
И.Р. Бёме
Изучение акустической сигнализации птиц — одна из наиболее важных проблем современной биологии. Изучение становления и развития видового репертуара в онтогенезе может стать тем критерием, который поможет заново и по-новому интерпретировать механизмы коммуникации птиц. Однако в литературе развитию акустической коммуникации уделено гораздо меньше внимания, чем дефинитивным акустическим системам.
К матуронатным птицам, по определению А.Н. Промптова (1956), относятся все зреловылупляющиеся, выводковые, птицы. К этим птицам принадлежат представители наиболее древних отрядов класса птиц, такие как страусоподобные, гагарообразные, поганкообразные, гусинообразные, куринообразные, журавлеобразные и пр. Однако птицы, имеющие имматуронатный тип развития, могут различаться по своему филогенетическому положению в системе класса.
Так, по птенцовому или полуптенцовому типу онтогенеза развиваются представители и таких древних отрядов, как аистообразные и пеликанообразные, и наиболее молодые группы птиц: попугаеобразные и воробьинообразные.
Тип онтогенеза акустических сигналов у матуронатных и имматуронатных птиц различается по целому ряду характеристик. Различия в голосе и сигналах птенцов и взрослых птиц сразу бросаются в глаза. Означает ли это, что их системы коммуникации различны? Какие изменения происходят с голосом в процессе развития птенца? До каких пор он остается ювенальным, а когда становится голосом взрослой птицы? Происходят ли эти изменения путем превращения птенцовых сигналов во взрослые, или они связаны с образованием совершенно новых типов звуков? Что такое ломка голоса, как и в какие сроки она происходит? Когда акустические сигналы приобретают индивидуальные и половые различия? Вот те основные вопросы, которые стоят перед исследователями, изучающими онтогенез вокализации птиц.
Онтогенез вокализации выводковых птиц
Для выводковых птиц изучение становления их звукового репертуара должно захватывать не только изучение вокализации птенцов после вылупления, но и включать формирование пренатальной вокализации. Пренатальный период развития звуковой системы птенцов играет большую роль в формировании слухового импринтинга и позволяет понять те процессы, которые поддерживают прочные связи птенцов в выводке между собой и взрослой птицей (матерью).
Развитие акустической сигнализации птиц в эмбриогенезе изучено очень слабо. Основное внимание было уделено «щелкающим» звукам эмбрионов, так как они легче всего регистрируются непосредственно перед вылуплением. В очень немногих, единичных работах делаются попытки рассмотреть коммуникацию насиживающей птицы
и эмбриона <> [<> — опущенные ссылки на литературу. Публикатор].
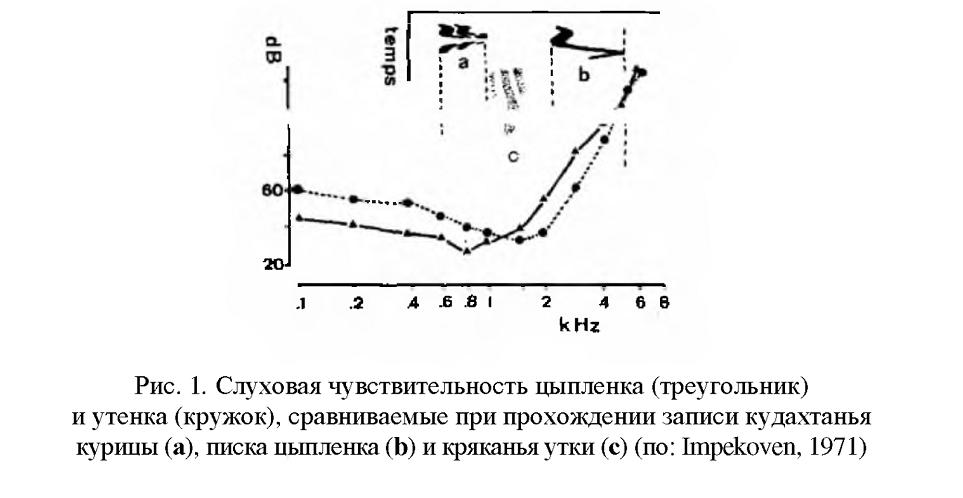
Как было показано <>, утята уже в яйце учатся различать голос матери, а их слуховой анализатор настроен на восприятие доминантной частоты ее сигналов (рис.1). Кроме того, во многих работах, проведенных на гусеобразных и других представителях выводковых птиц (нанду, куриных и др.), была показана важная роль пренатальной вокализации в синхронизации вылупления птенцов. Птенцы выводковых птиц к моменту вылупления из яйца обладают уже полным акустическим птенцовым репертуаром.
Развитие акустической сигнализации в пренатальном онтогенезе опосредованно связано со становлением легочного дыхания. Первые звуковые сигналы эмбрионы издают еще до выхода их в воздушную камеру яйца (Тихонов, 1975, 1977). По времени появления эти позывы соответствуют «спонтанному» дыханию эмбрионов, которое осуществляется за счет воздуха, находящегося в полости амниона. В этот же период формируется связь эмбриона и насиживающей птицы.
Звуковое общение широко используется выводковыми птицами, что имеет как предпосылку опережающее развитие слуховой системы в эмбриогенезе, по сравнению с развитием зрения. Оно обеспечивает максимальную чувствительность перед вылуплением в диапазонах частот, соответствующих основным энергетическим максимумам в звуковых сигналах родителей и собственной вокализации.
Диапазон воспринимаемых частот у птенцов пластинчатоклювых совпадает со спектральными характеристиками видоспецифических сигналов взрослых птиц, эффективных для соответствующих форм поведения, что имеет явное адаптивное значение. А именно, что видоспецифическая звуковая сигнализация между эмбрионами и взрослыми птицами обеспечивает синхронизацию вылупления выводка и поддержание его стабильности в дальнейшем (Тихонов, 1977).
Сроки включения звукоиздающих систем варьируют у представителей различных систематических групп. Первые сигналы эмбрионов следуют одиночными импульсами и разделены длительным временным интервалом (до 30-60 мин.). После прорыва эмбрионом подскорлуповой оболочки и его выхода в воздушную камеру яйца, звуковая активность эмбриона резко увеличивается. Некоторые авторы считают это увеличение активности индикатором перехода на истинно легочное дыхание (Тихонов, 1986). Интенсивность таких сигналов возрастает, но интервалы все равно достигают 20-30 минут. Проклёв скорлуповой оболочки сопровождается излучением писков, образующих серии из 2-3 импульсов.
Переход эмбрионов к дыханию атмосферным воздухом сопровождается ритмической организацией излучаемых звуковых сигналов. Определенные их категории имеют функциональное значение в процессе звукового общения эмбриона и насиживающей птицы (сигналы «дискомфорта», «комфорта»).
Таким образом, в пренатальном онтогенезе голос птиц развивается от единичных импульсов, не образующих серий, к ритмической последовательности, включающей большое количество структурных элементов. Параллельно в сигналах происходит усложнение и изменение форм частотной модуляции.
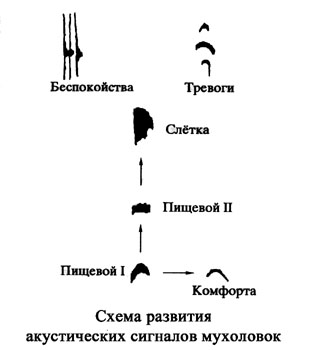
В большинстве случаев акустическая сигнализация птенцов выводковых птиц представлена пятью основными категориями сигналов, которые отражают как особенности внутреннего состояния птенца, так и его взаимоотношение с окружающей средой. Это сигналы дискомфорта, комфорта, ориентировочные звуки, сигналы тревоги и конфликтных ситуаций. В первые дни жизни птенцов сохраняются еще и » щелкающие» звуки, характерные для птенца на последних стадиях эмбрионального развития (Тихонов, 1986).
Основные типы позывов у выводковых птиц формируются независимо от условий, в которых развивались птицы. Только становление тревожных сигналов имеет ряд особенностей: здесь чрезвычайно велика роль птиц-родителей. Их поведение и сигнализация обеспечивают адекватное и своевременное формирование этих позывов у птенцов.
Таким образом, для пластинчатоклювых очень важно групповое обучение и сигнальная преемственность. Для развития голоса у уток и гусей были выделены следующие общие закономерности: в первые дни жизни (до 7-8 дней у разных видов) происходит повышение основной частоты акустических сигналов. Затем начинается медленное равномерное падение частоты, которое выходит на плато не позже, чем через 70 дней. Многие исследователи связывают такое изменение частотных характеристик сигналов с увеличением массы тела птенцов, а также с ростом и формированием трахеи и голосовых связок. Однако этот вопрос до конца не ясен, так как птенцы в возрасте 2-3 месяцев иногда могут издавать отдельные ювенальные высокочастотные сигналы <>.
Под ломкой голоса обычно подразумевают переход в онтогенезе от тонального ювенального сигнала к сонорному взрослому. Голос взрослого состоит из наложенных на ювенальный сигнал обертонов, шумовых структур и вибраций. Как показывают исследования на других видах птиц, ломка голоса затрагивает не все типы криков данного индивидуума и не все типы криков у разных полов в одинаковой мере. Изменения структуры отдельных элементов происходят в разное время.
В то же время полностью новые сигналы возникают позже. Ломка голоса позволяет постепенно перевести ювенальные части репертуара во взрослый. Этим консервативным способом развития происходит «открытие» новых форм сигналов взрослого репертуара (Бёме, 2009).
Если частотные характеристики сигналов претерпевают в ходе онтогенеза существенные изменения, то временные параметры и их синтаксис остаются более или менее постоянными. Число слогов (элементов) в сигнале в целом не изменяется в процессе онтогенеза. В сигналах трелевой структуры временные и ситуативные характеристики остаются постоянными.
Сигналы, используемые в критических ситуациях (бедствия, тревоги), переносятся без изменений во взрослый репертуар. В отношении развития других сигналов было показано, что развитие сигналов взрослых птиц происходит не только путем потери отдельных птенцовых криков, но и как модификация уже имеющихся сигналов. Кроме того, существуют сигналы, которые возникают в репертуаре уже после окончания раннего онтогенеза птенцов: это «роллен» серых гусей и » плач» белощекой казарки. Помимо этого, отмечается существование переходных звуков, причем не только между соседними типами сигналов.
Была рассчитана частота встречаемости различных переходных типов у птенцов белощекой казарки: только для двух типов она превышала 20%, в остальных случаях составляла 5-10%. Интересно, что переходные звуки несколько чаще встречались у ослабленных и больных птенцов <>.
В данных работах описан также процесс преобразования ювенального голоса во взрослый. У взрослых гусей и казарок звуки содержат большое количество нелинейных феноменов, из-за которых гармоническая структура звука остается замаскированной. Авторы показали, что по мере роста птенцы все чаще используют звуки с нелинейными феноменами; особенно заметно это становится после достижения птенцами 45-50-дневного возраста <>.
Для полувыводковых птиц, таких как представители чистиковых птиц, характерен как бы смешанный тип развития. Птенцы кайры после вылупления отвечают на крики родителей, которые они слышали еще в яйце, а птенцы гагарки имеют меньшую способность к распознаванию сигналов и часто ошибаются. Чем обусловлены такие различия у этих систематически близких видов? Кайры образуют плотные колонии на обрывах скал, где имеется дефицит мест для гнездования.
Для птенцов этого вида очень важным является способность опознавать своих родителей и отличать их от сотен других особей сразу после вылупления. В отличие от них, гагарки устраивают свои гнезда в небольших нишах или щелях, вылупившиеся птенцы в таких условиях имеют ограниченную возможность встретиться с другими взрослыми птицами, кроме родителей. Таким образом, у них, имеется достаточно времени, чтобы «выучить» голос своих родителей. На этом примере ясно видно действие отбора на два близких вида птиц. В ходе эволюции виды «вырабатывают» или «теряют» те или иные формы поведения в зависимости от экологических условий, в которых шло формирование вида.
Онтогенез вокализации птенцовых птиц
Перейдем теперь к рассмотрению типичных птенцовых, имматуронатных птиц. В ранних работах формирование вокализации через обучение традиционно рассматривалось как характерная черта видов, относимых к подотряду певчих птиц отряда воробьиных (подотряд Оscines) <>. Однако позже было доказано обучение и у представителей подотряда кричащих воробьиных (подотряд Сlamatores) <> [см. недавнее доказательство формирования через обучение песни птиц-звонарей Procnias sppp. Публикатор].
Развитие песни у птиц может идти самыми разными путями, и этот процесс отличается большим разнообразием у разных видов <>. Например, в подотряде кричащих воробьиных нормальная песня формируется даже при ранней изоляции или оглушении птенцов <>. У певчих же птиц птенцы для формирования нормальной видовой песни нуждаются во внешней модели <>.
Потребность в звуковой стимуляции — это единственная общая потребность для всех певчих птиц, тогда как конкретные модели обучения меняются у разных видов <>. У некоторых видов самцы могут сформировать песню при прослушивании ее магнитофонной записи, другие же виды нуждаются в живом учителе <>. Однако до сих пор не существует единого мнения о закономерностях развития песни в ходе онтогенеза (Бёме, 2006). Г.Н. Симкин (1981) выделяет два основных принципа становления вокального репертуара певчих птиц:
1) принцип активации и спонтанного саморазвертывания врожденных элементов песни;
2) принцип матричного копирования молодыми птицами особенностей напева старых самцов, иногда с элементами имитации отдельных звуков других видов.
Помимо этого была выявлена возможность «принудительного» экранирования и матричного копирования песни чужого вида (Симкин, 1981).
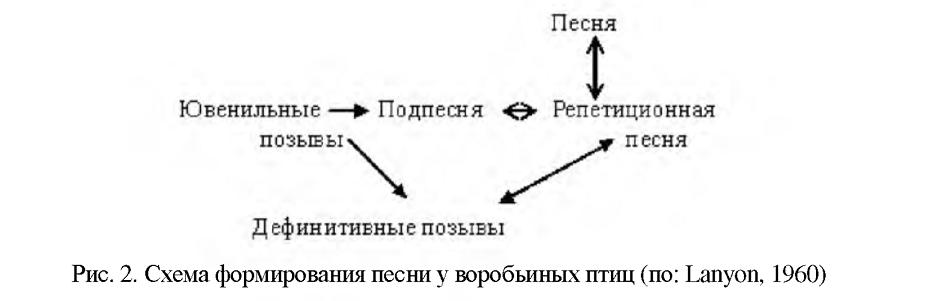
В процессе формирования песня проходит ряд последовательных стадий. Ланьон (Lanyon, 1960) предложил следующую схему развития песни воробьиных птиц (рис.2). Первые позывы выпрашивания пищи, а также птенцовое бормотание объединяются в подпесню, которую иногда называют «осенней подпесней» <>, так как этот процесс происходит осенью. Осенняя подпесня в течение нескольких месяцев претерпевает изменения: в нее птица включает отдельные фразы и колена видовой песни, которые молодой самец заимствует у взрослых птиц. Но на этой стадии «репетиционной песни» в ее состав могут входить позывы и песни других видов. Так, у зяблика в этот период отмечались заимствования от 60 видов, причем часть видов была африканского происхождения <>.
И, наконец, процесс формирования песни заканчивается созданием нормальной видовой песни (главных песенных мотивов), из которой исчезают все заимствования и другие добавочные элементы. Этот процесс Ноттебом <> назвал кристаллизацией песни. Данная схема затем была подтверждена работами других авторов <>.
У многих видов и групп воробьиных птиц песня в норме формируется в течение определенного периода времени, получившего название «критического» <>. Длительность критического периода различается у разных видов: у зяблика Fringilla coelebs он заканчивается к 14 месяцу жизни <>, у зебровой амадины Таеторуgia guttata ограничивается возрастом от 35 до 65 дней <>, у болотного крапивника Cistothorus
Песня птиц включает, вероятно, некоторые врожденные, закрепленные генетически элементы, которые передаются птицам вне зависимости от их слухового опыта. Так, песня канарейки богата различными элементами, и эти элементы канарейки способны выучивать в течение всей жизни. Поэтому песня канарейки расценивалась как полностью формирующаяся в процессе научения. Однако недавние исследования <> показали, что в репертуаре домашней канарейки имеются определенные фразы (фразы А, «сексуальные слоги»), которые имеются и у диких канареек. Эти фразы вызывают у самок позу приглашения к копуляции и чаще встречаются у тех самцов, которые занимают верхние ступени иерархической лестницы. Фраза А, извлеченная из репертуара дикой канарейки, была в той же степени реактогенна для самок домашних канареек, что и фраза собственных самцов.
Кроме того было показано, что даже воспитанные в изоляции от самцов самки отдают предпочтения песне, в которой присутствует фраза А <>. Все это заставляет предположить, что песня канареек включает некоторые врожденные, закрепленные генетически элементы, которые передаются птицам вне зависимости от их слухового опыта. Это в равной степени относится как к самцам, так и к самкам.
Обучаемость птиц новым фразам и структурам песни может быть типичным примером «принудительной индукции» (Симкин, 1976) и являться следствием обедненности звуковой среды при клеточном содержании птиц. Косвенным доказательством этого может служить тот факт, что молодых самцов канареек для обучения новым строфам песни помещают в изоляцию в темной комнате, а в качестве единственного звукового символа оставляют эталонную песню.
Таким образом, остается справедливой идея о наследственной детерминации песен и позывов. Наследственная основа песни может быть выражена для большого числа видов лишь в отношении общих способностей воспроизведения звуков, общей видовой характеристики песни и темпа исполнения. Но нормальная песня без влияния старых певцов у подавляющего большинства видов сложиться не может. У выкормышей, воспитанных без взрослых птиц своего вида, складывается аберрантное пение, часто отличное от видового.
Из всего изложенного можно сделать следующие выводы. Развитие акустических сигналов у выводковых и птенцовых птиц отличается по целому ряду признаков.
1. У матуронатных птиц первые акустические сигналы формируются на последних стадиях пренатального развития.
2. Для птиц с полуптенцовым типом развития (аистообразные, соколообразные) характерно долгое развитие птенцов до приобретения способности к полету. Акустические сигналы не совпадают у птенцов и у взрослых птиц. При переходе к самостоятельной жизни у птенцов формируется видовой репертуар акустических сигналов. Для аистов характерно наличие голосовых сигналов только в первые 3 недели жизни птенцов, у взрослых птиц присутствуют только механические сигналы (щелчки клювом)[1].
3. У имматуронатных птиц, относящихся к отрядам воробьинообразных и попугаеобразных, выявлена зависимость формирования сигналов от научения, хотя и у них многие позывы генетически фиксированы и могут входить в репертуар без предварительного научения.
Для представителей этих групп характерна способность к имитации, не отмеченная у представителей других отрядов, а также формирование локальных диалектов.
4. И у имматуронатных (воробьинообразные), и у матуронатных (журавлеобразные, курообразные и гусеобразные) птиц выявлена «ломка» голоса в ходе онтогенеза акустического репертуара. Однако у матуронатных птиц она касается основных позывов, а у воробьиных — различных стадий формирования песни.
Работа поддержана грантами РФФИ 07-04-00609-а и 11-04-00062-а.
Литература
Бёме И.Р., 2006. Формирование вокализации воробьиноообразных птиц в онтогенезе. Современное состояние проблемы // Ж. общ. биологии, т.67, № 4. — С.280-297.
Бёме И.Р., 2009. Формирование акустического репертуара Гусеобразных в онтогенезе // Казарка, т.12 (1). — С.33-46
Голубева Т.Б., 2006. Сенсорное обеспечение ранней акустической коммуникации выводковых и птенцовых птиц // Орнитол. исследования в Сев. Евразии: Тез. XII Международн. орнитол. конф. Сев. Евразии. — Ставрополь: Изд-во Ставроп. ун-та. — С.144-146.
Промптов А.Н., 1956. Очерки по проблеме развития биологической адаптации поведения воробьиных птиц. — М.-Л.: Изд-во АН СССР. — 310 с.
Симкин Г.Н., 1976. Опыт разработки функциональной классификации акустических сигналов у птиц // Групповое поведение животных.- М.: Наука.- С.338-341.
Симкин Г.Н., 1981. Песня восточного соловья как акустический маркер групповых и популяционных структур // Орнитология. — М.: Изд-во Моск. ун-та, вып.16. — С.73-83.
Тихонов А.В., 1975. Звуковая сигнализация у птенцов выводковых птиц // Мат-лы Всесоюз. конф. по миграциям птиц, ч.2.- М.- С.165.
Тихонов А.В., 1977. Звуковое общение эмбрионов и насиживающей самки у выводковых птиц // Тезисы докл. VII Всесоюз. орнитол. конф., ч.2.- Киев: Наук. думка.- С.210.
Тихонов А.В., 1986. Акустическая сигнализация и экология поведения птиц. М.: Изд-во МГУ. — 240 с.
Источник: Труды Мензбировского орнитологического общества. Т.2, 2011 г.
Примечания
[1] У взрослых черных аистов есть очень громкие, звонкие свистовые сигналы (см.: 8пода, Ретп8, 1998; Казаков и др., 2004; и др.) (Прим. ред.).
Рекомендуем прочесть
 Распродажа России
Распродажа России Куба не подошла для «твиттерной революции»
Куба не подошла для «твиттерной революции» Тропарёвский конфликт
Тропарёвский конфликт Сжечь ведьму! Или российские доброхоты о мигрантах.
Сжечь ведьму! Или российские доброхоты о мигрантах. Как фармкомпании коррумпируют науку. Ч.2.
Как фармкомпании коррумпируют науку. Ч.2. Деньги воспитывают целеустремлённых эгоистов
Деньги воспитывают целеустремлённых эгоистов Евграф Каленьевич Дулуман. Существовал ли Иисус Христос?
Евграф Каленьевич Дулуман. Существовал ли Иисус Христос? Подъём религиозной правой в США 1970-х-80-х гг.
Подъём религиозной правой в США 1970-х-80-х гг.
