

Ландшафтный парк «Троицкая роща», г.Троицк. Рекреационный лес, «захваченный» внутрь урболандшафта
Резюме. Городские экосистемы отличаются несколькими парадоксами; в работе И.А.Жигарева показана их причина — высвобождение ресурсов нарушением. Умеренное нарушение повышает продуктивность главнейших компонентов экосистемы, увеличивает биоразнообразие и вместе с притоком органического вещества и энергии в город из области экологического следа объясняет прочие парадоксы.
Исследование природы сообществ является одним из центральных направлений в экологии, как в прошлом, так и в настоящем.
Представления об организации реальных сообществ во многом строятся на общесистемных позициях. В основе их лежит теория конкуренции, блестяще подтвержденная экспериментами Г.Ф. Гаузе (1934, 1935), а позже Т. Парком (1948, 1954) и многими другими исследователями.
Сформированный на основе этих работ «принцип конкурентного исключения», или принцип Гаузе и есть тот фундамент, который, на первый взгляд, должен во многом объяснять структуру реальных сообществ. Г.Ф. Гаузе сформулировал ныне известный всем принцип, суть которого выражается в том, что
«Два конкурирующих вида вместе не уживаются».
Позже, связав конкуренцию с понятием экологической ниши, принцип стал формулироваться следующим образом
«Два вида, занимающие одну и ту же экологическую нишу, не могут устойчиво сосуществовать».
Суть этих представлений, так же как и общесистемных, логических допущений, в том числе математических моделей, говорят, что два разных вида, потребляющих один и тот же ресурс, не могут его потреблять одинаково – один должен делать это эффективнее другого. Почему? Потому что виды разные, и они не могут быть одинаково приспособлены к использованию этого ресурса. Конкуренция всегда асимметрична. Именно это и должно привести к вытеснению. Такую ситуацию можно назвать эффектом монополизации ресурса.
В этом, если хотите, смысл существования видов. Одно из логических следствий этого эффекта – уменьшение разнообразия внутри сообщества в экологическом масштабе времени.

Чарльз Сазерленд Элтон, создатель концепции экологической ниши как «профессии вида в сообществе»
С другой стороны конкурировать видам «не выгодно», это затратный процесс, даже в случае победы одним из видов и захвата всего ресурса. Поэтому в эволюционном масштабе времени, конкуренция должна приводить к сегрегации сходных видов и дифференциации их по градиенту ресурса. Логическим следствием этого положения является вывод, озвученный Ч. Элтоном (Elton, 1958, 1966) и Г. Хатчинсоном (Hutchinson, 1959), что близкородственные виды должны чаще размежевываться и иметь разные экологические ниши.

Джордж Эвелин Хатчинсон, автор концепции многомерной ниши (экологическая ниша как местообитание вида)
Однако с самого начала формирования этих представлений, в экологии сообществ стали накапливаться фактические полевые материалы, доказывающие, что близкородственные виды довольно часто сосуществуют совместно на тех или иных территориях. Более того, статистические оценки вероятности встреч видов одного рода в пределах сообщества часто оказываются выше, чем при случайном их распределении (обзоры Азовского, 1990; Шварца, 2004: и др.). Такое положение позволяет сделать эмпирическое заключение, что коль близкородственные виды должны быть близкими и экологически, следовательно, мы вправе ожидать их частое совместное обитание и сосуществование.
Критика принципа Гаузе началась буквально с момента его становления. В 1961 году Хатчинсон формулирует «Планктонный парадокс», суть которого заключается в том, что на поверхности водоемов нередко сосуществуют вместе десятки видов фотосинтезирующих организмов, не стратифицированных в пространстве.
Некоторые полевые факты также трудно объяснить с позиции «Принципа конкурентного исключения». Например, организацию луговых фитоценозов, где виды распределены по ограниченному количеству осей ресурсов нишевого пространства (в основном свет, вода и элементы минерального питания). В этих фитоценозах рядом всегда сосуществуют очень близкие и экологически схожие виды. На основании принципа Гаузе, это кажется вообще необъяснимым.
Несколько позже П. Де Бур (Den Boer, 1985) предложил даже заменить классический «принцип конкурентного вытеснения» на «принцип сосуществования». А некоторые исследователи, например Марков, даже предложили отказаться от принципа Гаузе, как не отвечающего полевым фактам и возможностью неоднозначной трактовки данных, подтверждающих принцип (Марков, 2004).
Более того, накопившиеся проблемы в теории конкуренции зачастую приводят к позиции отрицания конкуренции в природе, или, по крайней мере, в отсутствии необходимости использовать основные положения самой теории в объяснении дифференцирования видов в эволюции или сегрегации их в сообществах (Simberloff, 1970; Simberloff, Boecklen, 1991; Марков, 2004; и др.).
Нужно сказать, что не последнюю роль в негативном восприятии теории конкуренции ее противниками, играет частое и свободное упоминание термина «конкуренция», когда исследователи бездоказательно, априорно указывают на конкуренцию между видами, только потому, что они обитают совместно. Наличие или отсутствие конкуренции нужно доказывать, а это не всегда просто.
Так или иначе, все это существенно подрывает авторитет заявленного Гаузе «принципа конкурентного исключения» и требует подробного анализа существующего положения. Кто же прав в этом споре?
Во-первых, базовым принципом правила Гаузе было и остается положение о лимитировании ресурса. Это положение было осмыслено не сразу и пришло в экологию лишь к 50-м годам ХХ века, благодаря стараниям Хатчинсона. Эксплуатационная конкуренция имеет место только тогда, когда ресурс, за который происходит конкуренция, лимитирован. В противном случае конкуренции нет, и вытеснения, естественно, также быть не может. Мы наблюдаем реальное сосуществование видов. С такой поправкой принцип Гаузе будет звучать следующим образом –
«Два вида, занимающие одну и ту же экологическую нишу, не могут устойчиво сосуществовать, если ресурс лимитирован».
Очевидно, что общее число устойчиво сосуществующих видов не может превышать числа ограничивающих их ресурсов. Итак, мы можем наблюдать взаимное сосуществование, при котором вытеснения не происходит, и в то же время это не противоречит принципу Гаузе по следующим основным причинам.
Во-первых, вытеснения не происходит, если ресурс или ресурсы, за которые виды соперничают, находятся в избытке.
Ситуация, когда объем ресурса избыточен, часто недооценивается исследователями. Теоретически, в этом случае конкуренция должна отсутствовать, независимо от количества сообщество видов. В этом случае виды сосуществуют совместно, а их набор может быть во многом случаен. Поэтому, при выявлении роли конкуренции в сложении сообщества, желательно оценивать объемы ресурса, в противном случае, говорить о конкуренции сложно. Реальное перекрывание экологических ниш, без оценки ресурсов, не дает оснований утверждать о наличии конкуренции. Конечно, теоретически, ситуация избыточности ресурса долго существовать не может. Со временем, через дифференциальное размножение и миграцию, излишки ресурса должны быть изъяты. Однако периодические изменения в природе, ограничивающие численность видов, например, сезонность климата, могут препятствовать достижению насыщения.
Сильный конкурент просто может не успеть захватить весь ресурс. Условия обитания видов в природе никогда не бывают постоянны. Поэтому избыточность ресурса, видимо, явление реальное, хотя бы временно. Например, для грызунов обилие пищевого ресурса во время созревания урожая, или интенсивного нарастания вегетативной массы растений вполне может быть избыточно. Возможно, что отдельные наблюдения в природе сообществ, насыщенных близкими видами, может быть следствием такого положения. Особенно это касается сообществ зеленоядных видов. Кстати большинство специалистов, которые опровергали Гаузе изучали растительноядных и всеядных организмов – насекомых (Марков), жужелиц (Де Бур), муравьев (Симберлофф), и др.

Нельзя забывать и то, что в жизни любого вида наиболее существенным фактором является ограничивающий фактор (значение которого находится в недостатке или в избытке). Именно он лимитирует рост численности вида. Соответственно, нет ничего необычного в том, что в отдельных случаях, на протяжении длительного времени, в условиях подавления численности при воздействии какого-либо фактора (не ресурсного), создаются предпосылки, в результате которых по другим осям нишевого пространства формируется избыточность ресурсов.
Остановлюсь лишь на арвалоидных (серых) полевках. Многочисленные данные по распространению обыкновенной и восточноевропейской полевок при совместном обитании показывают низкий уровень биотопической сегрегации этих видов. В наших многолетних исследованиях у этих двух видов просматриваются лишь отдельные тенденции в предпочтении биотопов, в отличие от других грызунов, потребляющих более концентрированные корма. Эти виды без сомнения экологически близкие, однако используемые ими ресурсы (пища и места для убежищ), как правило, находятся в избытке, следовательно, их сосуществование вполне возможно и без конкуренции. С другой стороны, регулирующие факторы (видимо, в первую очередь, хищники и паразиты) большую часть времени вполне эффективно контролируют их численность, не позволяя, тем самым, значительно сокращаться пищевым ресурсам и местам убежищ этих полевок. Массовые вспышки численности серых полевок в открытых биотопах косвенно доказывают это положение. Сюда же можно отнести и отсутствие выраженных внутрипопуляционных регулирующих механизмов у этих полевок, в отличие от тех же рыжих полевок или мышей.
Это адаптации к условиям безлимитных ресурсов, что позволяет им без существенной конкуренции между собой обитать в одних и тех же местообитаниях. Для потребителей более концентрированных кормов, особенно землероек, напряженность по пище, видимо, более обычное положение, особенно зимой и весной.
Второй причиной сосуществования без конкуренции является разделение экологических ниш видов (например, они могут отличаться по спектру пищевых объектов, по способу добывания корма или активны в разное время суток). Именно это положение дает нам наибольшее количество примеров, объясняющих устойчивое сосуществование экологически близки видов в пределах одного сообщества, без «нарушения» правила Гаузе. Структура любого природного сообщества довольно сложна. И детальные описания экологических ниш близких видов часто приводят к выявлению отличий их параметров, которые были не видны в первом приближении. Это было показано на различных видах. В нашу задачу не входит необходимость подробного разбора этих примеров, которые обильно представлены в многочисленных сводках по конкуренции.
Исходя из общетеоретических положений – конкуренция энергетически затратный процесс для всех участников взаимодействия. Поэтому длительно конкурировать видам невыгодно. Эволюционно эта ситуация должна развиваться по двум сценариям: либо один вид вытесняет другой, вплоть до его вымирания (или миграции), и монополизирует ресурс, либо они расходятся по градиенту ресурса, монополизируя каждый свой участок, то есть специализируются. Кстати, слабые конкуренты (эксплеренты, сорняки, ценофобы), которые обычно являются неспециализированными формами по использованию пищи и пространства, нередко оказываются высокими специалистами по временной оси ниши. Ибо они захватывают ресурсы до прихода туда сильных конкурентов и имеют многочисленные приспособления для подобной стратегии. Так или иначе, расхождение близких видов требует какого-то эволюционного времени и, видимо, достаточно продолжительного, чтобы выработались соответственные адаптации.
Следовательно, мы вправе предположить, что продолжительные конкурентные отношения свойственны молодым сообществам, а также тем, которые испытывают внедрение чужеродного вида (видов). В то же время, в группировках, где виды долго сосуществуют, мы с большей вероятностью должны наблюдать отсутствие конкуренции, ее низкий порог или кратковременные эпизоды конкурентных отношений в критические периоды лимитирования ресурсов.
Третья причина сосуществования без конкуренции заключается в использовании видами различных микроместообитаний внутри выделенного биотопа. Расхождение видов по микроместообитаниям, как и по структурным параметрам ниши, наиболее убедительный механизм формирования гильдий. И примеров здесь много, остановлюсь лишь на некоторых методических подходах.
Нередко наши представления о биотопических предпочтениях видов базируются на формальных подходах, удобных исследователю. Большое количество работ, в которых исследователь переходил от крупных биотопических выделов к более дробным, демонстрируют падение сходства в распределении близкородственных видов в пространстве. Это было показано на мелких млекопитающих Е.А. Шварцем (2004), Б.И. Шефтелем (1990), И.Ю. Поповым (1998), и нами (Жигарев, 1993, 2004), а для различных групп беспозвоночных А.И. Азовским (2007) и мн. др.
Это выражается в «ошибках» масштаба выделения биотопов, чем «грубее» мы выделяем неоднородности местности, тем больше вероятность не заметить видовых предпочтений пространственных выделов. Например, грубое выделение биотопов, таких как «лес», «луг», «болото» и так далее, и даже более дробных – «еловый лес», «сосновый лес», без детального описания травянистой растительности может скрыть реальные биотопические предпочтения видов, обитающих в них. Стоит изменить масштаб выделения местообитаний: перейти от крупных выделов «типа леса» к более мелким выделам – парцеллам растительных ассоциаций, как математическое сходство распределений экологически близких видов существенно падает.
Так, наш анализ более 15 тысяч поимок близкородственных видов зверьков на юге Подмосковья подтвердил данные Е.А. Шварца (2004) по Валдаю. Индексы перекрывания между близкими видами при переходе от крупных выделов к парцеллам падает от 20 до 70%. Например для пары желтогорлой и лесной мыши он «сократился» практически на 20% (отличия достоверны, при р = 0,008), между обыкновенной и малой бурозубками на 10% (р = 0,05). У редких видов индекс перекрывания с другими видами «упал» еще больше, например, в паре мышь-малютка – лесная мышь более чем на 70% (р = 0,002), мышь-малютка – желтогорлая мышь, на 68% (р = 0,002), а орешниковая соня – лесная мышь на 74% (р = 0,00005).
Другими словами, обитая в одном пространстве, которое исследователь выделяет как более-менее однородное, виды существенно расходятся по микроместообитаниям, предпочитая разные парцеллы, или их комплексы таким образом, чем мельче размеры выделяемых биотопов, которые использует исследователь, тем, с математической точки зрения, экологически близкие виды «сосуществуют» в кавычках реже.
Четвертая причина сосуществования видов без противоречия принципу Гаузе связана с нестабильностью среды, в которой условия постоянно меняются, становясь благоприятными то для одного, то для другого конкурирующего вида. Здесь может сформироваться равновесие и окончательного конкурентного вытеснения не происходит. Виды будут сосуществовать неопределенно долго (пока имеются циклические смены условий).
Такая ситуация будет устойчивой, и сформируется равновесие только в том случае, если преимущество одного вида перед другим будет недостаточно продолжительным, чтобы конкурентное вытеснение завершилось. Исходя из общих (но не проверенных) соображений, хрупкость такого сообщества чрезвычайно высока.
Доказательства подобного сосуществования видов были получены еще в классических экспериментальных работах Т. Парка (Park, 1948) на мучных хрущаках. Возможно, что планктонный парадокс связан именно с этой особенностью среды.
Наконец, конкурентного вытеснения может не происходить, если численность конкурентно более сильного вида постоянно ограничивается хищником, паразитом или третьим конкурентом (конкуренты за иной ресурс). Они, выступая в роли регулирующих факторов и снижая численность конкурирующих между собой видов (жертв), способны высвобождать часть ресурсов, не допуская их лимитирования, тем самым уменьшить интенсивность конкуренции между видами и способствовать увеличению видового разнообразия в сообществе.
Вообще, любой ограничивающий численность потенциальных конкурентов фактор (кроме ресурсного), как-то: враги (хищники, паразиты, третьи конкуренты), нестабильность среды, и другие, при умеренном воздействии и при всех прочих равных условиях должны увеличивать разнообразие сообщества, так как при этом высвобождается часть ресурса (ресурсов).
Есть еще одна причина, которая приводит к «видимой» поддержке принципа сосуществования и якобы противоречит принципу Гаузе. Это смены сообществ. Очевидно, что всякие смены, обусловленные внешними или внутренними причинами, редко когда происходят мгновенно. Для завершения этого процесса требуется время, возможно, соизмеримое с продолжительностью жизни основных членов сообщества. Однако большинство экологических исследований представляют собой «моментальные съемки» и строятся на допущении о некоторой неизменности исследуемых группировок. Несложно предположить, что наблюдение переходных (во времени) группировок с видами «реликтами»
и «пионерами» в момент их смены может быть расценено как относительно «стабильное» сообщество, если продолжительность его смены превышает продолжительность наблюдений. Естественно, что в такой временной, смешанной группировке видовое разнообразие должно быть более высоким, по сравнению с крайними сообществами. Таким образом, арена временного сосуществования видов, на которой происходит конкурентное вытеснение одной группировки другой и где «работает» правило Гаузе, мы наблюдаем пусть и кратковременное, но повышенное биологическое разнообразие, что может быть принято за сосуществование. Если учесть, что в пространстве такие «смешанные» группировки могут повторяться и возникать вновь и вновь на новых участках, то впечатление об их стабильности может только укрепиться. Возможно, что большая часть примеров «опушечного эффекта» относится к этим условиям.
Наконец есть еще одна причина, позволяющая объяснить механизм сосуществования без критики принципа Гаузе, и заключается она в феномене нарушения сообщества (Жигарев, 2005). Мы в течение более 30 лет исследовали мелких млекопитающих в рекреационно нарушенных лесах Подмосковья. Рекреационное нарушение лесных сообществ легко стадируется и четко описывается. Тщательно подобранные серии площадок в точках с разной степенью нарушенности позволяют выявить интересные закономерности изменения видового состава млекопитающих. Нами в Подмосковье отмечено 18 видов мышевидных грызунов и насекомоядных.
Анализ более чем 40 тысяч поимок зверьков демонстрирует убедительное доказательство закономерного изменения видового богатства мелких млекопитающих по градиенту рекреационного нарушения. Независимо от исходного типа биотопа – на средних стадиях нарушенности можно наблюдать существенный рост числа видов. Это происходит за счет внедрения в лесной комплекс видов антропофильного компонента – полевой мыши, обыкновенных и восточноевропейских полевок, серой крысы, домовой мыши, малой белозубки. На более поздних стадиях дигрессии часть лесных видов не выдерживает нагрузок и исчезает, к ним относятся темные полевки, лесные мышовки, малая бурозубка, орешниковая соня, водяная кутора. Что приводит к падению общего числа видов. Изменение индексов видового разнообразия мелких млекопитающих по градиенту рекреационной дигрессии, так же, в общих чертах, повторяет особенности изменений видового богатства.
Аналогичные изменения видового богатства и разнообразия при рекреационных нарушениях можно наблюдать и в растительных сообществах (см. обзор Жигарев, 1993).
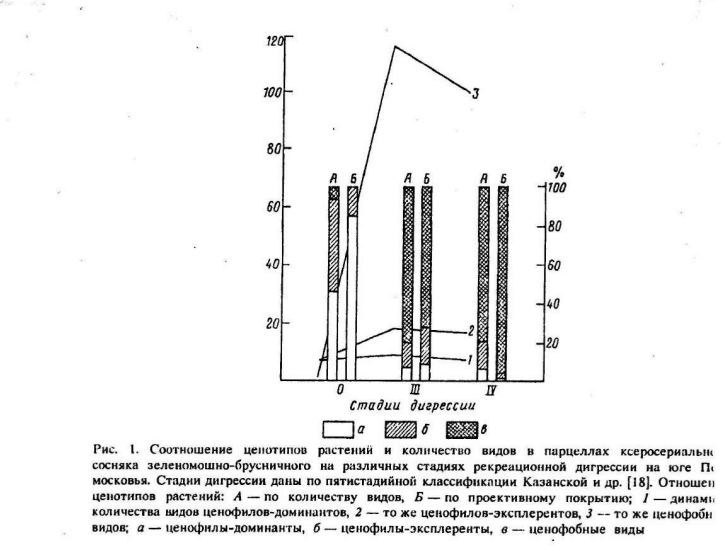
Источник. Жигарев И.А. Закономерности рекреационных нарушений биогеоценозов // Успехи совр. биологии. 1993. Т.113. вып.3
Кумулятивная (накопительная) рекреация вызывает существенное изменение в соотношении фитоценотипов растительных ассоциаций: доля ценофильных видов прогрессивно уменьшается вследствие их уничтожения, а доля ценофобов увеличивается в ходе внедрения новых видов сорняков. За счет внедрения ценофобов на начальных и средних стадиях рекреационной дигрессии сообществ возрастают структурные показатели фитоценозов – флористическое богатство, проективное покрытие, фитомасса и другие, а на последних – соответственно падают (Казанская и др., 1977; Жигарев, 1993; и др.).
Обычно принято считать, что наибольшие показатели видового богатства и разнообразия присущи природным сообществам. Такое положение во многом базируется на сравнении естественных и крайних, как правило, сильно деградированных антропогенных группировок видов. Однако, когда исследователь «не пропускает» промежуточные стадии деградации, то однозначной картины не наблюдает (Жигарев, 2004).
Схожая картина изменения видового богатства при нарушениях отмечена многими исследователями, изучающими не только млекопитающих, но и, например, птиц, почвенных беспозвоночных, грибов и так далее. Любопытно, по данным О.Е. Марфениной (2005), увеличение разнообразия микроскопических грибов при рекреационном нарушении происходит за счет нетипичных для данной зоны и субстрата видов, как правило, южных регионов, так же как и млекопитающих. Общеизвестны факты существования принципиально более высокого видового разнообразия фитоценозов умереннонарушенных лугов (выпас и сенокошение) по сравнению с естественными лесами и лугами (Вальтер, 1982; Работнов, 1983; Андреев, 1985; Горчаковский, 1987; и мн. др.).
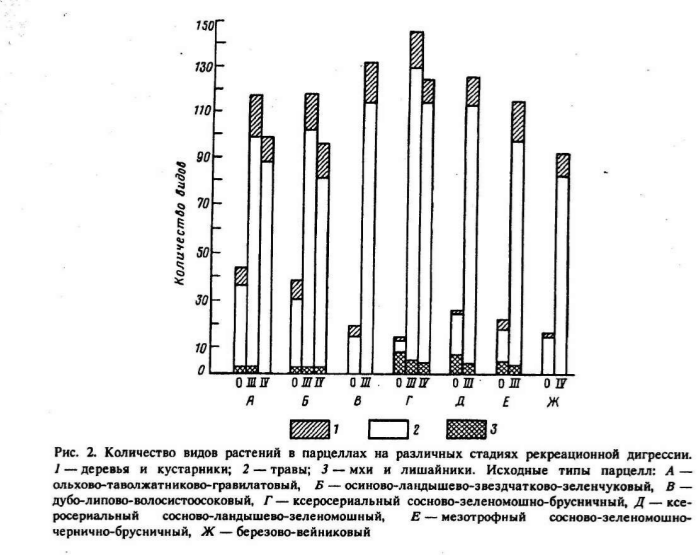
Источник. Жигарев, 1993, op.cit.
Широко известен факт, наблюдаемый при организации заповедного режима и снятия антропогенного нарушения, например сенокошения или выпаса, что нередко приводит к потере биологического разнообразия и усиления монодоминантности. Работы луговедов показывают, что умеренные нарушения – основа разнообразия многих луговых ассоциаций. Парадокс разнообразия луговых сообществ в их постоянном нарушении (сенокос, выпас, палы, паводки и др.).
В случае, когда умеренные нарушения (напрямую или опосредованно) подавляют в сообществе сильного конкурента, это способствует увеличению численности и благополучия других, более «слабых» членов сообщества, а также способствует возможному внедрению новых видов, потребляющих тот же ресурс. Следствием этого является усиление выравненности и повышение видового разнообразия. Наблюдается сосуществование при сильной конкуренции. Подобный эффект увеличения разнообразия при нарушениях можно назвать «эффектом высвобождения нарушением» (Жигарев, 2007). Это положение ни в коей мере не противоречит правилу Гаузе, а лишь дополняет его.
Совсем другие последствия должны происходить с сообществом, если воздействие затрагивает не доминанта, а его более слабых конкурентов. В этом случае мы вправе ожидать усиление позиций конкурентного доминанта, который максимально выбирает освободившийся ресурс, и, в результате, должно формироваться менее разнообразное сообщество.
Именно эффектом высвобождения ресурса при нарушениях мы объясняем повышение видового богатства мелких млекопитающих в нарушенных лесах. К аналогичному выводу о росте видового разнообразия при нарушениях пришел J. Connell (1978), который сформулировал «гипотезу промежуточного нарушения». Поддержание видового разнообразия конкретного сообщества на относительно высоком уровне возможно, по всей видимости, только при умеренном воздействии. Низкий уровень воздействия экзогенных факторов может быть слишком слабым, чтобы ограничить мощь конкурентных доминантов и подавить их господство. Слишком высокий уровень может существенно изменить условия обитания для всех членов сообщества, вплоть до их вымирания или миграции, что послужит причиной резкого видового обеднения.
На основании изложенных выше поправок к закону Гаузе, объясняющих сосуществование видов без конкурентного вытеснения, и трудностей в выявлении конкуренции в природе, никак нельзя отрицать сам закон Гаузе как один из фундаментальных законов, формирующих природные системы, как это делают некоторые авторитетные исследователи (Simberloff, 1970; Strong, et al., 1979; Strong, 1982; Simberloff, Boecklen, 1991; Марков, 2004 и др.).
Все описанные выше поправки мешают «включиться» в действие конкурентному вытеснению, но ни в коей мере не отрицают самого принципа Гаузе, поэтому выдвигаемой альтернативы – «сосуществование или вытеснение» – как таковой, нет. Следствием конкурентного вытеснения является уменьшение разнообразия в сообществе за счет удаления более слабых конкурентов. Все факторы, способствующие подавлению этого процесса, будут способствовать, при всех прочих равных условиях, увеличению разнообразия в сообществе (гильдии). К ним можно отнести хищников, паразитов, третьих конкурентов (конкурентов по другим ресурсам), нестабильность среды, умеренные нарушения и др. Так или иначе, все они способствуют подавлению сильного конкурента (конкурентов) и высвобождению ресурсов, а их избыток – увеличению разнообразия. Ограничением этого эффекта будет являться чрезмерное проявление перечисленных факторов. Очевидно, что это будет способствовать уменьшению разнообразия.

В стабильной и ненарушенной среде, при всех прочих равных условиях, будут сформированы относительно бедные сообщества. Это положение нередко упускается из виду исследователями.
Вторым следствием конкурентного вытеснения является дифференциация видов по осям нишевого пространства, в том числе и микроместообитаниям, что приводит к их сегрегации. Таким образом, чем исходно разнообразнее структура среды, тем более разнообразно видовое богатство данного сообщества. Очевидно, что достигнуть ситуации снятия конкуренции путем разделения ресурсов можно лишь по прошествии какого-то времени. А сами сообщества должны иметь эволюционную историю и формироваться как в результате конкурентного вытеснения в прошлом, так и в результате внедрения аллопатрично сформированных видов.
Следствием такого положения является более эффективное использование ресурсов среды, обусловленное специализацией (Вахрушев, Раутиан, 1993). Вторая позиция, с внедрением «готовых» видов, пусть и реальная, и подтвержденная примерами (Северцов и др., 1998; Шварц, 2004), но все-таки редкая.
Эволюционно молодые сообщества вовсе не обязательно должны иметь бедный видовой состав. Главным их отличием от эволюционно зрелых (не путать с сукцессионно зрелыми) должен быть случайный, и, видимо, непостоянный набор видов, пониженная специализация и высокий уровень конкурентных отношений.
Исходя из гипотезы сегрегации (конкурентного отталкивания), этот уровень со временем должен падать. Здесь можно согласиться с гипотезой М. Коди (Cody, 1973), считающего, что конкуренция характерна для относительно молодых сообществ, не завершивших свое формирование, в то время как в зрелых, окончательно устоявшихся, конкуренция отсутствует или является слабой, ввиду разделения используемых ресурсов.
Следует отметить, что для объяснения специфики организации сообществ одной только теории конкуренции недостаточно. Она во многом объясняет структуру гильдий, ассамблей, таксоценов, то есть сообществ, в которых преобладают «горизонтальные» связи, за счет использования схожих ресурсов (виды одного трофического уровня). В биоценозах же, кроме перечисленных факторов, отвечающих за видовое богатство, важнейшую роль играет длительность его существования. Сопряженная эволюция увеличивает число связанных трофических отношений, а также положительных, консортивных – мутуалистических и комменсальных. Поэтому чем древнее биоценоз, тем он богаче видами.
Список литературы
Азовский А.И. Таксономическое родство, морфологическое сходство и экологическая близость видов в сообществе морских псаммофильных инфузорий // Зоол. журн. 1990. Т. 69. No5. С. 5–16.
Андреев Н.Г. Луговедение. 1985. М.: Агропромиздат, изд. 2е., 255с.
Вальтер Г. Общая геоботаника 1982. М.: Мир, 264 с.
Вахрушев А.А., Раутиан А.С. Исторический подход к экологии сообществ // Журн. общ. биол. 1993. Т. 54. No 5. С. 532–553.
Гаузе Г.Ф. Экспериментальное исследование борьбы за существование между Paramecium caudatum, Paramecium aurelia и Stylonichia mytilus // Зоол. журн. 1934. Т. 13. No 1. С. 1–17.
Гаузе Г.Ф. Исследования над борьбой за существование в смешанных популяциях // Зоол. журн. 1935.Т. 14. No 2. С. 243–270.
Горчаковский П.Л. Лесные оазисы Казахского мелкосопочника. 1987. М.: Наука. 159 с.
Жигарев И.А. Изменение плотности населения мышевидных грызунов под влиянием рекреационного пресса на юге Подмосковья. // Зоол.журн. 1993б. Т. 72. Вып. 12. С. 117–137.
Жигарев И.А. Подходы к изучению животного населения в рекреационных лесных биоценозах // Научные чтения памяти профессора В.В. Станчинского. 2004. Изд#во СГПУ. Смоленск. С. 135–140.
Жигарев И.А. Закономерности рекреационных нарушений фитоценозов // Успехи совр. биологии. 1993а. Т. 113, вып.5. С. 564–575.
Жигарев И.А. Нарушения и видовое богатство сообществ. Эффект «высвобождения ресурсов нарушением». Научные труды Московского педагогического государственного университета. Физико-математические и естественные науки. Сборник статей. М.: Издательство «Прометей» МПГУ, 2007 С. 339–350.
Марков В.А. Экология и динамика численности лесных насекомых Центральной России: Монография. Ряз. гос. пед. ун-т. Рязань, 2004. 154 с.
Марфенина О.Е. Антропогенная экология почвенных грибов. 2005. М.: Медицина для всех, 196 с.
Попов И.Ю. Структура и динамика населения мелких млекопитающих в связи с сукцессиями растительности в Европейской южной тайге // Автореф. дис. … канд. биол. наук. 1998. М. 17 с.
Работнов Т.А. Фитоценология. 2е издание. 1983. М.: Изд-во МГУ. 296 с.
Шварц Е.А. Сохранение биоразнообразия: сообщества и экосистемы / М.: Т-во научных изданий КМК, 2004. 112 с.
Шефтель Б.И. Анализ пространственного распределения землероек в средней Енисейской тайге // Экологическая ординация и сообщества. 1990. М.: Наука, С. 15–32.
Connell J. H. Diversity in tropical rainforests and coral reefs // Science, 1978.N.Y. V. 199. P. 1302–1310.
Cody M.L. Coexistence, coevolution and convergent evolution in seabird communities // Ecology. 1973. V. 54. P. 31–43.
Elton Ch. S. The Ecology of Invasion by Animals and Plants. London, Methuen, 1958. 181 pp.
Elton C. Animal ecology. New#York. 1966. 207 p.
Den Boer P.J. Exclusion, competition or coexistence? A question of testing the right hypotheses // Zeitschrift für zoologische Sistematik und Evolutionforshung. 1985.V.23. N 4. P. 259–274.
Hutchinson G. E. Homage to Santa Rosalia, or why are there so many kinds of animals? // American Naturalist, 1959. V. 93. P. 145–159.
Hutchinson G. E. The paradox of the plankton // American Naturalist, 1961.V. 107. P. 406–425.
Park T. Experimental studies of interspecific competition. I. Competition between populations of the flour beetles Tribolium confusum Duval and Tribolium castaneum Herbst, Ecological Monographs, 1948. V. 18. P. 267–307.
Park T. Experimental studies of interspecific competition. II. Temperature, humidity and competition in two species of Tribolium. Physiological Zoology, 1954. V. 27. P. 177–238.
Simberloff D. Taxonomic diversity of island biotas // Evolution, 1970. V. 24. P. 23–47.
Simberloff D., Boecklen W. Patterns of extinction in the introduced Hawaiian avifauna: a re-examination of the role of the competition // Amer. Nat. 1991. V. 138. N 2. P. 300–327.
Рекомендуем прочесть
 Гуманизм, экология и рыночные отношения
Гуманизм, экология и рыночные отношения Уникальная экология человеческого хищничества
Уникальная экология человеческого хищничества “Макроэкология коронавирусной пандемии в Антропоцене”
“Макроэкология коронавирусной пандемии в Антропоцене” Человеческая деятельность формирует зимнюю экологию перелётной птицы, славки-черноголовки
Человеческая деятельность формирует зимнюю экологию перелётной птицы, славки-черноголовки Особенности архитектурно-градостроительного подхода к восстановлению биоразнообразия в урбанизированной среде в условиях юга Дальнего Востока
Особенности архитектурно-градостроительного подхода к восстановлению биоразнообразия в урбанизированной среде в условиях юга Дальнего Востока Последние птерозавры оказались не тем, чем казались
Последние птерозавры оказались не тем, чем казались Экспериментальная эволюция гуппи в речках Тринидада.
Экспериментальная эволюция гуппи в речках Тринидада. От ниш к нейтральности в биологическом сообществе
От ниш к нейтральности в биологическом сообществе
