

Слева: большой султанский дятел (Chrysocolaptes guttacristatus). Справа: индо-малайский золотоспинный дятел (Dinopium javanense). Источник.
Виды, непонятно почему слишком сходные друг с другом
Содержание
В связи с открытием олингито, которого долгое время путали с довольно-таки не родственным, но очень схожим зверем олинго [оригинал], я вспомнил вот что. Среди птиц есть пары видов, филогенетически совершенно не близких, разного размера и пр., однако с окраской, совпадающей до мелких подробностей, что структуры, что наполнения.
Особенно их много у дятлов: см. статью Martin L. Cody (1969). Скажем, дятлы из юго-восточной Азии Dinopium javanense и Chrysocolaptes lucidus (очень разного размера, у первого вида лапа 3-хпалая, у 2го – 4х). Или из пёстрых дятлов очень сходно окрашены, но разного размера волосатый Dendrocopos villosus и пушистопёрый Dendrocopos pubescens в Северной Америке. Недавно открыли ещё одну: было показано, что т.н. шлемоносная желна Dryocopos galeatus из «атлантических лесов» Юго-Восточной Бразилии относится к другому роду (и трибе) — целеусам, и должна быть перемещена туда.
«Хотя по окраске она более сходна с Dryocopus, форма клюва, ноздрей и некоторые мелкие детали оперения (например, белые верхние кроющие перья хвоста) напоминают Celeus. Так, клюв шлемоносной желны более тонок, поэтому она редко долбит, — это объединяет ее с дятлами-целеусами, но не характерно для активно разрушающих древесину в поисках пропитания желн — обладателей мощного клюва».
Источник элементы.ру
Впрочем, высказанное предположение, что «мелкие дятлы мимикрируют под крупных, чтобы уменьшить их агрессию» — чисто умозрительное. Оно давно высказано в двух вариантах — этом и противоположном у Cody: мол, разные виды дятлов потому и похожи, потому что между ними существует межвидовая территориальность. Везде, где их проверяли специально, они не подтверждались.
Таких пар неродственных видов с совпадающей окраской у птиц известно до десятка (у дневных хищных, кукушек и пр.), причём это если отбросить случаи мимикрии. Вопрос к биологам – какой из факторов эволюции может создать и поддерживать столь полное сходство окраски, притом что об экологической близости, сходстве размеров, биологии и р. речь тоже не идёт?

Олингито
Или это явление специального объяснения не требует, это просто «хвост» распределения, описывающего структуру разнообразия, скажем тех же гипербол Виллиса? То есть если в крупной группе типа дятлов около 400 видов, некое интегральное сходство их между собой по окраске распределено гиперболически, ну 1% видов будут совпадать по окраске очень точно. Какая из интерпретаций вам кажется более вероятной?
Правила Виллиса…
Для небиологов рассказу про правила Виллиса поподробней, они того заслуживают.
Ранговые распределения Виллиса замечательны тем, что раз за разом воспроизводятся во всех таксонах живых организмах, созданных трудом систематика, независимо от используемого подхода в филогенетических реконструкциях и независимо от тенденций чисто систематических («дробители» vs «объединители»)
Правило Виллиса состоит в следующем. Если для семейства или отряда мы по оси абсцисс отложим число видов в роде, по оси ординат – число родов с данным числом видов, то получим гиперболу. Математически точно такое же правило «ранг-размер» описывает закономерности урбанизации территории, именно связь между рангом города по людности, начиная с самого крупного, и числом городов данного ранга на территории, и много ещё что («законы Ципфа»).
В каждом надтаксоне ранга отряда-семейства будет 1-2 рода очень больших, вроде (у птиц) родов Larus, Columba или Turdus, и много родов с 1-2 видами. В более удобных логарифмических координатах гиперболы Виллиса превращаются в прямые. То же верно и для прочих таксонов, скажем, для распределения семейств с разным числом родов в классе (скажем, в тех группах животных, определительные ключи по которым обрабатывал А.А.Поздняков, статью которого «О значении правила Виллиса для таксономии» в Журнале общей биологии, т.66, №4, 2005 года перескажу ниже).
«В 1918 году английский ботаник Джон Кристофер Виллис, изучая свой «Словарь цветковых», обнаружил, что в каждом крупном семействе более трети родов — одновидовые. Странно: роды для того и вводятся, чтобы объединять сходные виды, почему же так много одновидовых? Всего в его Словаре описан 12571 род, из которых 4853 (т.е. 38,6%) содержат по одному виду. Двухвидовых родов оказалось 1632 (12,9%), трехвидовых — 921 (7,3%) и так далее — чем больше видов в роде, тем меньше таких родов. Точнее, распределение родов по числу входящих в них видов грубо выражается убывающей кривой, похожей на гиперболу.
Зато в каждом семействе оказался род-гигант (более сотни видов), а два рода цветковых (астрагал и крестовник) содержали более полутора тысяч видов каждый. То же самое Виллис обнаружил и на животных. Это уж совсем странно — почему не разбить их на меньшие роды? Однако ничего не выйдет: пробовали, разбиение оказывалось несуразным и отвергалось… Хвосты этих кривых нерегулярны, а середина всегда содержит горб, поэтому их лучше называть квази-гиперболами.
Они обладают общим инвариантом: 35-40% родов имеет по одному виду, 12-17% родов — по два вида, 7-11% — по три вида и т.д., тогда как основная масса видов состоит в немногих крупных родах, и непременно найдется род-гигант.
Сходным оказалось распределение родов по семействам: так, цветковых ныне насчитывают 474 семейства, из которых 170 — однородовые (36%)…. Высказывалось мнение, что кривые Виллиса являют собой не биологический, а психологический феномен: отражают не структуру биологического разнообразия, а способ классифицирования. Однако квазигиперболы обычны и в тех отраслях знания, где о влиянии психики исследователей говорить невозможно: были открыты аналогичные виллисовым распределения слов по их длине, букв и слов по их встречаемости в тексте, двигателей по их мощности, химических элементов по их обилию в природе, мутаций по их частоте и т.д. — все они тоже распределены приблизительно гиперболически…
Виллис высказал в 1922 году уверенность, что нашел общий закон эволюции: распределение видов по родам есть итог процесса, определяемого «видовой мутацией», т.е. появления (с заданной частотой) нового вида, и «родовой мутацией», т.е. распада вида (тоже с заданной частотой) на два рода. Это было в духе де-Фриза.
Модель данного процесса предложил приятель Виллиса математик Гаролд Юл в 1924 году: это ветвящийся процесс появления видов и родов…. С появлением теории фракталов оказалось, что ветвящийся процесс хорошо описывается как фрактальный рост. В частности, в логарифмическом масштабе гипербола предстает как прямая, тангенс угла наклона которой есть размерность фрактала; для модели Юла она равна отношению частоты «видовой мутации» к частоте «родовой мутации».» (из «Элементов эволюционной диатропики» Ю.И.Чайковского. Хоть писания данного автора требуют двойной предосторожности, бо содержат много вранья, тут с фактологией всё в порядке).
Другой важный момент – аналогичные гиперболы Виллиса получаются не только для естественной системы таксона, построенной так, чтобы дерево классификации отражало родство, но и для дерева тез-антитез определительных ключей, построенных по чисто прагматическим критериям удобства, точности и однозначности определений. Другой очевидный момент – гиперболы Виллиса отражают продукт эволюционного процесса, в то время как сам процесс эволюции описывается иным – колоколообразным распределением с направленным сдвигом «колокола» под действием естественного отбора.
Это опять же всеобщее правило для ветвящихся процессов с памятью. Скажем, оценки студентов университета и другие параметры эффективности обучения распределены более или менее колоколообразно. Однако если мы посмотрим научную продуктивность через 10 лет, то окажется что она распределена по Виллису. Аналогичным образом распределены доходы граждан в конкурентной социальной среде при капитализме, но в солидарной среде советского социализма они были распределены колоколобразно (п.5).
То есть гипербола получается из колокола, когда образующие его «точки» — особи, меняющиеся взад-вперёд в признаковом пространстве, заданном по оси абсцисс, конкурируют друг с другом. Конкурентоспособность есть функция положения в признаковом пространстве, выжить и размножиться могут «точки» с конкурентоспособностью не ниже некоторой, уровень которой и задаёт «пропуск в будущее». «Следует обратить внимание на следующий момент. Например, если взять достаточно представительную выборку черепов какого-нибудь вида млекопитающего и выяснить распределение по такому параметру, как длина черепа, то мы должны получить нормальное распределение, представленное колоколообразной кривой. Но, если мы возьмем по одному черепу каждого вида млекопитающего и выясним распределение по этому же параметру, то получим ранговое распределение» (А.А.Поздняков, ibid.).
Поскольку распределения Виллиса инвариантны для разных таксонов, филогенетических теорий и систематических практик, понятное дело, их подняли на щит номогенетики – вот, мол, имманентные законы структурирования биоразнообразия (ибо, более выспренне, развития биологической формы), которые не укладываются в ателические механизмы эволюции. Поэтому реконструкция порождающих механизмов для правил Виллиса – вещь крайне актуальная, что делает А.А.Поздняков «Значениях правила Виллиса для таксономии».
… И их порождающие механизмы
Каковы существующие предположения по этому поводу? Первое принадлежит Э.Майру (1971). Процесс видообразования независим от дивергенции по признакам (степень которой определяет, какие из новых видов мы выделим в отдельный род, семейство и пр.). Это означает случайный характер появления новых видов = случайный характер распределения признаков, с помощью которых дифференцируются виды. Переволдя эволюционную терминологию (видообразование, дивергенция) на язык таксономии это значит отсутствие корреляции между объёмом родов и признаками, с помощью которых дифференцируются роды. Количество видов отражает объём рода, и ранговое распределение Виллиса – это прежде всего распределение родов по их объёму (ну и других таксонов, выше рангом, с соответствующими подтаксонами: семейства – рода, классы – семейства и пр.).
Альтернативная гипотеза предполагает сильную корреляцию между объёмом и характеристикой родов. Она предложена А.В.Свиридовым (1994), основывавшимся на анализе определительных ключей. Согласно ему, признаки, дифференцирующие рода друг от друга, отражают гомологические ряды изменчивости Вавилова (с которым, по его мнению, связан закон Виллиса), и поэтому дифференцируются не случайно. «Гомологическая изменчивость в стволах пропорциональна расстоянию между ними».
Для проверки, какое предположение верно, А.А.Поздняков обработал определительные ключи, тезы и антитезы с признаками, составленные для различных групп организмов, включающих не менее 50 таксонов. Результаты приведены в табл.1 (ключи для определения родов), 1-6 – группы организмов. Использовали следующие статистики: F/T – количество дифференциальных признаков, приходящихся на 1 таксон, в определенном смысле это мера морфологического разнообразия последнего. Чем ниже F/T, тем меньше разнообразных признаков задействовано для различения соответствующей группы. S/T – среднее количество дифференциальных признаков, которые используются для различения данных таксонов.
То есть увеличение данного показателя означает либо что таксоны различимы плохо и для различения требуется привлекать множество признаков, либо, наоборот – отлично различаются по очень многим.
Так вот, распределение признаков по числу ссылок на них в исследованных группах оказывается близким к ранговому, в логарифмических координатах – к линейному (рис.1). Распределение семейств по количеству родов также ближе к ранговому (в логарифмических координатах – линейному), чем к нормальному (рис.2). Интересно, если исходить из степени приближения указанных распределений к линейному, то оказывается, что для наземных позвоночных, исключая амфибий, и растений завышено количество семейств, включающих один род, тогда как у амфибий завышено количество семейств, включающих два рода. Хотя вполне может быть, что сравнительно большое количество семейств с двумя родами у последних отражает таксономические реалии этого класса позвоночных.
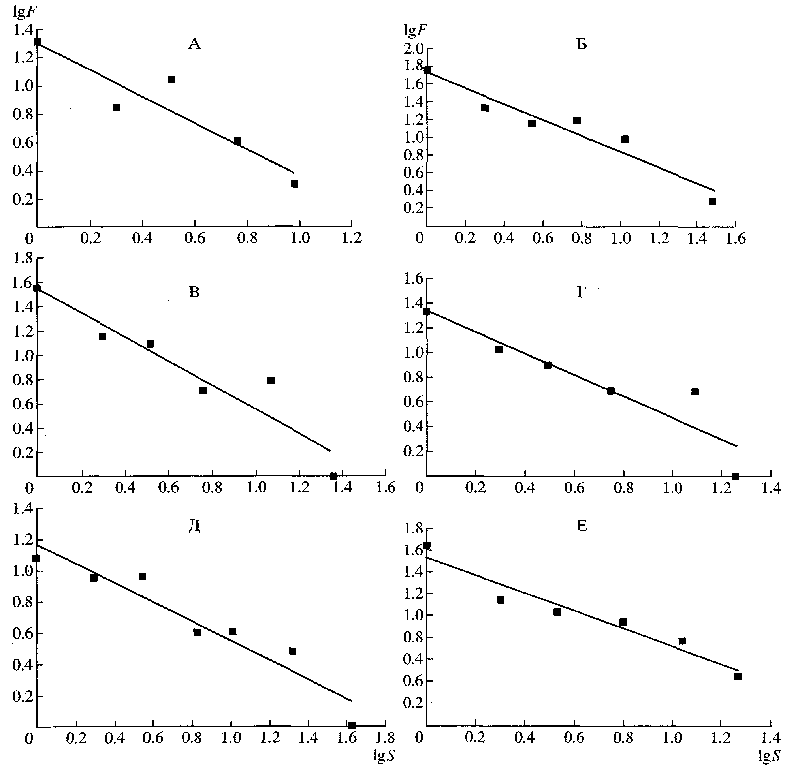
Рисунок 1.. Распределение признаков по количеству ссылок на них в ключах для определения родов разных групп организмов: lgS — логарифм количества ссылок на признаки; lgF — логарифм количества признаков; А — Nematoda; Б — Curculionidae; В — Syrphidae; Г — Cyprinidae; Д — Cruciferae; E — Umbelliferae.
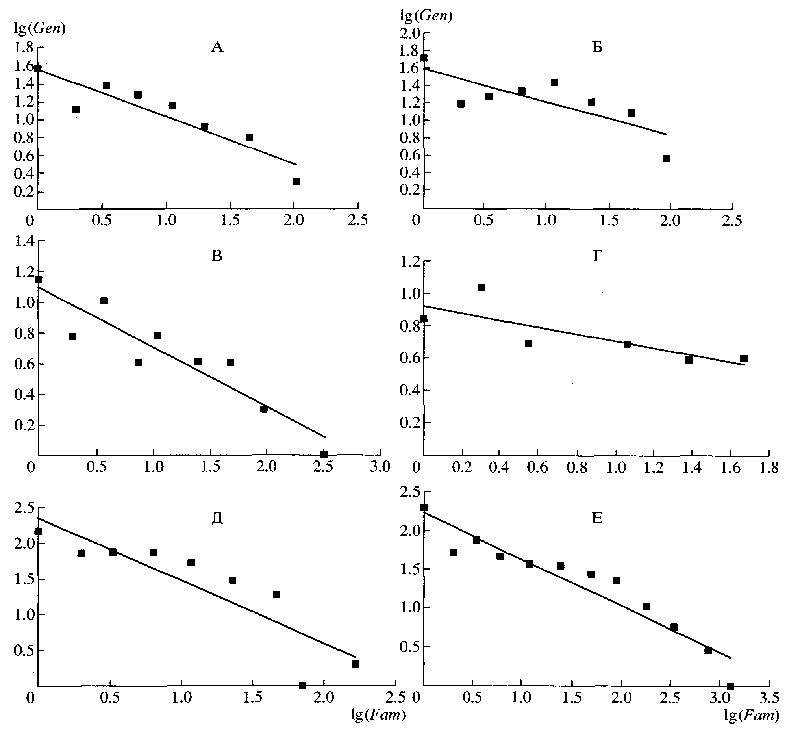
Рисунок 2. Распределение семейств по количеству родов: А — млекопитающие (по: Walker, 1968); Б — птицы (по: Беме, Флинт, 1994); В — пресмыкающиеся (по: Ананьева и др., 1988); Г — земноводные (по: Ананьева и др., 1988); Д — рыбы (по: Решетников и др., 1989); Е — покрытосеменные (по: Тахтаджян, 1987).
Данный результат «в первом приближении» подтверждает гипотезу Свиридова, а не Майра. Распределение семейств по количеству родов также ближе к ранговому (линейному в логарифмических координатах), чем к нормальному. Причём, если исходит из степени приближения таких распределений к линейному, то оказывается, у наземных позвоночных, исключая амфибий, и высших растений, завышено количество семейств, включающих 1 род, тогда как у амфибий – число семейств, включающих 2 рода.
Результаты анализа ключей для определения семейств см.табл.2. В них использовано больше признаков (F/T обычно >1, в табл.1 – меньше), что также согласуется с мнением Свиридова, ибо на уровне семейств эффекты гомологических рядов выражены хуже, чем на уровне родов. Однако распределение признаков по числу ссылок на них в ключах для определения семейств точно также близко к ранговому (в логарифмических координатах – к линейному, рис.3). То есть на уровне семейств правило Виллиса соблюдается не хуже, чем на уровне родов; следовательно, оно причинно не связано с гомологическими рядами Вавилова, иначе на уровне семейств мы бы видели больший отход от рангового распределения к нормальному.
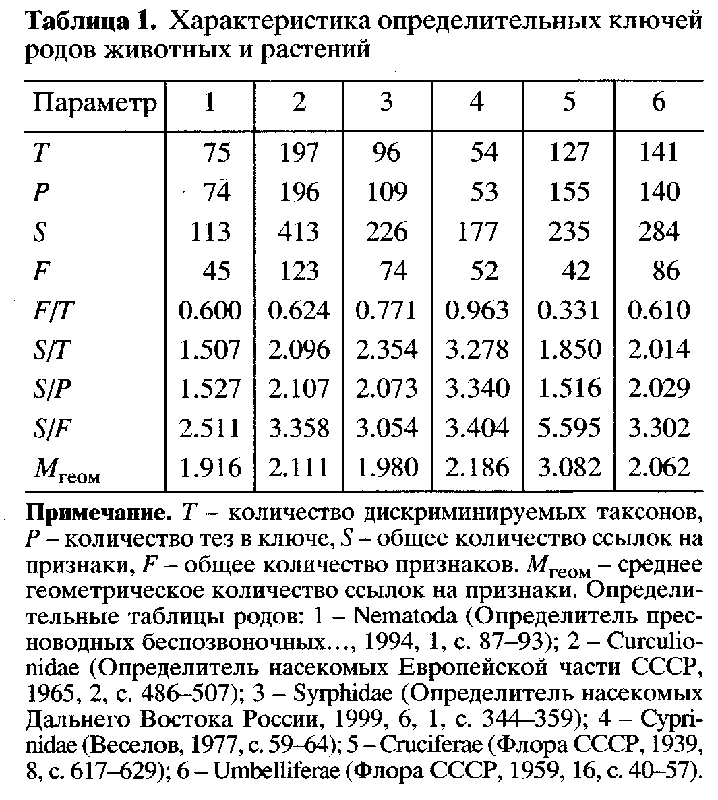
Таблица 1

Таблица 2
Итак, в определительных ключах (также как в результатах работы таксономиста) на разных уровнях классификации распределения таксонов по их объёму и дифференциальных признаков по частоте использования друг от друга являются ранговыми, т.е. соответствующими закону Виллиса. С математической точки зрения, то что распределено по нормальному закону, характеризуется максимальной энтропией или максимальной неупорядоченностью. Его противоположность — распределение по ранговому закону, — отражает максимальную упорядоченность.
Важно подчеркнуть, что данный способ упорядочивания биологического разнообразия поскольку инвариантен на всех уровнях и во всех формах последнего, отражает некую биологически содержательную теорию его порождения, причём несводимую к конкретным факторам эволюции, особенностям адаптивных зон и пр. отдельных таксонов, рассматриваемых в составе соответствующих групп (то, чем нагружены филогенетические сценарии, что исключительно интересно для каждого из них в отдельности, но скрывает общий инвариант у всех них вместе). Увы, о характере этой теории (отражающей механизм порождения Естественной системы, если высоким штилем) мы можем пока только гадать или расписываться в своём бессилии. См. удручающие попытки того и другого во введении в пересказе автора статьи.
Поскольку начинать надо с самого простого предположения, то, в общем-то, для порождения ранговых распределений достаточно одного лишь наличия памяти о прошлом состоянии системы, которое будет характерно в том числе и для ветвящихся процессов (или, по А.С.Раутиану, инерция биологической организации, изменимой отбором по разным направлениям с разной скоростью и разным риском вымирания, увеличивающимся пропорционально, с одной стороны, скорости изменений под действием отбора, с другой – «лёгкости» или, напротив, «опасности» изменений по данному направлению). «Так, согласно математическим моделям, ранговые распределения возникают при наложении двух ветвящихся процессов, например, возникновения видов и вымирания родов (Yule, 1924 – который собственно, первым и исследовал свойства ранговых распределений).
Или когда видообразование и вымирание видов идут случайно и независимо друг от друга, а возникновение новых родов происходит в результате дробления вымирающих родов. Я, как фанат биологической концепции вида и номогенетик, предложил бы другой механизм. Видообразование – процесс автоматический, идущий с более – менее постоянной скоростью, и связанный с тем, что разные географические популяции вида на градиенте «центр-периферия ареала» в периоды роста численности, «волн жизни» интенсивно дифференцируются и дивергируют друг от друга по признакам (оставаясь связными и именно из-за интенсивной сортировки особей по поведенческим потенциям между ними[1]).
А в периоды спадов, депрессий численности крайние популяции, самые сильно дивергировавшие, из-за прекращения подтока особей «обламываются» с общего градиента обмена особей внутри материнского вида, формируют свою замкнутую мини-систему обмена особями внутри только своего изолята. И по мере пребывания в изоляции вне подтока особей с основной части ареала увеличивают обособленность собственной «малой» популяционной системы от его «крупной» — пока не превысят видовой уровень обособленности (если до этого времени не начнётся новая волна жизни, сливающая обосабливающийся изолят в материнской формой).
Но образование нового вида (с т.з. биологической концепции, как обособленной популяционной системы) только полдела, дальше надо нишу найти и встроиться в сообщество. Ниш, соответствующих исходной морфологии (в целом предельно близкой к таковой исходного вида, ведь отпочковавшаяся форма отличается от неё только признаками, значимыми для «встраивания» особей в популяционные системы, но не факт, что признаками, нужными для приспособления к среде), на месте появления может не быть или ёмкость их может быть недостаточна. Тогда, чтобы выжить (поддерживать минимальную жизнеспособную численность популяции в условиях нового ареала, когда на подток рассчитывать уже не приходит, от исходной системы обмена особями новая форма «отключена») новому виду придётся эволюционировать и по приспособительным признакам тоже, всё дальше отеляясь от материнской формы. Тем дальше, чем больше разнятся доступная ниша в месте рождения нового вида и «подходящая» ему на основе первоначальной морфологии.
Виды, наиболее уклонившиеся от материнских в процессе такого рода адаптации, мы выделяем в отдельный род, когда степень несходства перейдёт некий предел, свой для каждой группы. То есть на «автоматический» процесс порождения новых видов (=политомического ветвления дерева) накладывается требование приспособления каждой полученной «ветки» к ближайшей подходящей нише, что «вытягивает» их в разной степени («вытягивание» означает анагенетический компонент дерева – степень дивергенции, отражающуюся в т.ч. в рангах таксонов). Будет или нет «рядом» с «родившимся» видов подходящая ниша и насколько она подходящая («как сильно надо меняться») тоже случайные величины. Их наложение на «автоматический» процесс видообразования даёт то самое ранговое распределение, только с сильно более содержательной биологической интерпретацией.
Это значит, что существующую филогенетическую систематику и кладистику можно рассматривать как попытки восстановить ход таксоногенеза в некой нерепрезентативно выделенной «части ветвей» дерева группы, но не как приближение к естественной системе всей этой последней. И действительно, посейчас отсутствует синхрония между появлением инноваций, по которым восстанавливается таксоногенез, и самими этими инновациями. Следующий минус касается эволюционной теории – доминирующие сейчас представления о способах появления инновации могут быть далеки от реальности (см. о левом и правом крыле дарвинизма, сейчас доминируют вторые, первые явно умалены, а из общих соображений нужна некая гармония тех и других, ведущая к новому синтезу). Наконец, филогенетические модели на низших уровнях таксономической иерархии получаются сильно более противоречивыми чем на высших, тогда как должно быть наоборот.
Это интересная проблема, которую стоит рассмотреть подробнее. Как уже говорилось, количество дифференциальных признаков на уровне родов меньше, чем на уровне семейств, а там, меньше, чем на уровне отрядов. В последнем случае F/T=2,75-2,88-2,83 для млекопитающих (разные ключи), 1,74 для птиц (ключ по Кузнецову, 1974; 27 отрядов 47 признаков), 2,44 (Иванов-Штегман: 18 отрядов 44 признака), для рыб – 2,4. При этом дискриминация семейств в отрядах и родов в семействе проводится с использованием разных признаков (это нетривиальная деталь к вполне тривиальному суждению, что если объёмы отрядов выше, чем входящих в них семейств, то и разнообразие должно быть выше).
«Например, при сравнении ключей для определения семейств Coleoptera и родов Curculionidae выясняется, что всего использовано 194 признака, из которых общими являются 27 (13.9%), а в случае Diptera и Syrphidae из 232 признаков общими являются только 9 (3.9%).
Получается, что дискриминация семейств в отрядах и родов в семействах проводится с использованием разных признаков. Так как в современных эволюционных моделях дарвиновской традиции отсутствуют формальные запреты на изменение (ограничение) разнообразия в процессе эволюции, то при использовании различного набора признаков при дискриминации таксонов различного уровня следовало бы ожидать примерно одинаковой величины показателя F/T на разных таксономических уровнях».
Однако в современных эволюционных теориях отсутствуют формальные запреты на изменение (ограничение) разнообразия в процессе эволюции, поэтому при использовании различного набора признаков при дискриминации таксонов различного уровня следовало бы ожидать примерно одинаковой величины F/T на разном таксономическом уровне. Следовательно, большая трудность построения филогении низших таксонов объективна, она связана с их меньшим морфологическим разнообразием в сравнении с высшими таксонами, что выражается в недостатке дифференциальных признаков.
«Логически можно предположить наличие двух способов формирования фрактала. Во-первых, путем дробления целого на последовательно уменьшающиеся части. Во-вторых, путем «умножения» единиц с образованием более крупных частей. Скорее всего, оба эти способа существуют в природе, но их описание возможно с помощью разных понятийных аппаратов, причем, видимо, дополнительных друг другу. Исторически признание наличия второго способа формирования Естественной системы связывается с «Происхождением видов» Чарльза Дарвина, причем на основе «умножения» видов с формированием таксонов более высокого ранга. Наличие первого способа формирования Естественной системы – путем дробления таксонов на подтаксоны – фактически провозглашено Вилли Хеннигом в «Основаниях теории филогенетической систематики».
С моей точки зрения, второй процесс – увеличение количества единиц Естественной системы – должен быть связан с образованием новых родов, или таксонообразованием. Первый процесс следует рассматривать с типологической позиции как изменение содержания интенсионала при таксонообразовании.

Шлемоносная (А) и полосатая (В) желны и королевский дятел (С). Источник
В связи с вышесказанным возникает проблема реальности Естественной системы и ее частей. Разумеется, Естественная система реальна как целое, как фрактал. Таксон родового ранга реален как универсальная единица Системы. Высшие (надродовые) таксоны являются фрагментами, частями Естественной системы и с определенной точки зрения они тоже реальны, как, например, реальны части лица. Однако с этой же точки зрения они не являются ни целостностями, ни имеют индивидных свойств и потому не обладают сущностью.
Однако проблема сущности слишком сложна и ее обсуждение требует отдельной статьи.
Следующим основным свойством фракталов является их дробная размерность, которая превышает их топологическую размерность. Биологические классификации, которые чаще всего представляют как список таксонов, расположенный в определенном порядке, имеют в этом случае топологическую размерность, равную единице. Одномерную топологию имеют классификации и в случае представления их в виде дендрограммы. Двумерную топологию имеют биологические классификации, выраженные в форме таблицы (Соболев, 1914; Догель, 1923; Шимкевич, 1929; Васильева, 1985), которая находит определенные теоретические обоснования (Васильева, 1987, 1990, 1993)».
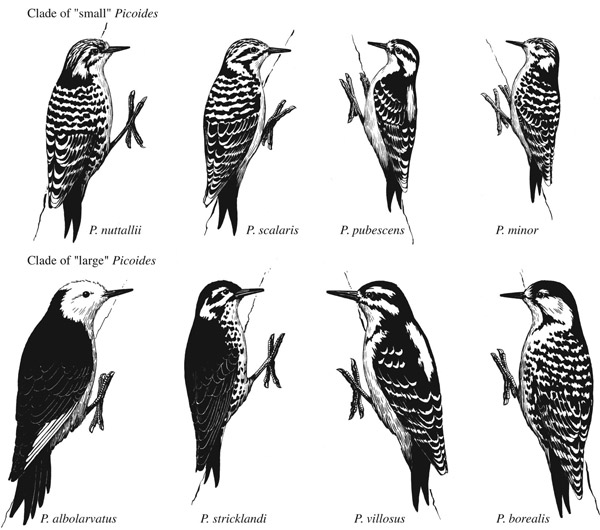
Мелкие (вверху) и крупные (внизу) виды пёстрых дятлов Северной Америки; обратите внимание на сходство в окраске villosus и pubescens. Правда, minor здесь лишний — это евразиатский вид, близкий к белоспинному дятлу D.leucotos. Источник
Заключение
Поскольку кривая Виллиса отражает структуру биоразнообразия, с её помощью можно делать прогнозы относительно «степени естественности» системы примерно того же рода, что делаются с помощью правила «ранг-размер» (это, собственно, то же самое правило), относительно избытка/недостатка на данной территории городских поселений разного ранга и, соответственно, что они в следующий период времени будут делать, расти или стагнировать. Правило Виллиса позволяет оценить приближенность существующей классификации группы, таксономической и мерономической (мерон – это класс частей тела; сравнением однотипных частей организмов, составляющих каждую группу, мы извлекаем те самые дифференциальные признаки, которыми различаем входящие в неё таксоны) к «естественной системе». Что даёт аргумент в вечном споре «дробителей» с «объединителями» что относительно дифференцируемых таксонов, что относительно дифференцирующих признаков. И, среди прочего, это означает, что профессия систематика ещё долго будет насущной, так как исследованные системы «движутся в направлении естественности», хотя в каждой из групп есть показатели, «портящие картину».
Отсюда же следуют представления об ограниченности изменчивости и таксономическом ранжировании дифференциальных признаков, так что естественная система представляет собой фрактал. Упорядоченность, т.е. структура биологического разнообразия, независима от порождающей его филогенетической истории, исследования того и другого оказываются взаимно-дополнительными.
Примечания
Рекомендуем прочесть
 «Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве
«Урбанистика» — имитационная игра по изучению общественного мнения в градостроительстве COVID-19 вскрывает правду
COVID-19 вскрывает правду Ревень Rheum palaestinum, самоорошающееся растение пустынь Израиля
Ревень Rheum palaestinum, самоорошающееся растение пустынь Израиля Анатомия кредитного кризиса. Взлет и падение цен на фермерские земли в США в 1920-е годы
Анатомия кредитного кризиса. Взлет и падение цен на фермерские земли в США в 1920-е годы Восприятие шумов
Восприятие шумов Культ книги в РККА: как красноармеец стал главным пропагандистом грамоты в стране
Культ книги в РККА: как красноармеец стал главным пропагандистом грамоты в стране Вибрационные сигналы в коммуникации животных
Вибрационные сигналы в коммуникации животных Марихуановое наступление продолжается
Марихуановое наступление продолжается
