

Резюме. Поведенческая сложность — понятие, описывающее, как поведение животного распадается на единицы, а те составляются из их базовых элементов (телодвижений и прочих моторных актов). [Понимание сложности включает в себя подбор метода объективного описания единиц, на которые оно распадается естественным образом, позволяющего или установить их на чисто структурной основе, охарактеризовав форму каждого, с дифференциацией друг от друга и от «фона» — либо показать невозможность подобного разделения на той же самой выборке данных. Здесь Шлейдт этой проблемы почти не касается, успешно решив её в работах 1980-х гг. Прим.публикатора] Чтобы понять её, нужно сперва определить сценарий, в рамках которого реализовано данное поведение: пространство и время активности животного [отслеживаемой наблюдателем по мере её развёртывания во времени, также как это делает партнёр/противник животного либо третьи особи, «зрители» взаимодействия»], характер среды, в которой всё это развёртывается, акторов (типология индивидов, действующих друг в отношении друга или же в «пустоту») и «оформление сцены» (отдельные компоненты среды обитания, вокруг которых развёртывается или на которые направляется действие акторов).
Поведение, если коротко, это всякое изменение организма [которое наблюдатель может выделить, охарактеризовать структурно, по специфике формы, т.е. образу, «вычерченному» движениями и действиями, реализующими данное изменение, как огонёк сигареты чертит буквы в темноте, одновременно отделив его от других форм поведения, выделенных подобным образом, и от «фона» — континуума активности, неразделённого — или неразделимого на подобные единицы. Иными словами, специфика формы означает, что данное поведение представляет собою отдельность, узнаваемую и отличимую от других равнозначных отдельностей.
Это делается исходя из «ползучего эмпиризма» натуралистов — что разделение на единицы, «схваченное» вниманием наблюдателя, естественно, т. е. данные единицы существуют в мире наблюдаемых особей, где также
1) дифференцированы друг от друга и от «фона» (возможно с другой «глубиной» разделения потока активности на «фигуры» и «фон») и, скорее,
2) всего существенны для участников взаимодействия, отслеживаемых наблюдателем, или для «зрителей», также отслеживающих происходящее там, где социальное взаимодействие действительно имеет место быть.
Дальше это предположение надо корректно проверить, что, увы, этологи забывали делать, и это сделало данное направление уязвимым для социобиологической критики. В отличие от критики этологических концепций, здесь социобиологи били в больное. От того и другого сравнительная этология пресеклась к 1990-м гг., и социобиология получила монополию в области исследований поведения2. Переводимая здесь работа Шлейдта излагает взгляд, позволяющий эту печальную ситуацию преодолеть (увы, без подробностей, только основы) — как и его статьи конца 1980-х, оставленные коллегами без внимания.
Другое «автоматическое» предположение наблюдателя, также требующее проверки — что выделенные единицы тем естественней, а форма каждой описана тем точнее, чем больше при «вычерчивании» контрастируют друг с другом и с «фоном», как как крупные льдины определённой формы в реку, забитую сплошь шугой во время ледостава. Контраст в этих случаях фиксируется интуитивно, а оценка границ между единицами и «схватывание» форм каждой из них проходят помимо сознания, почему всё перечисленное требует безусловной проверки аналитическими методами. Психологи знают, что впечатление склонно нас регулярно обманывать, и опытность наблюдателя ничего с этим сделать не может. Прим.публикатора]. Коммуникация может случаться всегда, когда поведение одного организма меняет поведение другого3 [что видно по резкому изменению образов, «сложенных» телодвижениями и действиями животного между периодом «до» и «во время» влияния, с одной стороны, и периодом «после». Они контрастируют между собой, ибо скачкообразная трансформация внешнего облика поведения в этот момент бросается в глаза на фоне его постоянства (или плавного медленными изменениями). Этот контраст «схватывается» вниманием наблюдателя, «автоматически» интерпретируясь как «отдельное влияние» демонстрируемого в тот момент (или несколько ранее) поведения особи Х на поведение особи У. Иными словами, влияние поведения одной особи на другое выглядит таким же квантованным, «распадающимся» на отдельности (единицы влияния), как его внешний образ, разворачивающийся перед наблюдателем, выглядит не непрерывным, а распадающимся на отдельности (единицы поведения, в сравнительной этологии называемые демонстрациями), каждая со своей специфической формой.
У влияний, соответственно, специфичен эффект — каждое из них меняет какое-то определённое поведение оппонента в предсказуемом направлении, также как предъявленные демонстрации с большей или меньшей вероятностью определяют выбор последующих а, значит, увеличивают предсказуемость поведения, реализованного особями на следующих «шагах» развития взаимодействия. Демонстрации это делают «со стороны» внешнего облика, влияния — со стороны его изменений, те и другие соответствуют правилу, равно верному как для онтогенеза, так и взаимодействий между особями в сообществе: «в должном месте возникает должное и действует должным образом».
Предположения о влиянии также должны быть проверены, они могут быть кажимостью в той же степени, что разделение потоков активностей участников на единицы, «схваченное» вниманием наблюдателя. Увы, влияния фиксируются чаще всего на тех же отрезках времени, на которых наблюдателем выделяются единицы, эффект первых бессознательно «сплавляется» с формой вторых, создавая новый источник ошибок в случае, если единицы поведения и их эффекты считаются непосредственно данными наблюдателю, лишь бы был достаточно опытным (а сравнительная этология держалась этого — дезориентирующего! — представления до самого своего конца). Специфика формы каждой из них начинает восприниматься по умолчанию как источник «влияний», в свою очередь рассматривающихся как «эффекты», а то и «значения» данных форм поведения. Оба таких восприятия могут быть ложными, как и родившее их неявное предположение (третье при чисто индуктивном подходе к описанию наблюдаемого поведения) может быть кажимостью ничуть не менее (на деле более) чем первые два. Поэтому оно также должно быть осознано, артикулировано, т. е. сделано явным, что позволит его проверить — как и первые два. Статья Шлейдта замечательна тем, что даёт простые и ясные методические основания для проверки всех трёх. Прим.публикатора].
Есть минимум 2 разных типа коммуникации. При фазовой (ФК) отдельный сигнал особи-передатчика вызывает постоянные изменения в особи-приёмнике. При тонической (ТК) поток сигналов первой особи модулирует поведение второй «в режиме реального времени», формируя тем самым устойчивую связь между ними.
1. Введение
Содержание
Нижеследующее — очень краткий обзор написанного [мной] о моделях поведения и коммуникации в последние 60 лет, на нескольких сотнях страниц. Я строю на фундаменте, заложенном Якобом фон Икскюлем, Оскаром Хейнротом, Конрадом Лоренцем и Нико Тинбергеном, используя новые инструменты и концепции современной физиологии и коммуникационной инженерии. Когда я рос с домашними животными и сельскохозяйственными животными, то сразу был очарован их поведением, и скоро осознал его сходство с поведением людей и младенцев.
Одна из первых моих работ была о поведении новорожденных детенышей разных млекопитающих (Prechtl and Schleidt 1950). На всём протяжении научной карьеры я предпочитал исследовать поведение легко доступных видов позвоночных: мышей, кошек, собак, кур, индеек (например, Schleidt 1948, 1961, 1998), как и людей (Schleidt 1992, Schleidt and Shalter 2003). Моим первым открытием была высокочастотная вокализация мышей и полевок, заставившая спроектировать и изготовить собственные усилители, микрофоны и громкоговорители (Schleidt 1948, 1950, Zippelius and Schleidt 1956). Поэтому мой подход к коммуникации животных был неизменно совместим с концепциями акустики и коммуникационной техники, что очень поощрялось Гансйохемом Аутрумом (1942, 1948). «Математическая теория коммуникации» Шеннона и Уивера (1949) стала мне главным источником просвещения. Мои ранние открытия в области коммуникации животных и способности на все руки мастера отметил Конрад Лоренц, и в 1951 году я стал его первым ассистентом на его исследовательской станции в Бульдерне (Вестфалия, Германия).
Первой задачей, поставленной Лоренцем, было переисследовать незавершённую работу его и Нико Тинбергена 1937 г. Они фиксировали реакцию молодняка разных видов птиц на модель «опасности сверху» — движущиеся картонные силуэты хищников и других (например, мирных) птиц. Задачей этих опытов было проверить существование у них врождённого образа «опасности» или, наоборот, «отбоя тревоги», и определить черты силуэта, определяющие то или другое (в том числе, насколько изменчив образ, всё-таки вызывающий опасность, по каким параметрам, и до каких пределов). Наиболее интригующим [в исходных опытах Лоренца с Тинбергеном] было то, что одинаковый силуэт — крест с парой равных и парой резко неравных перекладин, концы которых закруглены, для сходства с головой, хвостом и крыльями реальных птиц (рис.1)
а) вызывал крик тревоги и реакцию бегства, когда его двигали слева направо, короткой стороной вперёд, длинной назад (по сходству образа это назвали моделью «ястреба» — длинный хвост, короткая шея), но
б) однако она не вызывала какой-либо реакции, когда её двигали слева направо, длинным концом вперёд. Экспериментаторы посчитали, что она похожа на безобидную птицу — гуся и поэтому птенцам безразлична.
Рис.1. Знаменитое изображение Тинбергеном в 1951 году “манекена ястреба/гуся”, предположительно использованного Лоренцем и Тинбергеном в их опытах 1937 года.
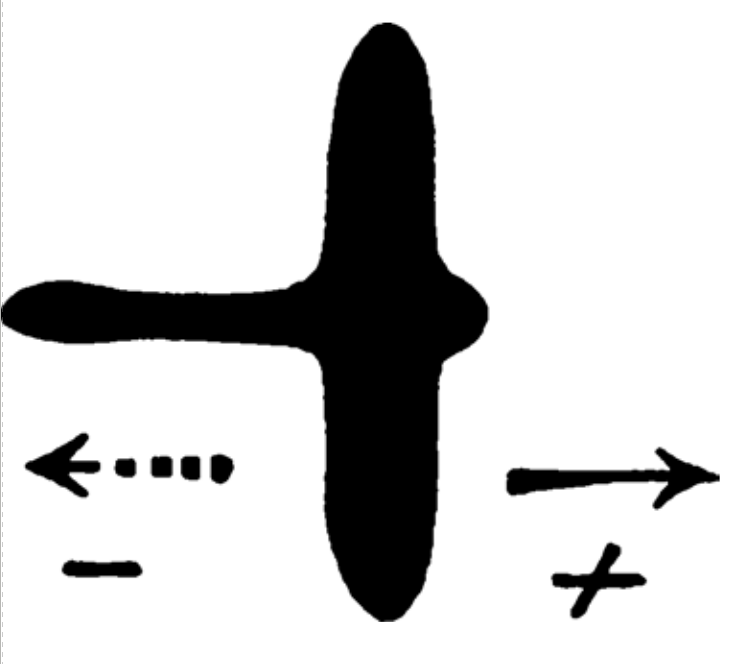
Исходные протоколы опытов были утеряны во Вторую мировую войну, и мнения Лоренца с Тинбергеном о том, что же у них получилось, различались деталями. Первый считал, что важней всего относительная скорость движения модели (пугающая летит быстро, безразличная — медленно), а эффект «гусь»/«ястреб» чётко фиксировался лишь у индеек. Второй считал, что птенцы выводковых видов — куриных, гусей и уток — реагируют бегством именно на короткую шею модели, она — релизер данной реакции (Tinbergen 1951, p. 77) [С лёгкой руки Тинбергена так и стали считать, а дальше — интерпретировали её как всеобщую, и это при том, что «гипотеза короткой шеи» на большинстве видов не подтвердилась, см.далее. Прим.перев.].
Опыты 1950-х гг. с молодняком куриных, утятами и гусятами не подтвердил «гипотезу короткой шеи». Дальше усилия сосредоточили на создании стаи индеек — единственных реагировавших бегством на силуэт «ястреба» как таковой, чтобы их было достаточно много для опытов. Вспышка смертельной протозойной инфекции (гистомоноз) заставила отложить опыты с моделями 1961 года. Воспроизведение опытов 1937 г. в расширенном варианте показала, что более сильная реакция на «ястреба» определяется предшествующим опытом птицы. Это отнюдь не врождённая реакция на релизер «короткая шея», а следствие прошлых наблюдений летящих объектов и избирательного привыкания к обычным их силуэтам (а «ястреб» необычен) (Schleidt 1961, Schleidt et al. 2011).
Десять лет ожиданий между 1951 и 1961 гг. не прошли впустую, в нашем распоряжении всегда были несколько здоровых индюков. Их демонстрации, позы с кулдыканьем стали предметом нескольких исследований (например, M. Schleidt 1954, Schleidt and Schleidt 1958, Schleidt 1964, 1974). Поскольку индейки очень социальны, имеют обширный набор вокализаций, служащих социальному общению и сплочённости группы, я приобрел значительный опыт изучения их поведения. Мое понимание их коммуникативной системы стало стандартом, с которым сравнивалось происходящее при [далее изучавшейся автором] коммуникации других птиц и коммуникации животных как таковой.
Также надо сказать, что самое первое открытие ультразвуковых сигналов в общении мышей и последующая потребность в получении минимального уровня компентентности в области акустической и коммуникационной инженерии заложили основы моего количественного метода исследований коммуникации животных (Schleidt 1973). Между прочим отмечу, что мои открытия способности различных грызунов слышать ультразвук и использовать его в своем общении стали «центром кристаллизации» для исследований на лабораторных мышах и крысах в нейробиологии, психофармакологии и психиатрии (с несколькими тысячами публикаций, например, Crawley 2007, Wöhr and Schwarting 2007). Кроме того, мое стремление больше узнать об интеллекте собак и волков (сперва в основном как хищников, нападающих на грызунов) привело к новым представлениям о коэволюции людей и волков (Schleidt and Shalter 2003).
2. Понимание сложности поведения
Что я имею в виду под «сложностью»? Это временные и пространственные связи между элементарными паттернами поведения4, которые мы можем различить [в потоке активности], отслеживаемой в наблюдениях за одним животным или несколькими [особенно взаимодействующими]. Позвольте мне дать краткий исторический обзор открытия элементарных единиц поведения как составляющих сложных форм сложного поведения, а особенно коммуникативных сигналов.
Одно из самых ранних обсуждений комплексности поведения и коммуникации находится в книге Чарльза Дарвина 1872 года «Выражение эмоций у человека и животных». Дальнейший пробел в исследованиях этой темы был прерван открытиями Хейнрота (1911) и Уитмена (1919). Затем 73 года назад, в 1936 г., произошёл прорыв, когда здесь, в Берлине, в Харнак-Хаузе в Далеме Конрад Лоренц прочитал лекцию «Концепция инстинкта прежде и теперь». Это был обновлённый вариант статьи об «arteigene Triebhandlungen», видоспецифических инстинктивных действиях, или видоспецифических формах выражения [врождённых] побуждений, в расширенном виде опубликованной под названием «К понятию инстинкта» (Lorenz 1937).
Это двигательные стереотипы, присущие каждому виду в сфере демонстраций и повседневной активности, характерные для него и поэтому узнаваемые, хорошо отличимые от (часто достаточно сходных обликом) однотипных паттернов телодвижений близких видов, почему явно различные между собой при очевидной гомологичности у данных видов (Lorenz 1932).
Лоренц поставил инстинктивные (тело)движения5 (Instinktbewegungen) в центр наблюдения и анализа этологов [в предположении их совпадения с arteigene Triebhandlungen — единицами, на которые разделён отслеживаемый поток поведения и которые характеризуются типологически определённой формой или образом, «вычерченным» соответствующими (инстинктивными) движениями, с одновременным отделением данного образа от образов других таких же единиц и от «фона». Прим.публикатора].
Единицы поведения, существенные для теории, в английском варианте получили название fixed action patterns (FAP). Роль FAP как дискретных форм или образов, «вкраплённых» в [континуальную прочую] активность животного, и [в функциональном плане] представляющих собой коммуникационные сигналы6, дальше была несколько упрощена в рамках понятий о социальных релизерах7 (Auslösers, Social Releasers) и врожденных разрешающих механизмов (ВРМ, angeborene Schemata, IRM; обсуждение концепций FAP и IRM см. Schleidt 1962, 1964, 1974, 1982).
[Правда, этологам не удалось решить и вторую важнейшую методическую задачу — насколько отдельности, реально существующие в отслеживаемой активности особей (более или менее дискретизованные, более или менее узнаваемые по специфике формы, более или менее устойчиво «вычерчиваемой» при каждом предъявлении образующими телодвижениями животного — соответствуют идеальным представлениям о «демонстрации», присутствующим в их теории? Насколько велика — или мала — степень этого соответствия для форм демонстраций у разных видов и в разных контекстах общения?
Для этого нужно разрабатывать объективный метод описания форм демонстраций, позволяющий оценить (лучше количественно или хотя бы графически) инвариантность воспроизводства каждой из форм в максимально гетерогенной выборке предъявлений (МГВ), степень дифференцированности их друг от друга и от «фона» или, что то же самое, скачкообразность смены единиц, дотоле удерживаемых стереотипно довольно подолгу.
Иными словами, нужно доказательно установить, насколько последовательность таких узнаваемых образов, «схватываемых» наблюдателем (предполагая по умолчанию, что это же делают и сами особи), не «фильм», а «слайдфильм». До работ Шлейдта этологи поступали просто — предполагали, что всякому компетентному наблюдателю, достаточно долго исследовавшему данные формы активности данного вида, искомые единицы даны непосредственно в опыте: «схваченные» его вниманием и описанные им как отдельности, они тривиально соответствуют единицам, действительно существующим в «мире» особей и существенным для них. Эта простота стала хуже воровства, т. к. немедленно вела к ошибке.
Шлейдт — и независимо от него Ноа Эшкол — её исправили, создали искомую технику объективного описания демонстраций, но в условиях концептуального кризиса сравнительной этологии их решение никого не заинтересовало. Прим.публикатора]
Серьезным недостатком поведенческих исследований в то время были трудности [точнее — ошибки] в документировании [наблюдаемого в отслеживаемой активности] и анализа наблюдений [от способа записи до собственно выделения единиц, спецификации их формы, оценки дискретности и устойчивости каждой из них, чтобы на этой основе сделать вывод, действительно ли их формы и образы инвариантны, т. е. реально присутствуют в активности особей, и равно «вычерчиваются» разными индивидами, с разным уровнем возбуждения, социальным статусом, уровнем специфической мотивации и др. собственными характеристиками, в разных частях ареала, в разные дни наблюдений и пр.? Или верно противоположное — это лишь кажимость, артефакт категоризирующего внимания наблюдателя, а поведение участников взаимодействия на деле не содержит подобных отдельностей, т. е. в данной части «развёрток» активностей представляет собой идеальный континуум? Прим.публикатора].
В лучшем случае наблюдатель писал в блокнот самые бросающиеся в глаза моменты и результаты происходящего. Действительно, один из самых уничтожающих доводов против исследований Лоренца — слова, что он не более чем коллекционер анекдотических случаев [из жизни животных] и «только рассказчик» о них.
Хотя многие из его сугубо научных работ переведены на английский, в [англоязычной] научной литературе достаточно часто цитируются лишь популярные книги Лоренца «Кольцо царя Соломона» и «Человек находит друга». Редко признавалась роль Конрада Лоренца как пионера научного кинематографа. Он начал снимать с 1935 г. и задокументировал опыты, проводимые вместе с Нико Тинбергеном в 1937 году (в конце концов опубликованы как «Поведение серого гуся», Lorenz 1950). Он также в поддержку своих сравнительных исследований [репертуаров демонстраций] фильмографировал ухаживание многих видов речных уток (Lorenz 1941, 1952, 1958, Schleidt and Oeser 2011 a, b), был одним из основателей Encyclopaedia Cinematographica.
Когда Лоренц сформулировал, что сложное видоспецифическое поведение животного составлено [также сложными] инстинктивными (тело)движениями (Instinktbewegungen), поведенческая теория всё ещё омрачалась предположением Декарта о том, что животные — это простые машины. После открытия Павловым условных рефлексов бихевиоризм Уотсона преобразовал декартовские идеи в простейшую схему «ответов на стимулы». В основе лежало предположение, что организм появляется с очень немногими врожденными рефлексами [Павлов их назвал безусловными], а вся информация, нужная для выживания, приобретается через обучение [особей, попавших в проблемную ситуацию, разрешать её, или через образование условных рефлексов, или через случайные «дёрганья» в разные стороны, с покреплением и усилением тех действий, которые приближают к решению (бихевиоризм). Прим.публикатора].
Идеи Лоренца включали два утверждения8. Первое — что сложные последовательности взаимосвязанных телодвижений, образующих закономерно развёртывающуюся структуру или форму9 могут выполняться «автоматически», т. е. без необходимости обучения и даже в отсутствие специфической стимуляции10 (оно вызвало бесконечные споры о том, что является врожденным, а что усваивается из предыдущего опыта). Второе — открытие сложной, закономерно развёртывающейся структуры поведения особей во взаимодействиях, складывающейся, как из строительных блоков, из следования друг за другом открытых Лоренцем «строительных единиц» — демонстраций, было принято как само собой разумеющееся, как факт [и не обсуждалось в сообществе, не подвергалось методическому и концептуальному скепсису. Почему этологи не смогли данное положение защитить, когда на него обрушилась волна критики — не были разработаны методы, как проверить, когда это имеет место в действительности, а когда нет. Прим.публикатора].
Поразительный технический прогресс в аудио- и видеозаписи во второй половине 20-го века, а также доступность регистраторов событий и компьютеров для хранения данных устранили нацарапанные заметки в полевых дневниках и гроссбухах с данными. Так разрешились многие проблемы, с которыми столкнулись первые этологи, кроме одной: анализа данных. Многие этологи сложили свои записные книжки и кассеты на полках и навсегда заперли дверь, потому что они не смогли найти способы проанализировать свои данные и извлечь ценную информацию. Фактически, этология все еще сталкивается с проблемой, схожей с проблемой, с которой столкнулась физика около 100 лет назад, а именно с отсутствием математических инструментов для решения рассматриваемых проблем
[точнее, с отсутствием метода, адекватного её объекту, в чём и состоял в том числе концептуальный кризис сравнительной этологии в 1970-х. До этого, при всей приблизительности и отмеченных выше огрехах, методы отцов-основателей этологии были пригодны для решения объявленных ими задач. Проблемы возникли с развитием этологии «вширь», когда исследований стало много, и по ухаживанию или угрозе одного и того же вида (или нескольких близких, предполагавших сравнение демонстраций и их эффектов для задач таксономии и микроэволюции) накапливалось по нескольку работ разных авторов, сделанных в разные годы, часто из разных стран и т. д. Единицы и/или эффекты поведения, выделенные разными авторами, оказывались несопоставимыми между собой.
Это едко описано в «Судьбах сравнительной этологии» Е.Н.Панова (2005); правда, Евгений Николаевич с водой выплёскивает и ребёнка, на этом основании заключая, что единицы поведения — это условность, нужная наблюдателю для удобства классификации, а в действительности наблюдаемая активность животного представляет собой идеальный континуум. Данная точка зрения попросту неверна; увы, защищая её, Е.Н.Панов полностью игнорирует существующие опровержения, в том числе ключевые, полученные самим Шлейдтом. Получается чисто словесная эквилибристика. Прим.публикатора].
Знаменитый математик Давид Гильберт (1862-1943) оплакивал бедных физиков, считая что их наука слишком сложна для них. Гильберт над ними сжалился, разработал новые методы математики и геометрии, значительно увеличившие их возможности (Hund, 1969).
[В науках о поведении] были прорывы, которые помогли экспериментальному дизайну и анализу данных в изучении поведения, особенно в психологии с помощью непараметрической статистики (например, Siegel 1956) или ориентации (например, Batschelet 1981). Но по сей день этологи ждут, что математик или статистик, обладающий способностями и преданностью Гилберта, разработает методы анализа поведенческой сложности [т. е. выделения форм и структур в поведении, с описанием их специфических образов, как ясно из последующего текста автора. Прим.публикатора]. Я рано осознал эти недостатки, несмотря на то (или отчасти из-за того), что мои собственные способности к математике чрезвычайно ограничены. Но даже при том, что у меня не математический ум, я хорошо разбираюсь в основах интуитивной геометрии и в практической инженерии. Это позволило мне с помощью терпеливых математиков и статистиков искать и находить методы и инструменты [решения отмеченных проблем этологии], пригодные, по крайней мере, для первого шага.
2.1. Дискретные элементы в поведении организмов
Как пример сложно организованной последовательности поведенческих актов я выбрал короткий (18 с) отрывок из первой сцены нашего фильма о половом поведении индеек (Schleidt and Schleidt 1962). Внимание наблюдателя концентрируется на отдельной индюшке, движущейся лёгким шагом вперёд в поисках [потенциального] партнёра.
![Рис.2. Пример 18 с. записи движения индюшки в поисках партнёра. Силуэты птицы — её положение и поза кадрах, соответствующих 1й, 5й, 9й и 13й секундам. Ниже схематически обрисованы поведенческие переменные: появление и смена 4-х форм поведения и 3 типов ориентации тела птицы во времени, читаемых как музыкальная партитура [в смысле закономерности и предсказуемости появления и смены в структуре паттерна] (сверху вниз: настороженность, пешая ходьба, ориентация тела в трёхмерных координатах, ориентация головы (правее-левее), высота головы (выше-ниже). 2 звука «дрр» следуют в кадрах 500 и 550 (см. Schleidt 1971, стр. 13; рисунки основаны на сцене 1 в Schleidt and Schleidt 1962).](http://www.socialcompas.com/wp-content/uploads/2022/12/Snimok-ekrana-ot-2022-12-23-21-50-22.png)
Рис.2. Пример 18 с. записи движения индюшки в поисках партнёра. Силуэты птицы — её положение и поза кадрах, соответствующих 1й, 5й, 9й и 13й секундам. Ниже схематически обрисованы поведенческие переменные: появление и смена 4-х форм поведения и 3 типов ориентации тела птицы во времени, читаемых как музыкальная партитура [в смысле закономерности и предсказуемости появления и смены в структуре паттерна] (сверху вниз: настороженность, пешая ходьба, ориентация тела в трёхмерных координатах, ориентация головы (правее-левее), высота головы (выше-ниже). 2 звука «дрр» следуют в кадрах 500 и 550 (см. Schleidt 1971, стр. 13; рисунки основаны на сцене 1 в Schleidt and Schleidt 1962).
Здесь можно выделить 4 [отдельные] формы поведения: пешее перемещение (характерной походкой), стояние, настороженная поза и контактный крик «Дрр», и 3 непрерывных переменных: ориентация тела (относительно стандартной сферы и окружностей, связанных с её осями), ориентация головы (относительно оси тела) и высота головы над землей. Ещё ряд переменных могут представлять интерес в других случаях, но в данном почти не меняются, например, «высота центра тяжести птицы над землей» и «координаты глобальной системы позиционирования» (~47°58’24’ с.ш. и 11°14’09’ в.д.) либо не могут быть зафиксированы при данном разрешении изображений, вроде изменения частоты дыхания или моргания глаз. [Зрителю] может быть интересно, что в комментариях к фильму дан подробный перечень наиболее ярких паттернов поведения, начиная с определенного номера кадра (Schleidt 1971).
Я должен отметить, что ради удобства описания этологи часто представляют паттерны поведения отдельным стоп-кадром, мгновенным изображением данного поведения, чаще всего на стадии кульминации. Их сложные образы, закономерно развёртывающиеся в [трёхмерном] времени и пространстве взаимодействия11 в этому случае сводится к простому ярлыку. Такие иконки полезны для краткого изложения [этограммы] — инвентаризации репертуара форм поведения, выделенных как отдельные у данного вида и в данном контексте общения/типе взаимодействий (например, Lorenz 1941, Schleidt et al. 1984). Однако они не должны отвлекать от того факта, что паттерны поведения мы выделяем в процессе [развёртывания отслеживаемой активности участников взаимодействия], который лучше описывается фильмом (Lorenz 1950), последовательностью кадров фильма (Лоренц 1958) или абстрактной диаграммой пространства-времени (Finley et al.198312).
2.2. Базовая геометрия в поведении организмов.
Обычно мы полагаем по умолчанию, что утка всегда остаётся сама собой, так же как роза, и можем предположить то же самое относительно кукареканья петуха
[однако в этом случае — также как для других демонстраций — это естественное предположение требует проверки аналитическими средствами, здесь недостаточно одного лишь впечатления наблюдателя о самотождественности «позы Х или вокализации У» в разных случаях независимого предъявления того и другого. Больше того, из истории сравнительной этологии точно известно, что такое решение ведёт к ошибке. Так получается именно потому, что паттерны поведения — это временные органы животного, их форма, «вычерченная» телодвижениями, может меняться плавно и незаметно для наблюдателя от одного случая «сборки» данного FAP к другому, от данного взаимодействия к другим у других особей, и т.д.
Наблюдатель, обманутый собственным категоризирующим сознанием, будет считать, что фиксирует «то же самое», и история этологии показывает, что опыт наблюдений здесь бессилен. Кажимость это или действительно отдельная форма поведения, требуется проверять каждый раз заново, и работы автора демонстрируют один из путей, каким это можно сделать.
А мои собственные работы — другой: отсюда исключительный интерес к работам Шлейдта, решившим ту же задачу за 20 лет для меня. Однако тогда решение не заинтересовало коллег, в том числе начинавших вместе с ним в кругу учеников Лоренца, чему автор очень сокрушался. Надо думать, сейчас, когда мода на социобиологию подходит к концу, показана внутренняя противоречивость её теорий, также как повсеместное несоответствие фактам, возможен возврат интереса к сравнительной этологии, ушедшего из данной области в конце 1980-х. Работы Шлейдта и, хочется верить, мои, тогда будут использованы для восстановления сравнительной этологии «на повышенном основании», с учётом новых данных, в изобилии накопленных исследованиями, выполненными в социобиологической парадигме (см.пример «расшифровки» сигналов агрессии у коростелей Crex crex), и решений, предложенных этологами в конце 1980-х и позже для выхода нашей дисциплины из концептуального кризиса, но тогда оставшихся втуне. Прим.публикатора].
Другими словами, всякий раз, когда вещь, животное, цветок или образец поведения легко идентифицировать и описать, вариативность принимается как факт [а если тяжело — наоборот, иггнорируется. Прим.публикатора]. Поэтому в первом приближении нам необходимо определить основные особенности таких вещей, как паттерны поведения. Чтобы понять сложность поведения — как оно складывается из базовых элементов — сперва необходимо определить сценарий реализации данного поведения: актора или акторов (кто с кем взаимодействует), сцену происходящего (в каком пространстве одна особь или несколько развёртывают поведение) и «реквизит» (какие события происходят друг за другом в каких частях окружающей среды, в смысле окружающей организм, рис. 3).
![Рис.3 Организмы в евклидовом пространстве [своей среды обитания]. (A): Большинство организмов представимы трехмерным телом [обладающего характерным силуэтом, с узнаваемыми изменения последнего каждым из доступных особи телодвижений (в сумме соответствующих «степеням свободы» изменения силуэта животного разными моторными актами, включая используемые при вокализации и/или мечении территории запахом]. Тело отделено от среды полупроницаемой мембраной - кожей, действующей как двусторонний фильтр. (B): Индейка в трёхмерном пространстве своей жизнедеятельности (левый рисунок основан на Schleidt and Crawley 1980).](http://www.socialcompas.com/wp-content/uploads/2022/12/Snimok-ekrana-ot-2022-12-23-21-50-39.png)
Рис.3. Организмы в евклидовом пространстве [своей среды обитания]. (A): Большинство организмов представимы трехмерным телом [обладающего характерным силуэтом, с узнаваемыми изменения последнего каждым из доступных особи телодвижений (в сумме соответствующих «степеням свободы» изменения силуэта животного разными моторными актами, включая используемые при вокализации и/или мечении территории запахом]. Тело отделено от среды полупроницаемой мембраной — кожей, действующей как двусторонний фильтр. (B): Индейка в трёхмерном пространстве своей жизнедеятельности (левый рисунок основан на Schleidt and Crawley 1980).
Поведение, как правило, представляет собой любое изменение организма и может быть описано одним или несколькими из следующих действий: изменение местоположения, ориентации, геометрии поверхности, поверхностное излучение и / или поверхностное поглощение вещества, энергии и/или информации (Рис.4).
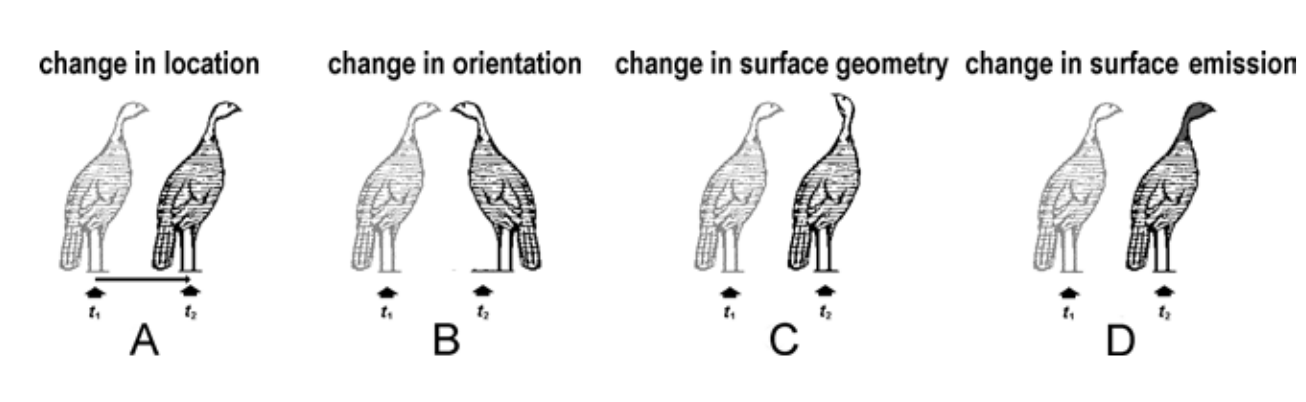
Рис.4. Индюшка в своем евклидовом пространстве, демонстрирующая четыре основные категории поведения как изменение между двумя моментами: t1 и t2: изменение местоположения (A), ориентации (B), геометрии поверхности (C) и излучения поверхности и / или абсорбция (D).
Возвращаясь к моему предыдущему утверждению, нам нужно определить
a) сценарий, в котором разворачивается данное поведение,
б) его субъект (-ы) (автор использует социологический термин «актор (-ы)») — конкретный индивид (-ы), действующие в определенной обитания (в т.ч. действие разворачивается в определённом пространстве-времени, обычно присущем поведению, а иногда и сугубо специфическом для него),
в) реквизит (значимые компоненты окружения или моменты сценария происходящего).
Что такое реквизит, я проиллюстрирую на примере значимых составляющих окружающей среды самки индейки (рис. 5).
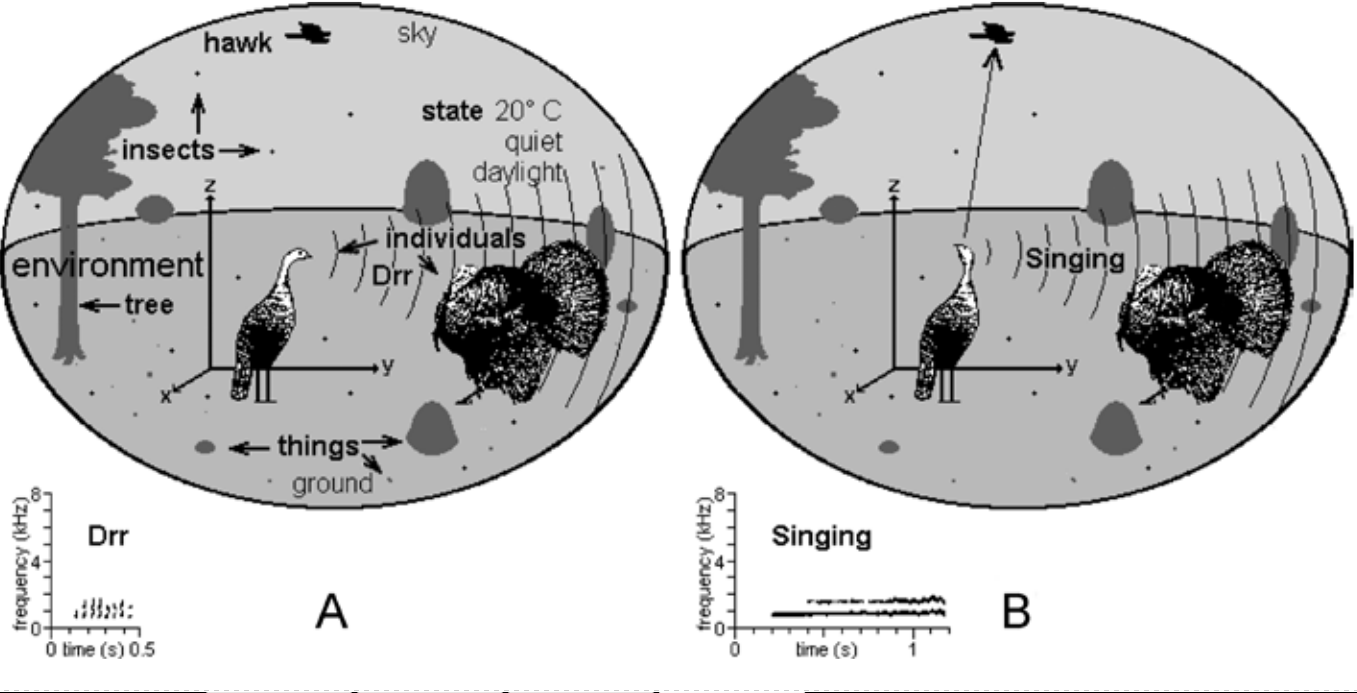
Рис.5. Индейкино окружение включает потенциальных партнеров (индюков), потенциальных хищников (ястреба), потенциальную пищу — насекомых, и вещи вокруг, могущие использоваться в разные моменты её жизни (реквизит): деревья, на которых ночуют и отдыхают, предметы на земле (обследуемые и переворашиваемые при кормодобывании) и пр. Присутствуют и дополнительные переменные, например температура, уровень шума вокруг, освещённость днём и пр., могущие влиять на поведение как её, так и её компаньонов.
В сцене слева (рис.5A) индейка подпускает к себе (сближающегося с ней) индюка, издавая контактный крик «Drr»; справа (рис.5B) она заметила над своей головой ястреба, и издала тревожный сигнал («пение»). Вокализации распространяются во всех направлениях, но на рисунках показано, что они адресованы сородичу, чтобы не загромождать их слишком большим количеством линий.
Должное внимание к окружающей среде [животного, реализующего поведение] в той форме, что описана выше, впервые было предложено Якобом фон Юкскюлем (1909) и незабываемо проиллюстрирована в его «Прогулке по образам мира [умвельтам] человека и животных. Иллюстрированная книга невидимых миров» ((von Uexküll and Kriszat 1934, von Uexküll 1957) ). Приняв во внимание предыдущие соображения, мы получим доступную концептуальную основу для количественного анализа моделей поведения, свободную от бесплодных споров о том, что не содержится в данных и соответственно, непроверяемо: в первую очередь, особенно является ли конкретный паттерн поведения врожденным или выученным, однако не закрывает возможность дальнейшего исследования этого вопроса, и точного определения соотношения генетических факторов и процессов обучения, сформировавших данный паттерн [поскольку не предрешает его, выбрав заранее одну из возможностей, и настаивая на ней. Далее автор показывает, что также надо поступать и в отношении подразделённости наблюдаемого поведения на отдельности со специфической формой или, напротив, континуальности происходящего — это должно быть выводом из анализа данных, а не исходной посылкой, как в концепции Е.Н.Панова, априори настаивающего на «идеальном континууме» и сугубой условности всех выделяемых единиц, а отчасти и у его оппонентов, классических этологов, слишком доверившихся непосредственному восприятию и забывших, что его сугубая категориальность отнюдь не обязательна для животных, развёртывающих отслеживаемую им активность (как мы убедимся далее). Прим.публикатора]
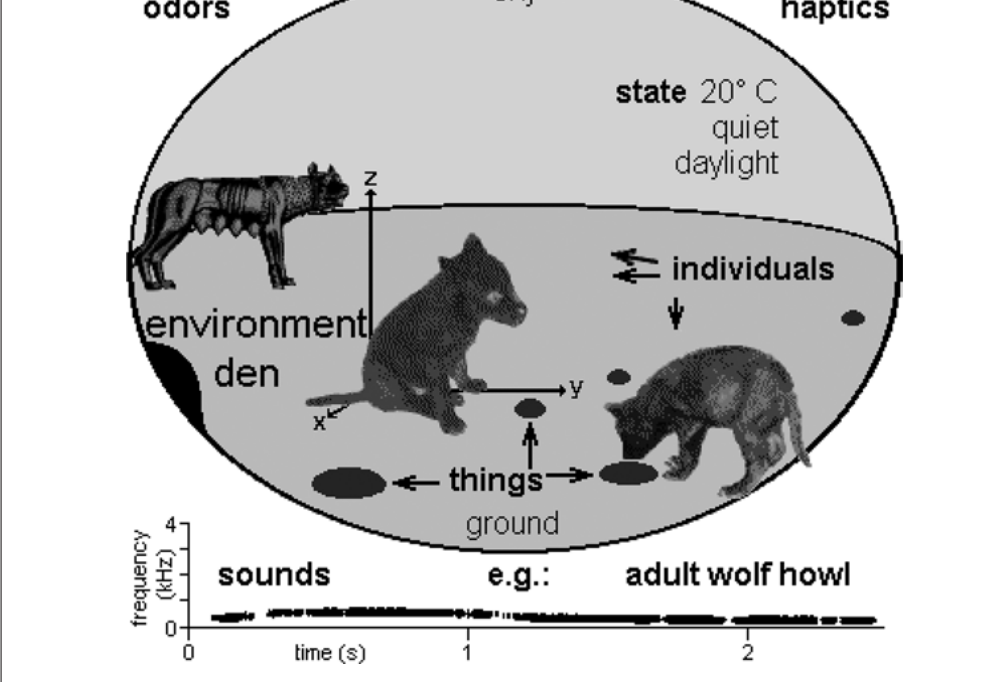
Точно также и не приобретенным, без исключения будущего анализа генетики и обучения. процессы, которые его сформировали. Кроме того, мы должны осознавать свою предвзятость как наблюдателей: она нас заставила предпочесть наблюдать, регистрировать и анализировать визуальное и акустическое поведение животных сильно больше, чем проиводимое им в других сенсорных модальностях (обоняние, вкус, осязание) [сюда же относится категориальность восприятия, следующая из использования языка]. У детёнышей многих млекопитающих глаза и уши после рождения закрыты; они полагаются на запах и тактильную информацию, чтобы найти своей матери, задолго до того, как увидят свет (рис. 6).
[Автором в 1980-е гг. были разработаны способы] тщательной регистрации поведенческих последовательностей, позволяющие преобразовать их в числовые данные. Благодаря им стандартные алгоритмы обнаружения, классификации и распознавания образов становятся полезными инструментами для оперирования как с поведенческой сложностью [выделение единиц, их описание и классификация, от дифференциации друг от друга и от «фона» до установления соподчинённости единиц разного уровня в потоке активности, какая какие включает, как матрёшка], так и с его типическим окружением (Schleidt 1985).]
Рис.6. Умвельт волчонка. Выходя из логова, волчата впервые воспринимают мир света, к их прежнему миру тактильных ощущений, запахов, звуков и вкусов добавляются видимые образы.
3. Коммуникация между животными
Прежде обсуждения роли дискретных элементов поведения как коммуникативных сигналов [в отличие от нас, автор «элементами» везде называет единицы], нужно не только прояснить, что именно мы подразумеваем под коммуникацией в рамках общего представления о поведении организмов, но также уделить особое внимание кластеру близких друг к другу [и важных для понимания «коммуникации»] концептов: «сигналы», «сообщения» и «информация». Все три иногда используются взаимозаменяемо, что приводит к значительной путанице: поскольку сенсорная система наблюдаемого вида [как минимум птиц и млекопитающих] достаточно сходна с таковой наблюдателя, знакомство последнего с видоспецифическими паттернами поведения [отмеченными как отдельности при собственных предыдущих просмотрах взаимодействий или, что сильно хуже, выделенных другими авторами] достаточно для обнаружения наиболее заметных сигналов.
Однако конкретное содержание сообщений, кодируемых сигналами, во многих случаях остаётся загадочным, как и что именно передаётся. Это сравнимо с отслеживанием пути заказного письма от почтового отделения, куда его отнесли, до передачи почтальоном непосредственно получателю в руки, когда тот своей подписью удостоверяет надёжность [и/или безопасность — английское safe означает то и другое, что крайне полезно именно в описании коммуникации, где обе проблемы неразрывно связаны между собой, ввиду позитивной корреляции эффективности и риска используемых сигнальных сообщений, и решаются участниками одновременно] доставки. Мы знаем, что общение состоялось, но нет подсказок, о чём общаются участники или что было передано в сообщениях.
Приняв во внимание эти соображения, наблюдатель может зафиксировать что в разворачивающемся перед ним взаимодействии происходит коммуникация — обмен сообщениями и передача информации между участниками, без знания, о чём конкретно общаются и что передаётся в отдельных сообщениях [благодаря чему он избавлен от априорных предположений по этому поводу, которые неизменно оказываются контрпродуктивными, т. к. заставляют исследователя подбирать поддерживающие их наблюдения и исключать не ложащиеся в эту картину; лучший пример — работы Е.Н.Панова, выступившего с такой априорной концепцией ещё в 1970-х и до сих пор активно и агрессивно отторгающего любые не согласующиеся с ней данные, проявляя при этом чудеса умственной изворотливости и одностороннего взгляда, ибо «не согласующееся» появляется на каждом шагу. Однако он об этом молчок. Прим.публикатора]. На деле, коммуникация и информационный обмен диагностируются с помощью относительно простой статистической парадигмы:
«Коммуникация между двумя животными происходит всякий раз, когда вероятность определенного поведения принимающего коммуниканта с положительной задержкой со временем связана с выходным сигналом передающего13».
3.1. Статистический анализ коммуникативных сигналов
«Песни» птиц, также как насекомых или лягушек, из-за их красоты вместе с видоспецифической природой, с античности были предметом спекуляций относительно их функции.
С появлением звукозаписывающего оборудования, особенно чувствительных микрофонов, магнитофонов и колонок во второй половине ХХ века, анализ вокализаций животных сильно продвинулся вперед, их документирование, статистический анализ и эксперимент с ними стали высоко изощрёнными. Из-за доступности биоакустических методов анализа именно звуковые сигналы стали предпочтительным предметом исследований в области коммуникации животных. Из всех переменных, требующих учёта в любом статистическом анализе, важнее всего частота появления сигнала. Скажем, контактный крик вроде «Drr» индейки (рис.7), издаваемый в некоторых ситуациях каждые несколько секунд в текущем диалоге членов стаи, в течение одного дня может вести к сотням событий и дать много данных для анализа.
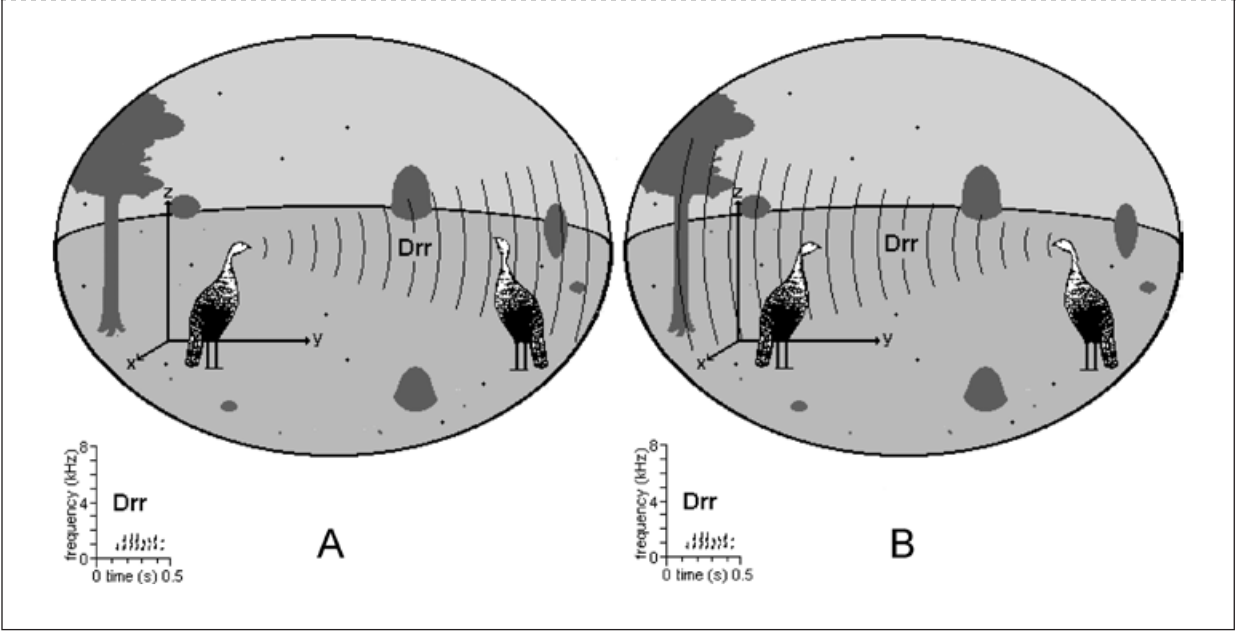
Рис.7. Индюшка в своём окружении, включая другую индейку той же стаи. В сцене слева (A) она идёт, издавая контактный крик «Drr», справа (B) индейка отвечает тем же «Drr». Как отмечалось в подписи к рис. 5, звук распространяется во всех направлениях, но на рисунках показана адресация конкретному сородичу, чтобы не загромождать их.
Другие коммуникативные сигналы, вроде серий сигналов тревоги с воздуха («пение», рис. 5Б), могут быть вызваны высоколетящим хищником только раз в несколько дней, даже недель. Таким образом, часто встречающиеся сигналы будут предпочтительнее редких. Эта предвзятость в поиске сигналов, подходящих для количественных исследований, важна не только в плане удобства (сокращения времени на сбор данных), короткий промежуток времени, за которое собраны пробы, обеспечивает основное требование статистического анализа — их однородность. Чтобы её обеспечить, система, из которой собраны данные [т. е. взаимодействие, отслеживаемое наблюдателем, непрерывно регистрирующим потоки активности участков, или фиксирующим их поведение, производимое через некоторые небольшие промежутки времени, скажем каждый несколько с] должна быть в «устойчивом состоянии», устойчивом по отношению к исследуемому сигналу [обстоятельства его издавания, внешние и внутренние, не должны качественно меняться, хотя могут развиваться преемственно результатами предшествующих событий взаимодействия. Или, иначе, особи реализуют одно и то же поведение, преемственно продолжающее начавшееся взаимодействие, и не «перепрыгивают» к качественно иной активности, хотя могут «перебивать» прежнее поведение паузами. Прим.публикатора].
Один из аспектов вышеизложенного алгоритма поиска и доказательства коммуникации во взаимодействии, между её потенциальными участниками, является «положительный лаг» [между паттерном поведения одной особи, вероятно с сигнальными свойствами, и характерным ответом на него, предположительно выбранным на основе сигнальной информации — именно по причине его характерности и специфической связи с данным паттерном поведения второй особи, более сильной и более устойчивой, чем аналогичная связь с возможными предыдущими формами поведения самой этой птицы.
Как верно заметил Е.Н.Панов, всякое обоснование коммуникации, в том числе исходящее из определения, представленного В.Шлейдтом (впервые его сформулировал Александером в 1960 г.) требует показать, что в соответствующих взаимодействиях «перекрёстные» влияния и зависимости (форм поведения одного участника от одновременных с ним форм поведения другого) у каждого из них бывают сильней «продольных» (зависимость текущего поведения демонстратора от его же прошлого). Причём показать на максимально гетерогенной выборке предъявлений, убедившись тем самым, что исследуемые влияния устойчивы во всём спектре варьирования внутренних (мотивация/уровень возбуждения демонстратора) и внешних (стимуляция данного оппонента, риск нападения хищников, привлекаемых демонстрациями обоих, разные место и время наблюдений, сообщества, в которых наблюдают, или же социальные статусы участников) обстоятельств предъявления демонстрации Х.
Те и другие могут дестабилизировать отыскиваемую нами связь между формой сигнала Х и ответом Х´, нарушить её. Также возможна ошибка противоположного рода: независимая обусловленность Х и Х´ одинаковыми обстоятельствами контекста может имитировать эту связь в ряде частных ситуаций (создав ложное представление, что первое сигнал, а виторое — ответ), но она «рассыпается» при рассмотрении полного спектра обстоятельств предъявления того и другого. Прим.публикатора].
«Положительный лаг» означает, что изменение поведения реципиента потенциальных сигналов начнётся через некоторое время после его испускания особью-передатчиком. Задержка определяется временем прохождения сигнала (что критично в случае звуков) и реактивностью принимающей особи (чем она выше, тем ниже время задержки между поступлением сигнала и ответной реакцией).
Нетривиальность таких рассуждений видна на примере индюков, общающихся рано на рассвете посредством кулдыканья с ночёвочных насестов, расположенных в 2 км друг от друга. Сигналы с «насеста один» распространяются со скоростью около 343 м/с, т. е. к насесту 2 звук придёт с задержкой около 6 с. Стандартный ответ индюков на него — закулдыкать самому. Время ответа зависит от громкости приходящего сигнала и в зависимости от реактивности меняется 200 мс (когда она максимальна) до более 1 с (она наименьшая). На дистанции в 2 км громкость кулдыканья с насеста 1 будет всего на несколько дБ выше средовых шумов вокруг второго насеста, что добавит еще секунду к времени распространения сигнала. Т.е. в этой системе из двух индюков, испускающих слышимые обоими крики так, что возникнет предположение об общении14, общий положительный лаг между ними должен быть не менее 7 с. Если индюк со второго насеста всё время кулдыкает с задержкой от 7 до 9 с после первого, можно предположить, что он это делает именно в ответ на данный сигнал [а не, скажем, от собственного возбуждения и общей радости жизни]. В этом случае коммуникация имеет место быть.
Однако обычно несколько птиц, индеек и индюков, делят общий насест. Поэтому время ответа одного из них на кулдыканье другого составляет лишь долю секунды, каждый такой звук вызовет настоящий взрыв кулдыканья всех индюков на данном насесте. За этим коллективным “мегакулдыканьем монстров” следует рефрактерный период в несколько с: замолкнув, индюки напряжено вслушиваются, ответят ли им с насеста 2. Таким образом, регулярные раунды обмена вокализацией между соседними насестами создают истинный диалог между индюшачьими стаями с временным интервалом около 15 с между кулдыканьями.
Основная функция кулдыканья рано утром у индюков, как и песен большинства певчих птиц — сигнализация соседям о присутствии и регулярной активности данного самца [интенсивность пения и устойчивость его временного ритма к песням других особей часто связаны с «качеством» певца, как и отдельные особенности акустической формы вокализаций, составляющих его песни]. Эти функциональные аспекты рассматриваются в следующей главе.
Другой важный аспект коммуникации — «отсутствие ответа» часто есть также ответ, показатель того, что коммуникация имела место, при наличии достаточного объёма выборки данных, чтобы посмотреть изменения вероятности ответа во времени [с момента испускания возможного сигнала]. Скажем, индюк, акустически изолированный от всех прочих в камере со стенками, поглощающими звук и не дающими эха, кулдыкает самопроизвольно с определённой периодичностью. Если ему проиграть запись кулдыканья, он немедленно закулдыкает сам, после чего следует долгий период тишины (намного дольший, чем промежуток между спонтанными кулдыканьями, бывшими буквально минуту назад). Таким образом, индюк, слышащий чужое кулдыканье, не просто специфически возбуждается им [почему мог бы кулдыкать столь же спонтанно, только в более интенсивном режиме], но также «переключается» от спонтанной активности к коммуникации.
Издав ответное кулдыканье, он тормозит выдачу следующих на время, большее промежутков в спонтанной активности, и вслушивается, не придёт ли следующий сигнал, отвечая уже на него. Сходный принцип работает и в перекличках петухов: крик другого на мгновение кукареканье самой птицы, но увеличивает вероятность ответного кукареку через несколько секунд. Статистические аспекты взятия выборки и анализа сигналов в этом и других случаях довольно подробно обсуждены Шлейдтом (1973).
3.2. Два типа коммуникации: фазовая и тоническая
Рассматривая широкий спектр случаев, в которых можно показать, что коммуникация имеет место, мы сталкиваемся с поразительной разницей между сигналами, появляющимися достаточно редко, и испускаемыми часто. Пример первых — крики тревоги, вызываемые только опасностью определённого рода, скажем, с воздуха, но не с земли [хотя тут есть нюансы: если разные крики не «именуют» определённых хищников, отличая одного от другого, а лишь выражают разный уровень страха при их появлении, этот последний — а, следовательно, и сам сигнал, может быть вызван искусственно разными прочими стимулами. Прим.публикатора]. В природе они оказывают поразительное воздействие на принимающих особей: открытое выражение паники, так что для доказательства того, что коммуникация имела место, не нужно значительной выборки ответов. Таков случай «пения» — сигнала воздушной тревоги индюков, издаваемого в ответ на летящего высоко в небе хищника (рис.5Б).
Я предложил называть этот тип коммуникации фазическим: он «переводит» особь-приёмника сигнала в другую фазу (в другой режим активности, способ действия), в определённую сторону меняет его поведение или внутреннее состояние, что продолжается некоторое время (обычно характерное для данного сигнала: несколько минут, часов, остаток дня или даже всю жизнь).
Второй тип общения, тонический, включают контактные крики социальных животных, сообщающие сородичам «Вот я — где ты?» (как это использовал К.Лоренц в названии для его Ethologie der Graugans («Этология серого гуся»; Lorenz 1988, 1991). Очень частое издавание контактных криков упрощает сбор большой выборки регистраций потенциальных сигналов и возможных ответов на них для исследований изменений вероятности последних как индикатора наличия коммуникативной связи между данными птицами, как в случае сигнала «Drr» у индеек (рис. 5A, 7). Я предложил назвать эту коммуникацию тонической, т. к. сигналы здесь действуют как поток нервных импульсов, поддерживающих тонус мышцы: их постоянное повторение поддерживает контакт между коммуникантами и может модулировать поведение обоих [обычно плавно и постепенно].
3.2.1. Фазическая коммуникация
Один из её самых впечатляющих случаев — переключение индюшек с насиживания яиц на вождение цыплят через прослушивание их повторяющихся контактных криков. Птенцы начинают издавать их ещё в яйце, и продолжают при вылуплении и дальше — в следовании за самкой. Не слышащая их индейка убивает появившихся индюшат как только увидит: принимает за хищников и сильно клюёт между глазами (Schleidt et al. др. 1960). Эту странность узнали случайно, когда глухая индейка немедленно убивала всех вылупившихся цыплят.
В последующих экспериментах насиживающим индюшкам с нормальным слухом предъявляли пушистые модели цыплят с динамиками внутри, могущие воспроизводить контактные крики. Увидев молчащие модели цыплят, наседка немедленно среагировала поведением защиты гнезда, взъерошила перья, шипела, клевала модели. Сразу после включения записи контактных криков поведение слышавшей их индейки сменилось: исчезла агрессия, закудахтав, она заглянула под себя, как будто знала, что звук должен идти от яиц (хотя она насиживала не настоящие яйца, а гипсовые модели).
Наблюдения за насиживающей индейкой при вылуплении птенцов показали, что когда из-под неё появляются первые обсохшие и пушистые индюшата, наседка сильно волнуется и даже клюёт их — но только пока те молчат. Как только цыпленок издал контактный крик или даже пищал от боли, мать закудахтала и успокоилась. Всякая враждебность у неё исчезла примерно за час, завершив переход от насиживания к вождению выводка. Столь же короткий коммуникативный процесс происходит во время «импринтинга» своих родителей у птенцов выводковых птиц, покидающих гнездо вскоре после вылупления (Lorenz 1935, Hess 1973).
Вопреки распространенному мнению, импринтинг не является ни мгновенным событием, ни постоянным и неизменным по результатам. Однако центральный момент, без которого он не случается — довольно короткий обмен сигналами, имеющий длительный фазовый эффект.
3.2.2. Тоническая коммуникация
Как уже упоминалось, пример тонической коммуникации — контактные крики социальных животных, сообщающие: «Вот я — где ты?». Они передают информацию об идентичности и текущем положении особи в пределах досягаемости для её компаньонов, могущих это слышать (т. е. её социальном «существовании» для них). Эти сигналы испускаются очень часто, поддерживая связь между коммуникантами, позволяя им действовать скоординированно, создавая между ними союз, превращающий участников общения [из независимых индивидов] в социальную единицу — семью, стадо и т. д. Их частое повторение облегчает фиксацию большого объёма данных для статистических исследований, скажем, чтобы «схватить» изменения вероятности ответа как индикатора коммуникации, имеющей место быть.
У многих видов птиц песня функционирует также как контактный крик, равно как сходные с песней вокализации самцов куриных (кукареканье петуха, кулдыканье индюка, «подь-полоть» европейского перепела). При тонической коммуникации поток сигналов кодирует характер передаваемой информации независимо от расстояния между коммуникантами, что ведёт к форме кодирования, которая в технике связи называется импульсно-кодовой модуляцией.
Существует минимум две разных формы коммуникации: фазическая (ФК, один сигнал особи-передатчика уже меняет [направленно и предсказуемо] поведение особи-приёмника) и тоническая (ТК; поток сигналов особи-передатчика непрерывно модулирует поведение приёмника и формирует социальное объединение из обоих). ТК найдена у многих видов: везде, где насыщенность сигналами канала связи высока, она высоко вероятна.
Функции песни — привлечение потенциальных партнеров и «отталкивание» других самцов — были известны уже некоторое время, но сложность системы долго недооценивалась. Оба эффекта интерпретировались как своего рода безусловный рефлекс, заставляющий самок приближаться к поющим самцам и удерживающий на расстоянии соседних самцов как эффект равновесий притяжения и отталкивания. Думаю, что сложность проблемы впервые проявилась вместе с возможностью качественной записи песен на магнитофон и воспроизведения их птицам. Оказалось, что (например у птиц-печников) каждый самец знает песни соседей, а в перерывах между собственным пение слушает чужое (как вышеупомянутые индюки). Если соседская песня звучит с другой стороны, это интерпретируется как нарушение «status quo», немедленно расследующееся [владельцем] на месте (Weeden and Falls 1959, Falls 1982, 1992).
4. Заключительные замечания
Во введении я указал на важность новых инструментов и концепций современной физиологии и инженерии коммуникации для нашего стремления расширить наше понимание поведенческой сложности в коммуникативных процессах. Биология больше не является изолированной дисциплиной, а, скорее, стала неотъемлемой частью естественных наук.
Рис. 8. Взгляд на аналитический инструментарий В. М. Шлейдта.
Когда речь идет о явном поведении, прочная основа в физике имеет первостепенное значение, равно как и химия и физиология при рассмотрении внутренних механизмов, например, мотивации. А несколько конкретных алгоритмов для исследования поведения и общения — инструкции по использованию и оттачиванию инструментов, которые я считаю наиболее полезными — перечислены в справочных материалах. На рисунке 8 я предлагаю заглянуть в свой набор аналитических инструментов. Благодарности Я благодарю Майка Шалтера и Дороти Грейси за их критическое прочтение этой статьи.
«Куда идёшь, поведенческая экология?» Прошлое, настоящее и будущее изменяющейся науки. International Symposium of the Humboldt-Universität zu Berlin and the German National Academy of Sciences Leopoldina Berlin, April 30 to May 4, 2009. // Nova Acta Leopoldina. Bd.111. №380. Halle (Saale), 2013. P.61-76.
Примечания
1Посвящается памяти моих учителей, особенно Конрада Лоренца, Ганса Боршайна, Ганса-Йохема Аутрума, Якоба фон Юкскюля и Оскара Хейнрота. Прим.автора.
2См. «Про силу научной моды», 1-2; «Взлёт и падение сравнительной этологии». Прим.публикатора.
3Курсив везде автора, за исключением мест, где специально указано иное.
4чтобы дальше использовать их как отдельности в нижеследующем анализе, скажем отграничивая их формы и устанавливая функцию в рамках описания поведения данного вида (создание этограммы). Анализ временных (что за чем следует) и пространственных связей между паттернами (в какой части участка что предъявляется, скажем, захватчику или потенциальному брачному партнёру) имеет смысл только если они 1) реально присутствуют в потоке активности как отдельности, 2) действительно дифференцированы друг от друга и от «фона», не просто кажимость, возникшая из-за особенностей восприятия наблюдателя или условность, удобная для описания, требующего как-то подразделять происходящее на ограниченное число операбельных категорий.
Для автора «паттерн» это «модель, шаблон»: везде, говоря о паттернах, автор определяет паттерны по специфике формы — распознаёт их среди прочей активности, отграничивает от неё или от других паттернов, следующих перед или сразу за данным, пусть даже отчасти или полностью составленных сходными моторными актами. Иными словами, каждый из них рассматривается им как потенциальная единица поведения, демонстрация; все вместе они образуют перечень — этограмму. Прим.публикатора
5Такое название точней, чем обычное «инстинктивное действие» или «-акт», ибо подчёркивает главное в инстинкте — закономерное и последовательное (поэтому предсказуемое) «вычерчивание» его характерной формы или образа, так что последняя оказывается инвариантной в максимально гетерогенной выборке предъявлений, в том числе остаётся устойчивой и узнаваемой (сохраняет ключевые характеристики, определяющие её как таковую и отличающие от сходных форм близких видов) несмотря на значительные межиндивидуальные различия в исполнении данной демонстрации, как в известной работе Pientz, 1985, посвящённой долгому крику двух южнополярных поморников, видов-двойников.
Инстинктивные (тело)движения направлены именно на устойчивость воспроизведения своей специфической формы, вопреки более или менее обильным «помехам», равно происходящим от возбуждения «внутри» демонстратора или противодействия оппонента вовне. Поэтому форма инстинктивных (тело)движений и их сериальных единиц — демонстраций — более или менее инвариантна в максимально гетерогенной выборке предъявлений (МГВ). Она включает данные по разным особям, или взаимодействия с разной интенсивностью, на разной дистанции, или демонстрации данных поз либо вокализаций с разной экспрессией, при нахождении адресантов под разными углами друг относительно друга и пр. Форму (хараткерный образ) демонстраций можно считать инвариантом, если он сохраняется, не «расплываясь», с ростом гетерогенности выборки (а иногда и делается более чётким). В этом случае форма демонстраций обладает типологической определённостью, сравнимой с таковой «обычных» морфоструктур, и может использоваться в систематике и эволюционной морфологии наравне с ними. С той только особенностью, что демонстрации — это «временный орган» животного, комбинация образующих телодвижений «собирает» их в определённые («должные») моменты взаимодействия, «выписывает» их сопряжённой работой специфический образ «должной» демонстрации, удерживает его более или менее долго (скажем, пока не подействует), чтобы затем сменить следующей столь же временной комбинацией. Другие типы движений — прямые действия и экспрессивные реакции, наравне с инстинктивными присутствующие в отслеживаемой активности, предъявляются совершенно иначе.
Прямые действия агрессии, бегства, спаривания и других побуждений животного выражают эти последние напрямую (а образующие телодвижения демонстрации — превращённо, в ритуализованной форме). Их реализация управляется целью действия, их движения гибки и пластичны, ибо меняются и перестраиваются при исполнении, чтобы её достичь (клюнуть противника, остановить самку и сделать садку и пр.) вопреки возникшим препятствиям. Поэтому их движения вообще не воспроизводят какой-либо определённой формы или образа, которой можно было бы охарактеризовать действие в целом, оно характеризуется только целью.
Экспрессивные реакции обусловлены общим неспецифическим возбуждением животного, и не имеют какой-либо цели действия. Поэтому они никуда не направлены: среди составляющих их движений примерно поровну движущих животноеназад и вперёд; к объекту, вызвавшему возбуждение, или прочь от него. Нет у них также и специфической формы, исполнение этих движений изменчиво и неустойчиво, как пламя свечи, хотя максимально демонстративно и ярко, почему они первыми бросаются в глаза наблюдателю, как движения крыльями больших пёстрых дятлов, ложно считавшиеся «угрозой слётку». При «вычурности» и «неестественности» исполнения, сравнимой с ритуализованными демонстрациями, у экспрессивных реакций не получается выделить инвариант образа, репрезентируемый партнёру и существующий как реальность для всех участников и «зрителей» взаимодействия (в меру устойчивости в МГВ). Для демонстраций его наличие обязательно.
Иными словами, телодвижения и действия в составе демонстраций отличаются типологически определённой формой, узнаваемой и характерной для каждой из них, прямые действия — специфической целью активности и определённым направлением движений из их состава. Экспрессивные реакции не имеют ни управляющей ими цели, ни устойчивой формы, но сравнимы с демонстрациями по «яркости» и «неестественности» исполнения. Прим.публикатора
6где «значение» или «эффект» закодирован набором характеристик, определяющих самоё форму, почему одно предъявление последней действует на животное-реципиент сильней непосредственной стимуляции. Большие синицы, услышавшие сигнал тревоги, возобновляют кормёжку позже, чем увидевшие хищника непосредственно. В полевом эксперименте использовали пары синиц, ранее незнакомых друг с другом: одна из них видела хищника, а другая реагировала на тревожные крики первой (Lind et al., 2005).
Пушистопёрые дятлы Dendrocopos pubescens зимой часто кочуют со стаей синиц. Дятлы перемещаются поодиночке, постоянной парой или группой из нескольких одиночек, кормящихся вместе, но совершенно независимых друг от друга. Заметив потенциальную опасность, а иногда и просто насторожившись, кормящийся дятел подает время от времени тревожный сигнал. Сигнал может информировать птиц о потенциальной опасности, а может выражать общее беспокойство или возбуждение особи, не связанное с конкретной опасностью и запуском оборонительного поведения.
Показатель уровня настороженности кормящейся птицы – частота осматриваний, интенсивности кормления – частота нахождения корма (скорость проглатывания добычи в единицу времени). Распределение временных интервалов между осматриваниями отличается от предполагаемого экспоненциального: длительность интервала обратно пропорциональна времени, прошедшему с момента последнего осматривания (это так называемая марковская цепь первого порядка). Дятлы чаще осматриваются в присутствии потенциальной опасности – это может быть человек, собака, или макет ястреба, поставленный исследователем. Осматривания урежаются при увеличении числа особей в стае (Sullivan, 1985a, b).
Следовательно, уровень оборонительной мотивации особи не является постоянной величиной, но циклически меняется под действием эндогенных факторов. Кроме того, частота осматриваний повышается, а интервал между осматриваниями укорачивается при наступлении «тревожных событий» вокруг особи, вроде обнаружения дятлом хищников, поступления тревожных криков других дятлов или контактных криков синиц как сигналов отбоя тревоги (Sullivan, 1984). В первом случае дятел и окружающие его синицы реагируют непосредственно на самоё опасность, во втором реагирование происходит опосредованно, на основании сигналов (которым животное должно «доверяться», что имеет свой выигрыш и свою «цену»).
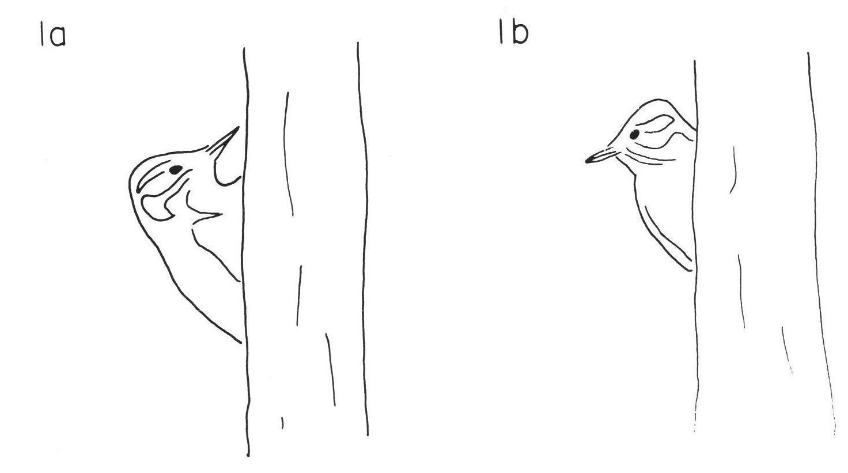
a. Пушистопёрый дятел смотрит вперёд. b. Он же осматривается, поворачивая голову направо
Последующий экспериментальный анализ показывает, что у D.pubescens система сигналов тревоги функционируют в качестве специализированного «языка», «указывающего» на опасность с воздуха. Кормящимся дятлам предъявляли макет ястреба и фиксировали их реакцию на появление настоящих ястребов. Дятлы не употребляли криков тревоги, находясь в одиночестве, в стае, не включающей самцов и самок, в стае из дятлов одного пола. Когда рядом находятся самцы и самки, они часто издают крики тревоги, причём с равной частотой у обоих полов (Sullivan, 1985b).
На тревожные крики (свои и синиц) дятлы реагировали точно так же, как на реальную опасность (ястреб) или потенциальную угрозу (наблюдатель, модель ястреба). Они замирали на несколько секунд, потом осматривались и возобновляли кормление через несколько минут. При кормлении в периоды проигрывания контактных криков синиц (сигнал «отбоя») одиночные дятлы сохраняют меньший уровень настороженности и достигают большей скорости кормления, чем в периоды отсутствия контактных криков или при проигрывании дятлам криков юнко Junco hyemalis, воробьиной овсянки Spizella arborea или соснового чижа Spinus pinus, — обычных птиц, никогда не входящих в синичьи стаи (Sullivan, 1985a-b).
Это означает «доверие» к сигнальной информации в жёстких зимних условиях, когда «нечестная» или «неточная» коммуникация стоят птице жизни – либо из-за нападения хищника, либо из-за сниженной эффективности кормления из-за вынужденной постоянно высокой бдительности («доверие» к «честным сигналам» позволяет её существенно снизить). Такое «доверие» к сигналам «оправдывается», интенсивность осматриваний становится пропорциональной риску, причём для особи полезно не только наличие, но и отсутствие сигналов. Первое информирует об опасности, второе экономит усилия особи, уменьшая уровень настороженности в «безопасные» периоды, сочетание того и другого увеличивает эффективность кормления, особенно в долговременной перспективе. Язык здесь понимается в смысле де Сооюра, как специализированная знаковая система — это набор дифференцированных знаков, соответствующих дифференцированным понятиям. Это определяет язык «вообще», безотносительно к мышлению и представлениям (неотделимым от использования знаков в случае человеческого языка), к тому, стоят ли за знаками какие-то внутренние представления индивида, насколько осмыслены/осознанны их предъявление и смена по ходу общения или же (как и имеет место быть в коммуникации животных, осуществляющей кооперативную реализацию инстинкта разными индивидами-участниками соответствующ0их взаимодействий) всё происходит полностью автоматически, как модем считывает информацию в компьютерной сети и передаёт её дальше в интересах целого. Особи с их демонстрациями — узлы-ретрансляторы коммуникативной сети, «получающие» от участия в ней возможность выиграть территориальные или брачные взаимодействия, повысив тем самым свою дарвиновскую припособленность.
Возвращаясь от сугубой теории к конкретике наблюдений. Сигналы угрозы, ухаживания, предупреждения об опасности, выраженный соответствующей демонстрацией, влияет на поведение гораздо сильней, чем прямое действие агрессивного/сексуального характера, или прямое наблюдение хищника, разворачивающегося для атаки.
Тогда в самом общем плане коммуникация – это предсказуемое взаимодействие, в пределе – воздействие, целенаправленно регулируемое и контролируемое самими участниками, когда мощность внутренних систем управления и контроля превосходит мощность внешних воздействий (Кастлер, 1960).
Эту предсказуемость, а особенно – её непрерывное возрастание по ходу процесса вопреки безусловному антагонизму «интересов» особей Г.Кастлер (1960) рассматривал как главное доказательство существования информационного обмена в соответствующей системе взаимодействий. Рост предсказуемости носит взаимный характер: всем участникам коммуникации одинаково выгодно как передавать информацию о ситуации, так и получать её от другой особи, если первое и второе равным образом модифицируют поведение партнёра. Действительно, в том и другом случае прогнозируемость поведения одинаково растёт и в отношении полноты, и в отношении точности «прогнозов» (Кастлер, 1960), причём соответствующий рост устойчив к воздействию разнообразных «помех коммуникации» и стремлению участников всё время противодействовать демонстрациям друг друга….
…Сигналы как посредники упорядочивают и структурируют процесс взаимного выражения агрессивных (и любых иных) побуждений и намерений между членами определённого сообщества. В таких сообществах визуальные или акустические демонстрации (у млекопитающих – также запаховые или следовые метки) становятся посредником между намерением и действием в отношении другой особи и, соответственно, сигнализируют о наличии первых и вероятности вторых. Например, между агрессивным намерением и действием (удар, клевок, вытеснение или же, наоборот, отступление и бегство) или сексуальным (сближение с ухаживанием, попытка силового удержания самки и копуляции). Так, исследовали роль агонистических демонстраций глупыша Fulmarias glacialis как сигналов-посредников в столкновениях индивидов, скапливающихся вокруг выброшенных внутренностей пинагора (Cyclopterus lumpus). Каждому куску находится свой «бесспорный владелец»: он поедает корм и одновременно угрожает остальным, крутящимся вокруг и пытающимся вырвать часть или завладеть всем. В 22% наблюдений владелец не мог отстоять пищу и сменялся другим глупышом; вероятность смены очень слабо зависит от размера или пищевой ценности куска.
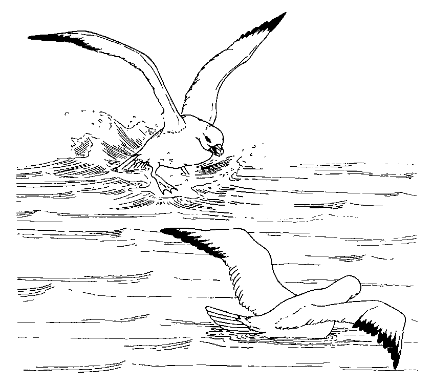
Взаимные угрозы глупышей. Налёт сверху со сгорбленной спиной (левая птица) и угроза раскрытыми крыльями (правая)
Эффективность угрожающих демонстраций определялась числом одержанных побед, риск – процентом случаев, вызвавших физическую борьбу птиц. Уровень агрессивности в столкновениях за пищу определяли, исходя из того, что голодный глупыш обороняет пищу дольше, чем сытый. Для всех выделенных демонстраций показана линейная зависимость эффективности и риска (Enquist et al., 1985).
Важно подчеркнуть: уровень агрессивной мотивации глупышей определяется именно готовностью пойти на бόльший (или меньший) риск демонстрирования, а не большим или меньшим проявлением открытой агрессии. Это означает «доверие» всех собравшихся птиц к эффективности демонстраций как ритуальных средств, используемых, чтобы противостоять отбиранию пищи. «Доверие» существует притом, что скопление образуется случайным образом, оно анонимно, его члены полностью независимы друг от друга, то есть оно необъяснимо исходя из давнего знакомства животных между собой и персонализованности их связей. «Нечестная» коммуникация в таких условиях абсолютно выгодна, и тем не менее наблюдается «доверие» к сигналам, а «обман отсутствует». Следовательно, названные демонстрации не просто имеют сигнальные свойства (которыми животные могут пользоваться или нет), они обладают сигнальной функцией, возникшей и совершенствующейся в эволюционном процессе. Поэтому она проявляется также при демонстрировании в условиях вроде вышеописанных, когда могла бы и не проявляться….

Ещё ритуализованные угрозы глупышей. Левая птица угрожает с воздуха, правая отвечает раскрыванием изогнутых крыльев
…Формирование специализированного «языка» (в смысле Соссюра — системы дифференцированных знаков, соответствующих дифференцированным значениям, причём в момент демонстрации знака существующим чисто потенциально, не ощущаемым непосредственно) предупреждения об опасности самок на гнёздах исследовали у самцов краснокрылых трупиалов (Agelaius phoeniceus). Весь сезон размножения самцы-владельцы территорий издают сигналы предупреждения об опасности. В каждый данный момент большинство из них издаёт один и тот же тип сигнала; если же в ответ на присутствие потенциального хищника какой-либо самец резко сменит его, его соседи сразу же переходят на новый вариант, чем ретранслируют сигнал по всему району возможного перемещения опасности.
Три ряда фактов подтверждают нам, что тревожные крики самцов сигнализируют самкам на гнёздах именно об опасности, не воспринимаемой непосредственно (а не «показывают» уровень возбуждения или беспокойства самого самца). Во-первых, лишь владельцы территорий слушают сигнал друг друга и немедленно реагируют на изменение типа вокализации. Нетерриториальные самцы издают точно такие же сигналы, но лишь при непосредственном столкновении с опасностью (очевидно, они не включены в коммуникативную сеть группировки). Во-вторых, резко отличается реакция самок на проигрывание повторяющихся и сменяющихся сигналов самца: они проявляли беспокойство и предпочитали оставаться возле своих гнёзд только во втором случае. В третьих, самки были более бдительны при отлучке самцов со своих территорий (что легко определить по отсутствию «повторяющихся» сигналов поблизости) и менее бдительны в их присутствии (Beletsky, 1989).
Полевые опыты показали, что действительно темп издавания криков тревоги кодирует информацию о дистанции до хищников или о степени опасности, исходящей от отдельных хищников. Для трупиалов, гнездящихся в густых зарослях, то и другое во многом – взаимозаменяемые параметры: разная степень опасности от потенциальных хищников требует разных дистанций тревоги и начала оборонительной реакции, но почти не требует запуска разных реакций беспокойства, отведения или бегства. Птицам предъявляли чучела ондатры, ястреба, виргинского филина и сороки на разном расстоянии от гнездовой территории, а также фиксировали темп вокализации в ответ на приближение наблюдателя. В обоих случаях темп тревожной вокализации был обратно пропорционален дистанции до потенциальной опасности, спад интенсивности подачи крика с
увеличением дистанции зависел от степени опасности (Beletsky, 1991).»
Из: Фридман В.С. «От стимула к символу». Сигналы в коммуникации позвоночных животных. Прим.публикатора.
7Релизер – ключевое понятие классической этологии. Это специфические морфоструктуры (брачные украшения, цветные пятна, кожистые выросты и пр.), телодвижения и их комплексы (особенно демонстрации) или (реже всего) действия (составляющие инстинктивный акт, вроде движения клюва гусыни, закатывающего обратно в гнездо выкатившееся яйцо). Все они благодаря специфической форме или характерному образу для рецептивного партнёра являются знаковыми стимулами и в качестве таковых используются в процессе коммуникации. Релизеры — это стимулы, самого факта предъявления которых уже достаточно для высвобождения ответной реакции, точно соответствующей стимулу, если животное находится в соответствующем мотивационном состоянии и рецептивно в отношении данного стимула.
Название происходит от англ. to release – высвобождать, калька исходного немецкого термина der Auslöser. Кроме «высвобождать», auslösen означает «запускать, включать, приводить в действие», и «сбрасывать бомбу в цель» [курсив мой – В.Ф.]. Все эти смыслы покрываются понятием социальных релизеров, как оно введено Лоренцем в работе «Компаньон в мире птиц» (Lorenz, 1935). Коммуникативная действенность релизеров состоит в немедленной «автоматической» выдаче специфического ответа на стимуляцию, который точно соответствует характеру стимула. «Знаковость» релизеров определяется тем, что воздействие оказывается именно формой демонстрации, специфический образ которой «антоматически» распознаётся реципиентом через врождённый разрешающий механизм, общий для всего круга потенциальных участников — или «зрителей» — взаимодействий данного типа. Специфическое воздействие стимула, могущее быть «уколом» или «давлением», лишь подкрепляет этот эффект; в предельном случае так называемых сигналов-символов (англ. referential sigmals) в нём нет необходимости, достаточно предъявить характерную и узнаваемую форму демонстрации, визуальной или акустической.
Точное соответствие стимула и реакции наиболее существенно в процессе коммуникации, когда обе особи используют дифференцированные наборы стимулов (демонстраций ритуализованной угрозы, ухаживания и пр.) для воздействия друг на друга, а специфические реакции особей также представляют собой демонстрации – элементы того же набора релизеров. Своевременная выдача демонстраций в соответствии с характером стимула позволяет процессу взаимодействия продолжаться и далее, направленно развёртываясь до некого биологически осмысленного результата.
8взаимно-обуславливающих друг друга через идею, позднее успешно подтверждённую данными, что инстинкты действуют не непосредственной стимуляцией, но специфической формой или образом, в меру устойчивости и точности её «черчения» образующими телодвижениями, т. е. пропорционально стереотипности исполнения демонстрации. И наоборот, ВРМ включают в себя системы распознавания образов, а их модели, отделяющие сигнал от шума с использованием искусственных нейронных сетей (Ghirlanda, Enquist, 1998), успешно используются в моделировании коммуникации животных.
«…Хотя форма демонстраций так или иначе варьирует, искажаясь при каждом акте воспроизведения сигнала, самообучающаяся нейронная сеть способна выделить и точно определить соответствующий сигнальный инвариант, эффективно дифференцировав его от инвариантов других сигналов ряда, и раз за разом точно распознавать его. Фактором искажения форм видовых демонстраций выступают
1) неустойчивость состояния особи (подверженного циклическим изменениям эндогенного характера и направленным – под действием стимуляции от противника); при том, что и сама воспроизводящая моторика варьирует от одного индивида к другому) и
2) изменчивость контекста общения в ситуациях, когда должен быть продемонстрирован и удерживаться определённый сигнал. Контекст общения быстро меняется, развиваясь в определенную сторону под влиянием событий, внешних относительно взаимодействующих особей (хищники, фактор беспокойства), и изменений в поведении партнёра, изменяющих характер, интенсивность стимуляции от него.
Оба фактора через «автоматические» изменения мотивации животного не могут не повлиять на моторику, воспроизводящую демонстрации, добавлять экспрессивности, амплитуды выразительным движениям и вообще всем реакциям особи. Тем не менее, особи с неустойчивым и меняющимся мотивационным состоянием воспроизводят специфические инварианты форм видовых демонстраций вполне устойчиво и эффективно дифференцируют их друг от друга – на уровне, достаточном для того, чтобы нейронная сеть могла распознать и разделить их (Enquist, Ghirlanda, 2005).
Предъявление выделенных «чистых форм» коммуникативных сигналов особям вызывает чёткую специфическую реакцию, более сильную и более устойчивую, чем на демонстрации собственного партнера, что доказывает правильность произведённого выделения «значащих структур» из потока действий животного. Наиболее убедительные опыты произведены с позывками гаичек Poecile и с визуальными демонстрациями агамы Amphibolurus muricatus (Nickerson et al., 2006; Peters, Davis, 2006)….». Фридман В.С., op.cit. Прим.публикатора.
9это и есть инстинкт — «…генетически первичная форма поведения, рассматривается как сложная структура, отдельные части которой слагаются наподобие элементов, образующих ритм, фигуру или мелодию» (Словарь Л.С.Выготского, 2004: 44). Прим.публикатора, курсив мой.
10вариант так называемой «вакуумной активности», die Leeraktivität. Она показывает, что побудительные механизмы реализации инстинкта внутренние, а не внешние, стимулы, и даже релизеры данные формы поведения только высвобождают (корректируют выбор ответа из нескольких форм демонстраций — возможных ответов в данном контексте, имеющихся в видовом репертуаре), но не формируют его своим действием. Прим.публикатора
11или, например, в плоскости, заданной положением одного из участников, которому адресуются ухаживания или угрозы — а он отвечает, как это имеет место у райских птиц, чудной Lophorina superba или синезатылочной Parotica lavesii (Frith, Frith, 1981) или в «классическом» случае ухаживания кряквы и других речных уток. Прим.публикатора
12Как я уже писал, при использовании её в данной работе выделены дискретные единицы и показана скачкообразность их смены в процессе ухаживания селезня кряквы за уткой. Т. е. считавшееся первоначально «фильмом» оказывается «слайдфильмом», где подобными методами этолог фиксирует реально присутствующие «слайды», они же демонстрации, и вправе полагать их существенность также для самих птиц. Прим.публикатора
13Schleidt 1973, p. 384.
14так бывает обычно из-за регулярности обмена криками, временной сопряжённости предъявляемого поведения и — если формы последнего меняются во времени — из-за направленности и устойчивости подобных смен (часто также с сохранением и временной сопряжённости, и согласованности по характеру изменения, от каких форм к каким из имеющихся в видовом репертуаре оно происходит). Прим.публикатора
Рекомендуем прочесть
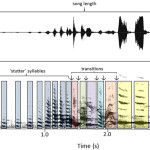 Пение самок воробьиных птиц: исключение или закономерность?
Пение самок воробьиных птиц: исключение или закономерность? Вольфганг Шлейдт, Ноа Эшкол и концептуальный кризис сравнительной этологии
Вольфганг Шлейдт, Ноа Эшкол и концептуальный кризис сравнительной этологии Как птицы обучаются инстинктивному поведению
Как птицы обучаются инстинктивному поведению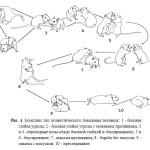 Синдром социальности: сравнительное исследование поведения песчанок
Синдром социальности: сравнительное исследование поведения песчанок Что не так с социобиологией?
Что не так с социобиологией? Так кто всё-таки сэкономил на мозгах?
Так кто всё-таки сэкономил на мозгах? Взлёт и падение универсальных «базовых эмоций» Экмана
Взлёт и падение универсальных «базовых эмоций» Экмана Насилие как коммуникация, и связь с угнетением
Насилие как коммуникация, и связь с угнетением
