![Print PDF В 1898 году Хермон Бампус впервые определил действие естественного отбора на дикую популяцию домовых воробьëв (1). Птиц, пойманных замерзающим в снежной буре, выхаживали в лаборатории. Выжили однако не […]](http://www.socialcompas.com/wp-content/uploads/2023/01/220886543_4154593707959463_6611152342217199361_n-465x190.jpg "Быстрая эволюция холодостойкости у анолисов в южном Техасе")

Бурый (Anolis sagrei) и североамериканский красногорлый (A.carolinensis) анолисы в позе угрозы
В 1898 году Хермон Бампус впервые определил действие естественного отбора на дикую популяцию домовых воробьëв (1). Птиц, пойманных замерзающим в снежной буре, выхаживали в лаборатории. Выжили однако не всё, причём выжившие воробьи были более длиннокрылыми и весили больше погибших. События, сходные с такой снежной бурей, преобразуют чувствительный вид быстро и мощно в демографическом и эволюционном отношении (1-4). Несмотря на их краткость, подобные катастрофы могут составлять большую часть общего селективного пресса, воздействующего на популяцию (5). Однако вопреки технологическому прогрессу и достижениям в анализе данных и статистических выводах [используемых в исследованиях] до с из пор удивительно мало эмпирических исследований естественного отбора, вызванного экстремальными погодными явлениями (1, 3, 4).
Еще хуже изучены регуляторные и генетические механизмы, на которые в этом случае «нацелен» отбор (6, 7) [и характер их изменений, обеспечивающий эффективное приспособление].
В работе Campbell-Staton et al. (2017) изучен отбор в популяциях американских красногорлых анолисов Anolis carolinensis Техаса, вызванный экстремальной волной холода 2013-2014 г., дошедшей до самого юга штата, и созданные им изменения на фенотипическом, регуляторном и генетическом уровнях. До воздействия, в августе 2013 г. авторы определили критический тепловой минимум (CTmin) в пяти популяциях с севера на юг штата Техас (рис.1А). СТmin равен температуре, при которой охлаждаемые ящерицы теряют координацию, в том числе не могут перевернуться обратно после помещения на спину (8).
Зимой 2013-2014 ослабление арктической зоны низкого давления (“циркумполярный вихрь”) вызвало резкое похолодание на всей территории южных штатов США (рис.1), в результате чего температурные минимумы были значительно ниже, чем в предыдущие 15 лет во всех 5 пунктах, от Браунсвилля на юге Техаса до Ходжена на севере [Тест Уэлча для двух выборок t; Браунсвилл, Техас (BRO): Р << 0,01; Виктория, Техас (VIC): Р = 0,047; Остин, Техас (AUS): Р = 0,021; Арлингтон, Техас (ARL): Р << 0,01; Ходжен, Оклахома (HOD): Р<<0,01].
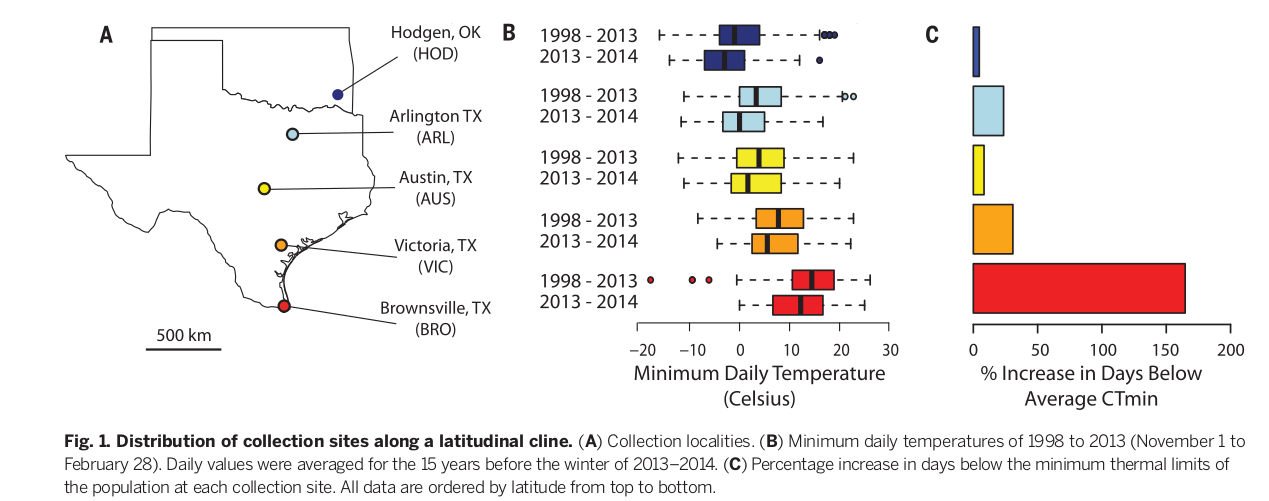
Рис.1. Места сбора материала вдоль широтной клины. (A) Места сбора. (B) Минимальные дневные температуры холодного периода с 1998 по 2013 год (с 1 ноября по 28 февраля). Данные усреднены за 15 лет, предшествующих зиме 2013-2014 годов (её температуры для каждого пункта даны во второй строчке) . (C) рост числа дней с температурой ниже СТmin в каждом из пунктов, проценты. Все данные упорядочены по широте сверху вниз.
Устойчивость ящериц к холоду естественным образом возрастала с юга на север штата (9). Тогда уровень стресса, вызванного данной волной холода, в каждом из пяти пунктов определяется числом дней, в которые ящерицы испытывали температуры ниже средних значений CTmin, отмеченных здесь же в предыдущие зимы. Наибольший прирост этих «дней холодового стресса» испытали самые южные популяции (BRO: 164,71%, 28 дней). Более северные также испытывали стресс, но меньший (VIC: 30,77%, 20 дней; AUS: 8,33%, 7 дней; ARL: 26,9%, 20 дней; HOD: 4,63%, 5 дней).
Авторы предположили, что подобно воробьям Бампуса, экстремальные холода элиминировали менее морозостойких особей, повысив устойчивость выживших к данному фактору экстремальности. Они изучили, так ли это в действительности.
При повторном посещении BRO и AUS в апреле 2014 года они определили минимальную температуру активности у выживших анолисов. Последняя почти везде снизилась, по уровню холодовой толерантности южные популяции приблизились к более северным до волны холода, и сильнее всего — самая южная популяция (BRO) (линейная модель смешанных эффектов, t = -2,09, P = 0,043). В центральных же популяциях (AUS) изменений не выявлено (t = -0,182, P = 0,856) (рис.2).
Чтобы исключить возможное объяснение происходящего сезонной пластичности, минимальную температуру активности ещё раз измерили в тех же пунктах. Если весенние изменения холодовой толерантности были б вызваны сезонной пластичностью, к лету значения этого параметра вернулись бы к прежнему уровню, особенно в Браунсвилле. Однако увеличение переносимости холода там сохранялось всё лето (t=-2,72, P= 0,009).
Авторы повторно посетили оставшиеся популяции в конце июля 2014 года, чтобы определить географическую вариабельность этого эффекта. Значительный рост переносимости холода выявлен также в VIC (t = -2,057, P < 0,05), но не в более северных районах штата (AUS: t = 0,116, P = 0,908; ARL: t = -0,818, P = 0,429; HOD: t = 1,064, P = 0,299). У зелёного анолиса холодовая толерантность наследуема (9) и изменяема в ходе локальных адаптаций (9, 10) поэтому сильный отбор по этому признаку может дать быстрый эволюционный ответ. Что и наблюдалось в действительности.
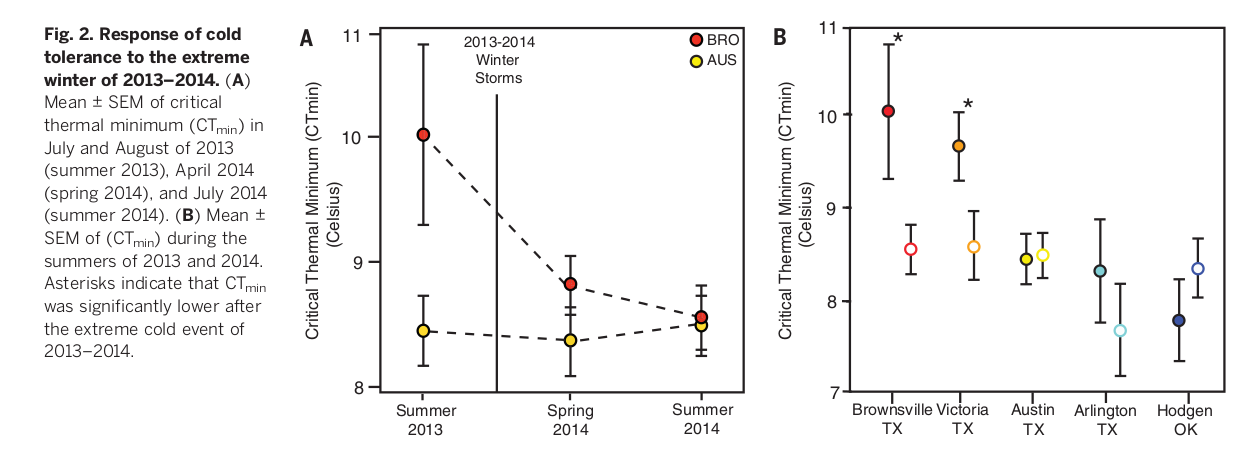
Рис. 2. Ответ изменением холодовой толерантности на экстремальную зиму 2013-2014 годов. (A) Средние значения ± SEM температурного минимума активности (CTmin) анолисов летом 2013 г. (в июле и августе), весной 2014 г. (апрель) и летом 2014 г. (июль). (В) Среднее значение ± SEM этого же показателя за лето 2013 vs лето 2014 годов для одних и тех же популяций (пустые vs заполненные кружки одного цвета). Звездочки – значимое снижение CTmin в наиболее южных популяциях после экстремального похолодания 2013-2014 гг..
Авторы упорядочили 48 транскриптомов из печени ящериц, собранных до и после фокальной зимы. Для этого перед экстремальными холодами они отобрали по 8 анолисов с обоих полюсов широтной клипы и 14-дней выдерживали в лаборатории при температуре 30°C (BRO: N = 4; HOD: N = 4) или 20°C (BRO: N = 4; HOD: N = 4), чтобы произошла акклимация. Еще 10 анолисов были пойманы в промежуточных пунктах клины перед экстремальной зимой и акклиматизированы до 30°C в течение 14 дней (VIC: N = 6; AUS: N = 4).
После этой зимы отловили 22 ящериц в 4х пунктах (BRO: N = 8; VIC: N = 6; AUS: N = 8), и тестировали после 14-дневного выдерживания при 30°C. Авторы использовали программу WGCNA (11) для идентификации модулей коэкспрессируемых генов во всех 48 транскриптомах печени, вариации коэкспрессии анализировали методом главных компонент. Мерой выраженности каждого модуля считали балл по первой из главных компонент, а дальше исследовали зависимость «интенсивности работы» каждого модуля от широты (рис.3). Всего идентифицировали 57 модулей коэкспрессии, три из них сильно и значимо связаны с исследуемым градиентом север-юг рис.1 (после контроля вариаций массы, пола и условий акклиматизации, таблица S1).
Дальше отслеживали изменения в экспрессии генов, вызванные экстремальным холодом — на особях, выживших после него и выдержанных при 30° C. У наиболее южных из них (BRO) экспрессия генов разных модулей сдвинута в основном в сторону средних уровней экспрессии самой северной популяции (HOD) (точный биномиальный тест; Модуль 16: P = 0,038; Модуль 18: P = 0,041; Модуль 53: P << 0,001) (рис.3). Эти сдвиги подтверждают гипотезу о том, что экстремальная зима 2013-14 гг. в самой южной популяции произвела отбор регуляторных фенотипов, наиболее сходных с таковыми более северных анолисов, чаще переносящих зимы сравнимой суровости. Данное изменение не было явственно в других популяциях (см. Дополнительные материалы), видимо, из-за более слабого воздействия экстремальной зимы (рис.1).
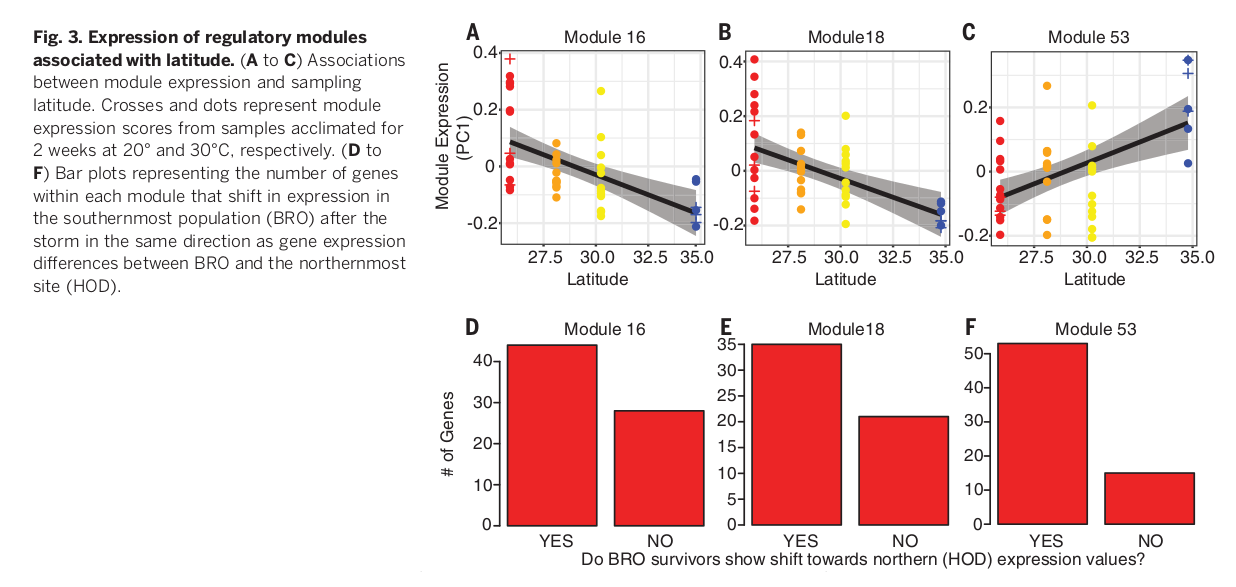 Рис. 3. Зависимость «интенсивности работы» (ордината) разных регуляторных модулей от широты (абсцисса). (A-C) Показаны аппроксимируюшие регрессии ± доверительный интервал. Крестики и точки — экспрессия данного модуля при акклиматизации в течение 2 недель при 20° и 30°C соответственно. (D-F) Столбцы — число генов каждого из 3-х модулей, чья экспрессия в наиболее южной популяции (BRO) после экстремальной зимы изменилась в ту же сторону, что различия в экспрессии генов между BRO и самой северной популяцией (HOD) в обычных условиях.
Рис. 3. Зависимость «интенсивности работы» (ордината) разных регуляторных модулей от широты (абсцисса). (A-C) Показаны аппроксимируюшие регрессии ± доверительный интервал. Крестики и точки — экспрессия данного модуля при акклиматизации в течение 2 недель при 20° и 30°C соответственно. (D-F) Столбцы — число генов каждого из 3-х модулей, чья экспрессия в наиболее южной популяции (BRO) после экстремальной зимы изменилась в ту же сторону, что различия в экспрессии генов между BRO и самой северной популяцией (HOD) в обычных условиях.
Далее авторы определили вероятные геномные мишени отбора, вызванного волной холодов, для наиболее южной популяции (BRO). Они сопоставили данные секвенирования РНК у 8 ящериц, собранных до и после зимы 2013-2014 гг., отыскивая геномные регионы показывающие значимый рост различий «текста» РНК (сила и значимость этих выбросов оценивались по Fst). Всего их набралось 14 (рис.4А). 4 участка из них также показывают дивергенцию между Браунсвиллом и самой северной популяцией клипы (рис.4А). Видимо, приспособление к одноактному холодовому стрессу затронуло и участки генома, вовлеченные в приспособление к устойчивым климатическим различиям между севером и югом Техаса. Абсолютная дивергенция, dXY (12), была значительно выше у выбросов Fst южных популяций по сравнению с северными (среднее значение внутри них dXY= 0,00063, вовне dXY= 0,00054; критерий суммы рангов Уилкоксона: Р = 0,01). Исходя из этого можно заключить, что наблюдаемые паттерны дифференциации не артефакт «попадания» в геномные области с низким разнообразием (12).
Дифференциально экспрессируемые гены в «местах» выбросов (N = 12) были обогащены активностью таких ферментов, как гидролазы карбоновых эфиров (геномная область GO:00052689, P = 0,012) и нейромедиаторные натриевые симпортеры, NSS (GO:0005328, P = 0,014; GO: 000345, P = 0,016). Они участвуют в поддержании синаптической функции, предполагается, что они вносят вклад в вызванную температурой потерю функции и смерть у эктотермических видов (13). Трое из этих генов [ацетилхолинэстераза, LOC100557088 (холинэстераза) и LOC100561946 (холинэстераза)] — ортологи ацетилхолинэстеразы, играющей важную роль в передаче нервного импульса (14). Кроме того, два переносчика гамма-аминомасляной кислоты (ГАМК) (SLC6A1 и SLC6A8), находящиеся внутри наиболее значительных выбросов, участвуют в обеспечении активности NSS.
ГАМК — это тормозящий нейромедиатор, критически важный для поддержания мышечного тонуса. Все пять генов расположены на 2й хромосоме генома зеленого анолиса, в одном большом выбросе (рис. 4B-F). Дифференциальная экспрессия этих генов в печени может отражать различия функционирования всей вегетативной нервной системы. Требуются дальнейшие исследования для изучения потенциальной роли данных генов в функционировании организма в зависимости от охлаждения.
Экстремальные погодные явления учащаются и утяжеляются в связи с развертывающимися антропогенными изменениями климата. Эти события могут радикально влиять на природные популяции, вызывая мощные селективные давления и стимулируя современную эволюцию (16). Понимание биологических последствий этих событий имеет важные последствия для дальнейшего выживания видов по всему земному шару. Это исследование демонстрирует, что такие события могут в наблюдаемой популяции быстро произвести отбор на фенотипическом, регуляторном и генетическом уровнях, и привести к паттернам дивергенции состояния «после воздействия» от состояния «до», однотипным с той дивергенцией, которая обусловлена локальной адаптацией, развивающейся медленно и постепенно вдоль естественных климатических градиентов.
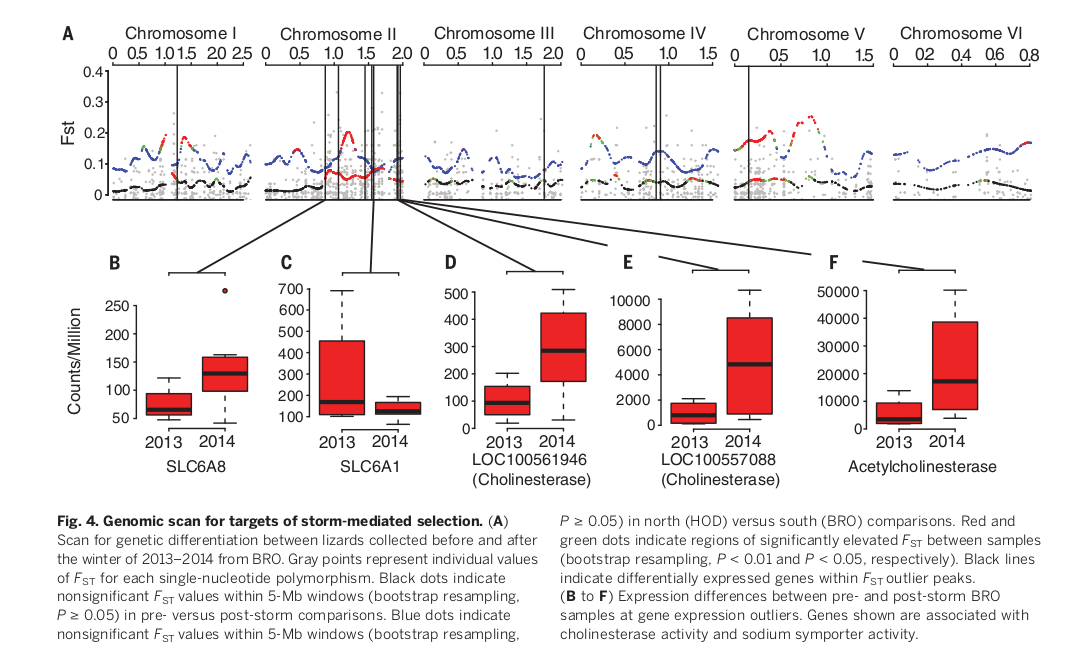
Рис.4. Геномное сканирование для поиска мишеней отбора, произведённого экстремальным холодом. (А) Места генетической дифференциации между ящерицами, собранными до и после зимы 2013-2014 годов из BRO. Серые точки — индивидуальные значения Fst для каждого однонуклеотидного полиморфизма. Черные точки — незначимое FST в пределах 5-мегабайтных окон (бутстреп, P ≥ 0,05) для сравнений до и после шторма. Синие точки — незначимое Fst в пределах 5-мегабайтных окон (bootstrep resampling — имитация множества случайных выборов и распределения результатов для этого множества, при многократной генерации выборок, P ≥ 0,05) в сравнениях северных популяций анолисов (HOD) с южными (BRO). Красные и зеленые точки указывают на области значительно повышенного Fst между выборками (бутстреп, P < 0,01 и P < 0,05 соответственно). Черные линии указывают на дифференциально экспрессируемые гены в пределах более высоких пиков. (B-F) Различия в экспрессии генов у анолисов BRO до и после экстремальной зимы в тех участках генома где отмечены выбросы. Показанные гены связаны с активностью холинэстеразы и активностью симпортера натрия.

Ящерица показывает камуфляжную окраску. Белая полоса по хребту у самок
Рекомендуем прочесть
 «Гены урбанизации» и приспособление к средовому стрессу
«Гены урбанизации» и приспособление к средовому стрессу Экспериментальная эволюция гуппи в речках Тринидада.
Экспериментальная эволюция гуппи в речках Тринидада. Быстрая эволюция в связи с урбанизацией: примеры
Быстрая эволюция в связи с урбанизацией: примеры Мини-рецензия на книгу Менно Схилтхёйзена «Дарвин в городе. Как эволюция продолжается в городских джунглях»
Мини-рецензия на книгу Менно Схилтхёйзена «Дарвин в городе. Как эволюция продолжается в городских джунглях» Эпигенетическое программирование фенотипических различий в поведении и эволюция социальности у грызунов
Эпигенетическое программирование фенотипических различий в поведении и эволюция социальности у грызунов Эволюция в пространстве возможностей: Дарвин и Вавилов
Эволюция в пространстве возможностей: Дарвин и Вавилов Дарвиновская эволюция и регуляторные генетические системы
Дарвиновская эволюция и регуляторные генетические системы Эволюция на двух уровнях: о генах и формах
Эволюция на двух уровнях: о генах и формах
