется устойчивость и репродуктивный выход системы в целом, а не какой-либо из категорий индивидов («лучших»).
И наоборот – жизнеспособность локальных поселений и устойчивость воспроизводства в условиях непрерывного выбытия особей в силу смертей и/или переселений больше зависит от устойчивости подтока переселенцев со стороны, занимающих эти вакансии, нежели от успеха размножения местных птиц, связанного с «качеством»/выработанностью локальных адаптаций. Иными словами, воспроизводство специфического паттерна системной структуры оказывается управляющим фактором, перемещения и взаимодействия индивидов в рамках популяционной системы – управляемым откликом. Впрочем, авторы не доходят до системоцентрического взгляда на популяционную динамику, а честно фиксируют несоответствие «морфологического подхода» к популяционной системе (частью которого являются успешно развивающиеся сейчас модели типа «источник – сток») традиционному взгляду, что дисперсия особей препятствует видообразованию (Winker, 2000). Изменение представлений о популяции в сторону системоцентризма позволяет снять это противоречие: не препятствует, а способствует, так же, как межпопуляционной дифференциации. (link).
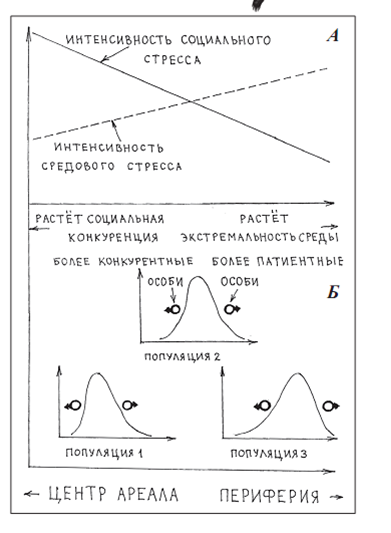
Популяционная система вида согласно «морфологическому подходу»: градиенты «центр – периферия» ареала (А) с сортировкой особей по потенциям (степень развития конкурентной и патиентной стратегий) вдоль них (Б).
Соответственно, более дальние переселения происходят пропорционально реже, и менее интенсивным потоком, но для связывания территориальных элементов популяционной системы вида в одно целое этого достаточно. Что обеспечивает всё большую дивергенцию между разными популяциями на градиенте, причём без потери связности между ними, а то и с усилением. Фактически именно она «пролагает пути отбору», которому остаётся лишь «очищающая» роль, одновременно ускоряя процесс приведения разных фенотипов и жизненных стратегий в максимальное соответствие среде обитания, с одновременным снижением цены элиминации.
***
Тут я вижу основу для возрождения номогенеза, акцентирующего направленность и закономерность в эволюции биосистем. Почему и что это за направленность с закономерностью? Вспомним упомянутый выше трейд-офф между вложениями в социальную активность (цель – размножение) и в самоподдержание (цель выживание). Лучшее и, главное, опережающее реагирование на опасность по вторичным сигналам, а не по фактам самих нападений хищника позволяет животным больше вложить в приобретение территории или статуса, их удержание занятие в условиях конкуренции,в приобретение партнёра и пр. И наоборот, более точная сигнализация об опасности сплошь и рядом поддерживается половым отбором, независимо от того, идёт речь о специфических сигналах тревоги[8], или неспецифических телодвижениях и прочих актах поведения, ассоциированных с риском нападения хищника.
То есть особи используют информацию, циркулирующую по популяционной или социальной системе, заключённую в сигнальном поле (профили страха & профили привлечения) и коммуникативной сети сообщества соответственно, тем более что они демонстрированием во взаимодействиях постоянно «вкладываются» в поддержание этой сети, независимо от того, как и сколько используют «общую» информацию. Благодаря этому они реагируют на опасные места и/или благоприятные возможности с опережением, оказываясь в среде, наиболее подходящей для развития их поведенческих потенций, с существенно большей вероятностью, меньшим риском и меньшей затратой времени, по сравнению с ситуацией, если бы информации, циркулирующей по системе, просто не существовало, и животные избегали опасности/выбирали благоприятные места только собственными усилиями, без опоры на вышеописанный общий ресурс.
В свою очередь, эта «опора», непрерывно поддерживаемая сигнальным поведением индивидов во взаимодействиях друг с другом и между ними, меняет как жизненные стратегии особей, так и популяционную систему. В первом случае она способствует ещё большему выделению альтернативных стратегий в популяции, ещё большей специализации поведения особей в каждой из альтернативных стратегий, при проигрыше промежуточных особей, уменьшении их частоты и пр. Во втором – описанный механизм внутрипопуляционной регуляции служит[9] не только устойчивости (статике системы в условиях определённой интенсивности «возмущений» среды), но одновременно её динамике, эволюции.
Происходит направленное развитие популяционной / социальной системы вида – структура меняется, а регуляторные механизмы совершенствуются. причём селективные процессы, конкуренция и отбор выступают не движителем («мотором» и «рулём»), но лишь «топливом», стимулируя процессы саморазвития системы и «закрепляя» результаты вышеописанной сортировки. Он лишь отсеивает особей, неточно реагирующих на системную информацию и/или плохо «продуцирующих» соответствующие сигналы — в недолжных ситуациях или с существенным искажением формы.
Естественным следствием этого оказывается совершенствование системы информационного обмена на уровне социума и популяции в целом (рост отделённости ритуализованного поведения от повседневного, большая точность продуцирования сигналов, и реакций на них), вместо ожидаемого во всех микроэволюционных моделях сдвига признаков особей. Особи лишь приспосабливаются к популяционной структуре и, соответственно, социальной среде группировок, направленно меняющейся в ответ на средовой стресс на значительных территориях, часто с существенным запозданием, что хорошо видно в процессах урбанизации «диких» видов.
Выделенное шрифтом – именно то, что номогенетические модели эволюции стараются обосновать, механизмов чего они искали и не нашли (а сейчас вырисовываются, как описано выше), причём наблюдаемые «на собственной территории» СТЭ, в микроэволюционных процессах. Именно, направленное изменение пространственно-этологической структуры системы в сторону большей устойчивости воспроизводства и меньшей чувствительности к существующим типам «возмущений», вроде однотипное с рационализацией онтогенеза стабилизирующим отбором, только для видов и популяций. Здесь конкуренция и отбор лишь средства, позволяющие системе – социуму ли, популяции ли — «уйти» от существующих конструктивных противоречий в организации, «снять» их.
«Мотор» же и «руль» происходящего — информационные взаимодействия в системе, оценка особями социальной активности других конспецификов и биоценотических партнёров. Тем более что как минимум у позвоночных есть какие-никакие, но мозги, способные оценивать «качества» партнёра, и возможности регуляции внутреннего состояния, а также запас устойчивости, достаточный для автономизации социальной жизни вида и поддерживающего её информационного обмена от внешней среды. Соответственно, они всё больше «берут в свои руки» все выборы жизненных стратегий, дальше «оцениваемых отбором» с помощью большей и меньшей приспособленности.
См., например, как самки попугаев решают, быть или не быть половому отбору. Самое интересное здесь то, что половой отбор, выявляющий «лучших» самцов, «выключается» в интересах устойчивости популяционной системы как целого хотя казалось бы, именно при их перепроизводстве конкуренция должна особенно обострять чтобы к воспроизводству пошли «самые лучшие». Но нет она снижается и именно потому, что иначе дестабилизируется то системное целое, в котором идёт конкуренция — пространственно-этологическая структура популяции. Иными словами, эффект всякого движущего отбора должен анализироваться не сам по себе но в контрбалансе с давлением стабилизирующего отбора, охраняющего целостность надыиндивидуальной системы (социума, популяции), в рамках характерных отношений которой только и возможно «состязание» особей. Отбор, слишком быстро меняющий признаки индивидов или структуру отношений в социуме, имеет то неприятное побочное следствие, что падает жизнеспособность целого. Поэтому в ответ или стабилизирующий отбор ограничивает этот движущий (превращая его в циклический) или запускаются селективные процессы «отдачи», изменяющие другие признаки особей в контрапункте с изменением самого первого, чтобы организация не вымерла в процессе селекции
Вот более радикальный вариант той же самой модели, предложенный Оловниковым. В отличие от неё, мне более правдоподобным кажется ситуация, когда при выборе поведения, формировании морфологии/жизненной стратегии в онтогенезе и прочих выборах, «оцениваемых» отбором, организм ориентируется не на внутренний сигнал, скажем, поданный стрессом, а на внешний (-ие). Например, созданные сигнальным полем группировки, демонстрациями конкретного конкурента, с которым надо справиться или, если не можешь, поддаться и терпеть или уйти восвояси пр. Тем более что он работает и для «глупых» особей
Чем лучше получается у животных оценивать «качество» друг друга (что по понятной причине одновременно усиливает социальную связанность между индивидами и социальное влияние на такую оценку), тем эффективней «сортировка» и сложней социальность, что ведёт к ещё большей автономизации и пр. Фактически «оценивание» и подбор компаньонов «своими силами» дополняется «опорой» на коммуникативные сети социума и сигнальное поле популяции, оба механизма взаимно усиливают друг друга. Результат оказывается один – сортировка особей по потенциям развития определённого поведения (в известной степени и внутреннего состояния, и морфологии — всего, что «тянут за собой» поведенческие изменения) в определённой среде, а они разные в разных группировках, с понятными эволюционными последствиями в виде саморазвития системы.
Фактически через подобные механизмы уже высшие животные начинают контролировать эволюцию собственных популяций, если не управлять ей, то как минимум ускорять движение по сложившемуся креоду. Естественный отбор здесь никакой творческой роли не играет, это лишь средство, используемое для «испытаний» того что эволюционирует — структуры системы, примерно как общество в том же смысле «использует» классовую борьбу и политическую жизнь для прогрессивного развития, хотя бы в смысле «уйти от нынешнего состояния», не устраивающего слишком многих.
Развитием этих идей можно прийти к номогенезу «нового образца», модели которого дают проверяемые следствия и допускают простую проверку в поле и в опыте. Номогенез «старого образца», Любищева и Мейена, слишком много сил клал на доказательство «ненужности отбора» или «недостаточности его для эволюции», так что не оставалось ничего на «положительную программу» — чем именно обеспечивается искомая направленность. Плюс «борьба с отбором» — борьба с ветряными мельницами, ведь его эффекты строго показаны, если он есть, от него не уйдёшь. Сейчас же понятно, что повсюдность и действенность отбора для номогенетических концепций не плохо, а хорошо — конкуренция и отбор оказываются «топливом», питающим направленное развитие надындивидуальных систем — социумов, популяций и видов.
Примечания:
[1]Я уже писал, что у всех позвоночных, от гуппи и девятииглых колюшек (не путать с трёхиглыми!) до грызунов и снежных баранов включительно, существует трейд-офф между вкладами в социальную активность, так или иначе связанную с последующим размножением и/или предписывающим его формы и сроки, и вкладами в витальную активность, в самосохранение и пр. У таких социальных существ, как птицы с млекопитающими (иногда и рептилии, и костистые рыбы), последнее также делается коллективно. Тревожный сигнал адресован не только хищнику, но и прочим кормящимся особям, чтобы те «взвесили» риск «с точки зрения» собственных обстоятельств, а реакция на сигнал предполагает не только спасение определённым способом, но тот или иной способ ретрансляции сигнала дальше. Это верно и в случае использования видом сигналов-стимулов, а тем более уж сигналов-символов.
[2] Поскольку структура ареала вида фрактальна, картина переселений, вполне изоморфных вышеописанным местным, складывается и в более мелком масштабе, между соседними популяциями вида и потом, между географическими расами. Соответственно, более дальние перемещения происходят пропорционально реже.
[3]дополнительные затраты времени и энергии на оценку опасности в будущем и перемещение в новый район с непредсказуемыми последствиями – и всё это лишь на основании «доверия» к сигналам, да ещё и чужим, независимо от возможности (иногда и реального присутствия) «обмана».
[4] Также как коммуникативные сети внутри сообщества, позволяющие особям, не участвующим во взаимодействиях прямо сейчас, совершенствовать поведение, «подсматривая» за актуальными участниками и корректируя собственную модель поведения;, а также место и время вступления в следующее взаимодействие на основании информации, переданной их демонстрациями и/или исходом их взаимодействия.
[5]Например, недостаточно стереотипно воспроизводящим форму сигнала, так что она искажается, или недостаточно чётко привязывающим сигнал к ситуации, так что он оказывается ложным.
[6] Как минимум у всех «социальных» видов позвоночных, хотя «профили страха» должны быть существенны и для «несоциальных» видов.
[7]Точней, в тех конкретных исследованиях, где этот механизм выявлен или предполагается с высокой вероятностью, она идёт вдоль данного градиента. Кроме того, разделение на альтернативные стратегии, описанное у самых разных видов позвоночных, основано на оппозиции «большая успешность в социальной конкуренции и большая уязвимость к экстремальной среде» vs «слабость и уступчивость в социальной конкуренции с высокой устойчивостью к неблагоприятным условиям внешней среды».
[8] Так, в опытах, где курица выбирает из 2-х незнакомых петухов, и спаривает с выбранным – они конкурируют у неё на глазах, а она лишена какой-либо более долговременной информации о них, — вдруг оказывается, что не важны «брачные украшения»,вроде размера и цвета гребня. Но зато эффективным предиктором успеха разных петухов оказывается интенсивность и точность подачи криков тревоги «с воздуха» и «с земли», также как доминантный статус. Роль второго понятна, а вот первого иначе как через поддержку наибольшего «вклада» в поддержание такого публичного ресурса, как «профиль страха», не объяснишь.
[9] Помимо очевидной и немедленной выгоды — в виде снижения груза элиминации и ухода от дилеммы Холдейна.
Рекомендуем прочесть
 Спаситель курса акций и его упитанные апостолы
Спаситель курса акций и его упитанные апостолы «Безответственность граждан» по-саксонски
«Безответственность граждан» по-саксонски В Стокгольме туман
В Стокгольме туман «Study of African IQ levels proven to be substandard»
«Study of African IQ levels proven to be substandard» «Блэз Компаоре — мёртв, Томас Санкара — жив!»: образы, нарративы и дискурсы народного восстания в Буркина-Фасо (2014-2015 гг.)
«Блэз Компаоре — мёртв, Томас Санкара — жив!»: образы, нарративы и дискурсы народного восстания в Буркина-Фасо (2014-2015 гг.) Филантропический обман
Филантропический обман Расстрел Белого дома и другие события 21 сентября — 5 октября 1993 года: документы и материалы Комиссии ГосДумы
Расстрел Белого дома и другие события 21 сентября — 5 октября 1993 года: документы и материалы Комиссии ГосДумы Трудно быть с богом
Трудно быть с богом
