
 Ивлев А.А., Воронин В.И.
Ивлев А.А., Воронин В.И.
Резюме публикатора. Анализ долговременных изменений соотношения тяжёлых и лёгких изотопов углерода в древесине прибайкальских лиственниц показывает временный и преходящий характер усиления фотосинтетической ассимиляции СО2 в ответ на рост его концентрации в атмосфере, созданный глобальным потеплением. C 90-х гг. XX в. оно прекращается: климатическая динамика, развивающаяся в связи с потеплением, делает июнь более жарким и намного более сухим месяцем, чем раньше. Это активизирует дыхание много сильней фотосинтеза, что отражается в утяжелении изотопного состава углерода отлагающейся древесины, и снижении продуктивности лесных участков. Показана повсеместность таких изменений в лесах разных районов Северной Евразии (в противоположность тундровым сообществам, в этот период ещё усиливавшим ассимиляцию — что, правда, и там завершилось к концу 2000-х гг.). Иными словами, по ходу развития глобального потепления в ХХ-ХХI вв. компенсаторная роль лесов в регуляции содержания СО2 в атмосфере до 1980-х гг. росла, а с 1990-х гг. резко снизилась.
Резюме авторское. В рамках предложенного ранее осцилляционного механизма фотосинтеза, согласно которому ассимиляция СО2 и фотодыхание представляют два реципрокных сопряженно осциллирующих процесса, контролируемых переключениями рибулозо-бисфосфат карбоксилазы/оксигеназы, дано объяснение связи между глобальным потеплением климата Земли, в значительной мере вызванным повышением концентрации СО2 в атмосфере, и изменением изотопного состава углерода растений.
Это объяснение находит подтверждение в изменении изотопного состава углерода годичных колец деревьев и свидетельствует о том, что до 90-х гг. XX в. обогащение легким изотопом 12С углерода годичных колец деревьев было результатом усиления фотосинтетической ассимиляции СО2. Последующее наблюдаемое резкое обогащение углерода годичных колец изотопом 13С, продолжающееся по настоящее время, свидетельствует о том, что произошла утеря компенсирующей роли фотосинтеза бореальных лесов для климата Земли.
***
Содержание
- 1 ***
- 2 Осциллирующий механизм функционирования фотосинтезирующей клетки. Связь с факторами внешней среды
- 3 Изотопное фракционирование углерода в биомассе фотосинтезирующей клетки как показатель ассимиляционной и фотодыхательной активности фотосинтезирующих организмов.
- 4 Материалы и методы
- 5 Результаты и обсуждение
Одну из главных угроз человечеству в XXI в. представляет глобальное потепление климата Земли, вызванное парниковым эффектом.
Многие связывают его с поступлением в атмосферу значительных количеств углекислого газа, образующегося при сжигания топлива. Согласно научному отчету, подготовленному большой группой ученых для межправительственной комиссии (IPCC, 2007), с постиндустриального уровня СО2 в среде, составлявшего 280 ррm [частей на миллион], концентрация СО2 выросла до 379 ррm в 2005 г. Причем только за последние 10 лет (1995-2005 гг.) прирост СО2 составил 1.9 ррm в год. По некоторым оценкам (Sellers et al, 1997) ежегодно в атмосферу Земли поступает до 7 млрд. т углерода. Из них 3 млрд. т остается в ней, 2 млрд. т абсорбируется океаном и 2 млрд. т, как полагают, связывается при фотосинтезе.
Осциллирующий механизм функционирования фотосинтезирующей клетки. Связь с факторами внешней среды
Чтобы понять механизм регуляции фотосинтезом содержания СО2 в атмосфере, необходимо рассмотреть как происходит сопряжение фотоассимиляции и фотодыхания на уровне самой клетки. В основе сопряжения незатухающие осцилляции углеродного метаболизма, состоящие в периодических переключениях функционирования клетки с ассимиляции СО2 на фотодыхание и обратно (Ivlev et al., 2004). Переключения осуществляются ферментом рибулозо-бисфосфат карбоксилазой/оксигеназой (РБФК/О), благодаря его способности работать как карбоксилаза и как оксигеназа в зависимости от соотношения в клетке концентраций О2/СО2 (Laing et al, 1974; McFadden, 1974). Согласно предложенному механизму (Ivlev et al., 2004), изменение соотношения концентраций О2/СО2 происходит за счет различия скоростей диффузии СО2 в клетку и из нее, с одной стороны, и скоростей реакций карбоксилирования рибулозобисфосфат карбоксилазы/оксигеназы (РиБФ) и декарбоксилирования глицина, — с другой. Первые медленнее, чем вторые. Обоснование этого гипотетического механизма дано в упомянутой работе, здесь же мы сошлемся на имеющиеся экспериментальные свидетельства существования осцилляций, не связанные с изотопными данными по углероду. Они следующие.
1. Обнаружение осцилляций концентраций СО2 в межклеточном пространстве листа табака при концентрациях близких к точке компенсации, т.е. в условиях, когда фотодыхание проявляется в наибольшей степени (Roussel et al., 2006);
2. Постиллюминационный всплеск выдыхаемого СО2 (post-illumination burst, PIB), свидетельствующий о разновременности ассимиляции СО2 и окисления части ассимилированного углерода при фотодыхании (Bulley, Tregunna, 1971; Atkin et al., 1998, 2000);
3. Обогащение выдыхаемого растением кислорода тяжелым изотопом 18О, обнаруженное еще в 1959 г. и подтвержденное последующими работами (Виноградов и др., 1960; Кутюрин, 1972). Оно, как показали авторы, является следствием исчерпывания кислорода, образовавшегося при фотосинтезе, на световое дыхание, сопровождающееся изотопным эффектом (эффектом Релея). Исчерпывание происходит потому, что кислород образуется порциями благодаря дискретному поступлению СО2 в клетку при ассимиляции (Ивлев, 1989).
4. Результаты исследования индукционных кривых замедленной люминесценции хлорофилла, свидетельствующие о влиянии фотодыхания на электрон-транспортную цепь (фотосистему II) (Satoh, Katoh, 1983). Как показал анализ индукционных кривых (Кукушкин, Солдатова, 1996), в спектре замедленной люминесценции хлорофилла должны быть два пика, отражающие влияние цикла Кальвина (ассимиляции СО2) и гликолатного цикла (фотодыхания) на электрон-транспортную цепь, разделенные интервалом в 1.6 с, что говорит о разновременности этих процессов.
Хотя длительность осцилляций не определена, по косвенным оценкам, полученным в упомянутых работах, она составляет от десятых долей до нескольких секунд.
Полагая гипотезу о фотосинтетических осцилляциях верной, периодические изменения концентраций СО2/O2, обеспечивающие переключения РБФК/О в клетке, и возникновение осцилляций могут быть описаны следующим образом (Ivlev et al., 2004).
В карбоксилазную фазу концентрация СО2 в клетке падает, поскольку скорость реакции ферментативного карбоксилирования РиБФ больше скорости диффузии СО2 в клетку. Однако из-за убывания субстрата различие в скоростях ферментативной реакции и диффузии снижается (рис.1, отрезок 1-2) и при некоторой концентрации СО2 (рис.1, точка 2) происходит переключение фермента на оксигеназную функцию. В оксигеназную фазу концентрация СО2 в клетке начинает расти (рис.1, отрезок 1-3), так как скорость глициндекарбоксилазной реакции в клетке суммируется со скоростью поступления СО2 в клетку из среды. Затем рост СО2 замедляется из-за снижения скорости ферментативной реакции вследствие исчерпывания субстратов РиБФ и О2. После того как концентрация СО2 в клетке становится выше, чем в среде, начинается собственно дыхание — выброс углекислого газа из клетки (рис.1, отрезок 3-4), происходящий за счет диффузии. При этом рост концентрации СО2 в клетке еще более замедляется, так как глициндекарбоксилазная реакция и диффузия действуют на концентрацию СО2 в противоположных направлениях. По достижении некоторого максимального значения концентрации СО2 (рис.1, точка 4), определяемого внешними факторами, происходит переключение фермента на карбоксилазную функцию. Концентрация СО2 в клетке вследствие меньшей скорости диффузии по сравнению со скоростью РиБФ-карбоксилазной реакции начинает падать (рис.1, отрезок 4-1′). Далее все процессы периодически повторяются.
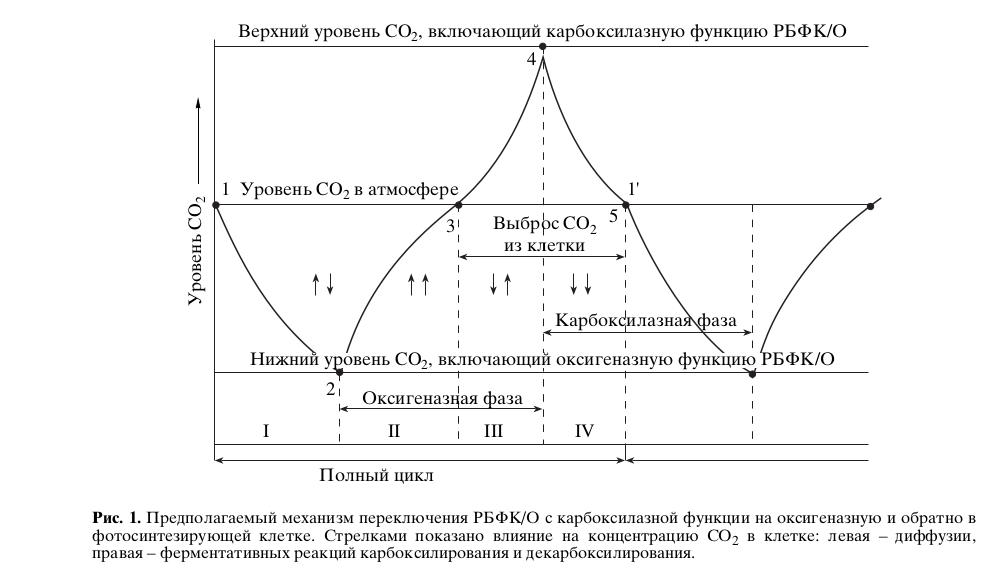 В аспекте данной работы наиболее интересен вопрос, как контролируются точки 2 и 4 триггерного переключения фермента (рис.1) с карбоксилазной функции на оксигеназную. Эти точки определяют протяженность карбоксилазной и оксигеназной фаз. Мы полагаем, что длительность фаз контролируется внешними факторами, которые в конкретных условиях среды являются критичными для метаболизма (лимитирующими скорость процесса). В благоприятных для организма условиях больший вклад дает ассимиляционная составляющая, контролируемая длительностью карбоксилазной фазы, а в неблагоприятных стрессовых условиях большую роль начинает играть фотодыхательная составляющая, контролируемая длительностью оксигеназной фазы. На роль дыхания в формировании адаптационного синдрома указано в работе Усманова с соавт. (2001).
В аспекте данной работы наиболее интересен вопрос, как контролируются точки 2 и 4 триггерного переключения фермента (рис.1) с карбоксилазной функции на оксигеназную. Эти точки определяют протяженность карбоксилазной и оксигеназной фаз. Мы полагаем, что длительность фаз контролируется внешними факторами, которые в конкретных условиях среды являются критичными для метаболизма (лимитирующими скорость процесса). В благоприятных для организма условиях больший вклад дает ассимиляционная составляющая, контролируемая длительностью карбоксилазной фазы, а в неблагоприятных стрессовых условиях большую роль начинает играть фотодыхательная составляющая, контролируемая длительностью оксигеназной фазы. На роль дыхания в формировании адаптационного синдрома указано в работе Усманова с соавт. (2001).
Среди факторов, определяющих эволюцию фотосинтезирующей жизни и земной атмосферы, основная роль отводится отношению концентраций СО2 и О2 в среде и температуре (Igamberdiev, Lea, 2005). При исследовании углекислотного газообмена хвойных в Предбайкалье показано, что температурный стресс увеличивает дыхание хвои у сосны в 7 раз, а у лиственницы в 3 раза (Суворова и др., 2005). В конечном счете, влияние внешних факторов проявляется в изотопном составе углерода биомассы и, в частности, в изотопном составе углерода годичных колец.
Изотопное фракционирование углерода в биомассе фотосинтезирующей клетки как показатель ассимиляционной и фотодыхательной активности фотосинтезирующих организмов.
Для последующего анализа принципиально важно то, что каждый из названных процессов сопровождается фракционированием изотопов углерода с эффектами противоположными по знаку (Ивлев, 1993). При ассимиляции СО2 происходит обогащение углерода биомассы изотопом 12С. Фотодыхание обогащает биомассу тяжелым изотопом 13С.
Таким образом, противоположность знаков изотопных эффектов порождает в фотосинтезирующей клетке два изотопноразличающихся углеродных потока: поток углеродных субстратов, связанных с углеводами, образующимися при ассимиляции, и поток углеродных субстратов, связанных с углеводами, образуемыми при фотодыхании. Вклад этих потоков в синтезируемую биомассу зависит от интенсивности фотоассимиляции и фотодыхания (Ивлев, 2004). Это позволяет использовать изотопный состав углерода биомассы фотосинтезирующих организмов синтезирующих организмов как способ оценки интенсивности указанных процессов, а следовательно, как способ изучения регуляции посредством фотосинтеза содержания СО2 в атмосфере. Этот способ был использован нами при изучении средорегулирующей способности деревьев, а в качестве объекта исследования был выбран изотопный состав углерода биомассы годичных колец лиственницы. С точки зрения фракционирования изотопов важным является то, что лиственница, по сравнению с елью и сосной, отличается высоким уровнем как фотосинтетической активности, так и дыхания на свету (Щербатюк и др., 1991).
Материалы и методы
Керны древесины лиственницы сибирской Larix sibirica Ledeb. для изотопного анализа отобрали с пяти деревьев (по пять кернов с одного участка ствола дерева) в 70 км северо-западнее г. Иркутска (52°14′ с.ш., 104° 11′ в.д.) в сосново-лиственничном разнотравном лесу (Воронин, 2005). Среднегодовое количество осадков последние 25 лет составляет 470 мм. а среднегодовая температура воздуха 0.8°С (данные Иркутской гидрометеорологической обсерваторией (ГМО)).
Под микроскопом буровые керны древесины препарировали на годичные слои. Для каждого года концентрировали пробы древесины по пяти кернам, в результате чего достигалась необходимая масса навески и получалась средняя проба для отдельного дерева. Натронной варкой полученной древесины выделяли целлюлозу, которую и использовали в изотопном анализе.
Определение изотопного состава древесины для временного периода 1682-1998 гг. проводили в Институте химии и динамики геосферы (Юлих, Германия) на изотопном масс-спектрометре IRMS (Optima) (Voronin et al., 2001). В 2006 г. были дополнительно отобраны керны деревьев с тех же самых деревьев и получены изотопные данные для периода 1999-2005 гг. Изотопный анализ выполнен в Центре по исследованию окружающей среды (Лейпциг-Халле, Германия) на масс- спектрометре Delta S (Finnigan). Точность определения составляет 0.02‰ (Voronin et al. 2001). Протяженность изотопной хронологии, полученной по пяти деревьям, составляет 323 года (1682-2005 гг.). Среднее значение δ13С целлюлозы годичных колец лиственницы равно -22.58‰.
Изотопные данные приводятся в общепринятых единицах δ13С, представляющих нормированную разность отношений изотопных концентраций 13С/12С в пробе растительного материала (Rp) и стандарте (Rref): δ13Сp (‰) = (Rp — Rref)/Rref, х 103 (Craig, 1954). В качестве стандарта Rref использовали международный стандарт РDВ, в котором отношение (13С/12С)ref равно 1123 х 10-3 (Craig, 1957).
Расчет и анализ функции отклика изотопных значений на действие основных климатических факторов выполнены в программных пакетах DPL-99 (программа RESPO) (Holmes, 1998) и Statistica v.5.5.
Результаты и обсуждение
Изотопный состав углерода годичных колец деревьев как показатель способности фотосинтеза компенсировать парниковый эффект углерода
В отличие от многих других фотосинтезирующих организмов, например, от океанического планктона, изотопный состав углерода биомассы которого отражает сиюминутные условия фотосинтеза, изотопный состав углерода годичных колец у многолетних растений позволяет проследить влияние внешних факторов на большом временном отрезке.
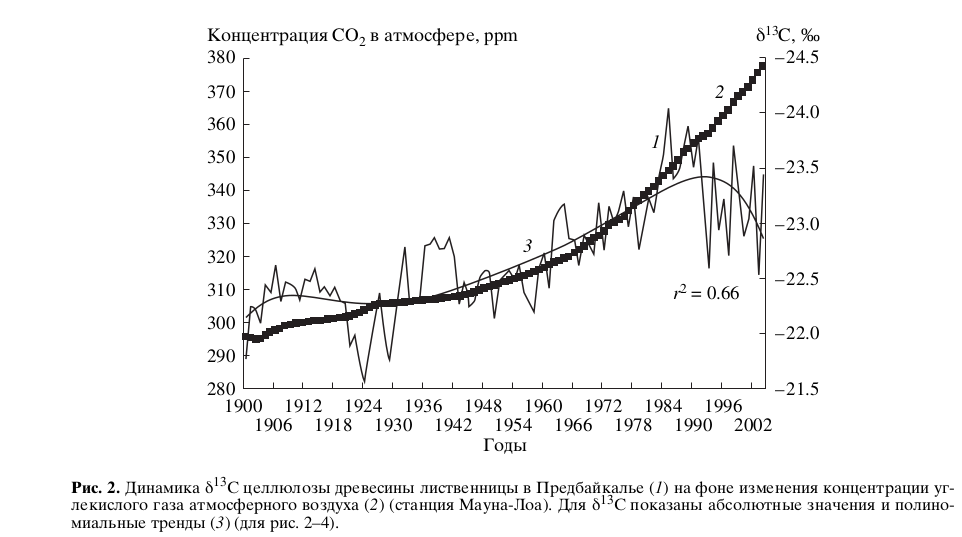
В настоящей работе мы оперируем изотопными данными. охватывающими временной отрезок с начала прошлого века до наших дней, поскольку этот период обеспечен надежной метеорологической информацией (Воронин и др., 2003а, 20036; Воронин, 2005). Из рис.2 видно, что с середины прошлого столетия наблюдается монотонное обогащение целлюлозы деревьев изотопом 12С, которое в 90-х г. XX в. сменяется столь же устойчивым. но гораздо более динамичным обогащением ее тяжелым изотопом 13С. В рамках рассмотренного механизма такое поведение изотопного состава углерода годичных колец до начала 90-х гг. XX в. можно объяснить усилением ассимиляционной активности деревьев, что приводит к обогащению биомассы легким изотопом. Этот процесс был закономерной реакцией деревьев на рост в атмосфере концентрации антропогенной СО2, образующейся, в первую очередь, за счет сжигания органического топлива, а также при лесных пожарах, и в силу этого имеющей более легкий изотопный состав углерода, чем атмосферная СО2(-20‰…-30‰ против -7‰).
Изотопное утяжеление, которое наблюдается после 90-х гг. XX в., мы объясняем тем. что доминирующая вначале ассимиляционная составляющая фотосинтеза уменьшилась, а фотодыхательная усилилась. Это подтверждается неуклонным снижением сезонной продуктивности хвойных пород (лиственницы и сосны) в Предбайкалье. что зафиксировано в ходе многолетних измерений ассимиляции СО2, с помощью ИК-газоанализатора Infralit, ГДР (Суворова и др., 2005).
 В свою очередь, смена тренда изотопного состава углерода годичных колец может означать смену внешнего фактора, контролирующего переключения функций РБФК/О. Можно допустить. что в системе фотосинтеза появился новый управляющий фактор. Им, как мы полагаем, стала температура воздуха, что привело к увеличению длительности оксигеназной фазы (усилению вклада «тяжелых» продуктов фотодыхания в синтез биомассы). В пользу такого допущения свидетельствует сопоставление значений изотопного состава углерода годичных колец лиственницы из Предбайкалья с динамикой июньской температуры в исследуемом регионе (рис.3). Антибатный характер изменений сопоставляемых характеристик с начала 1990-х гг. меняется на симбатный.
В свою очередь, смена тренда изотопного состава углерода годичных колец может означать смену внешнего фактора, контролирующего переключения функций РБФК/О. Можно допустить. что в системе фотосинтеза появился новый управляющий фактор. Им, как мы полагаем, стала температура воздуха, что привело к увеличению длительности оксигеназной фазы (усилению вклада «тяжелых» продуктов фотодыхания в синтез биомассы). В пользу такого допущения свидетельствует сопоставление значений изотопного состава углерода годичных колец лиственницы из Предбайкалья с динамикой июньской температуры в исследуемом регионе (рис.3). Антибатный характер изменений сопоставляемых характеристик с начала 1990-х гг. меняется на симбатный.
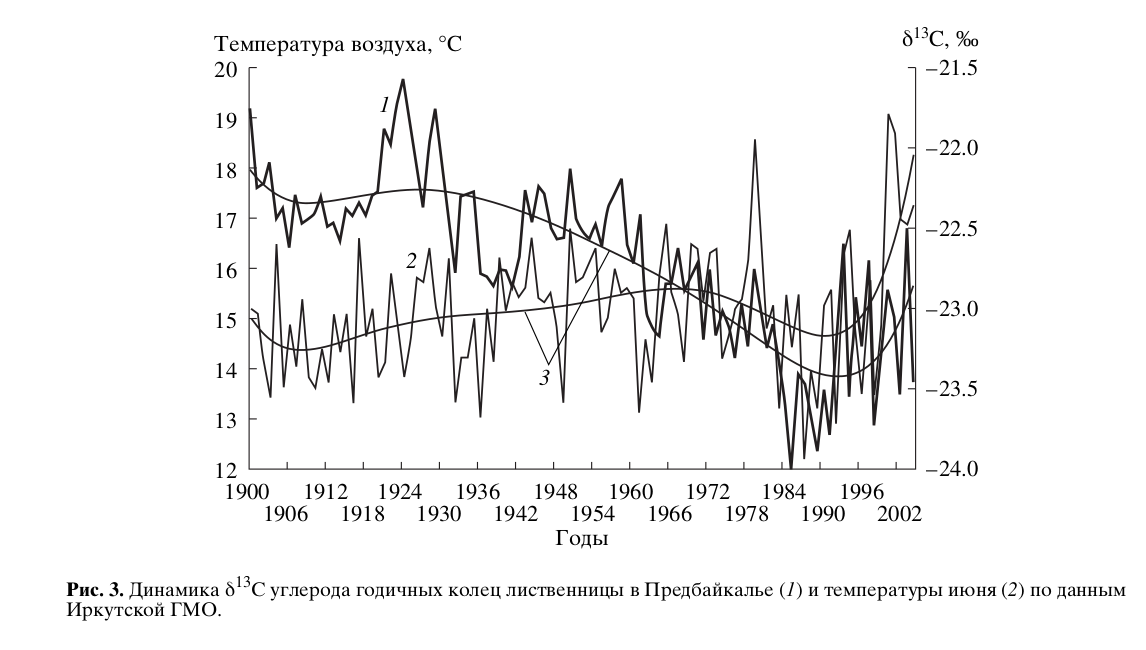 Акцент на раннелетней температуре сделан с учетом результатов расчета функций отклика концентрации изотопов 13С/12С в целлюлозе лиственницы на месячные значения метеопараметров за период 1980-2005 гг. Статистически значимыми для динамики δ13С оказываются только июньские значения температуры воздуха и суммы осадков. Причем связь с изотопными данными для температуры положительна, а для осадков — отрицательна. Баланс этих двух факторов и отражает влияние среды на динамику δ13С целлюлозы лиственницы. Как видно из рис.3, в последние годы произошел существенный рост температуры июня, и тренд их динамики имеет экспоненциальный характер. Тренд же июньских осадков остается стабильным на протяжении последнего века и текущих лет и аппроксимируется линейной функцией (рис.4).
Акцент на раннелетней температуре сделан с учетом результатов расчета функций отклика концентрации изотопов 13С/12С в целлюлозе лиственницы на месячные значения метеопараметров за период 1980-2005 гг. Статистически значимыми для динамики δ13С оказываются только июньские значения температуры воздуха и суммы осадков. Причем связь с изотопными данными для температуры положительна, а для осадков — отрицательна. Баланс этих двух факторов и отражает влияние среды на динамику δ13С целлюлозы лиственницы. Как видно из рис.3, в последние годы произошел существенный рост температуры июня, и тренд их динамики имеет экспоненциальный характер. Тренд же июньских осадков остается стабильным на протяжении последнего века и текущих лет и аппроксимируется линейной функцией (рис.4).
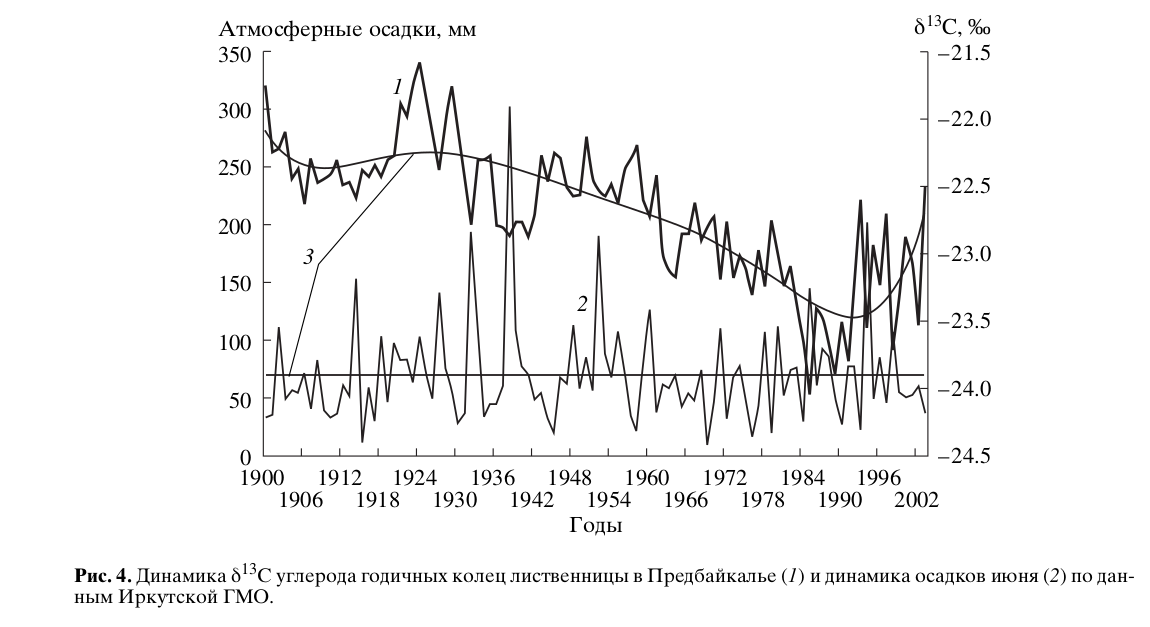 Чтобы проследить, как меняется характер связи δ13С с этими факторами, сравним коэффициенты корреляции за периоды 1900-1979 гг. и 1980-2005 гг. Для суммы осадков июня они будут равны -0.43 и -0.12, а для температуры воздуха значения составляют 0.41 и -0.13 соответственно. Из сравнения видно, что характер связи δ13С с суммой осадков этого месяца не претерпел кардинальных изменений — изменились только абсолютные значения, чего нельзя сказать в отношении температуры воздуха июня. Произошла смена знака связи и из статистически незначимого параметра она перешла в ранг управляющего фактора. В период 1900-1979 гг. эту роль играла температура июля. Атмосферное увлажнение июля в среднем на 30% больше, чем июня, и в результате баланса между температурой и осадками не наблюдалось заметного утяжеления целлюлозы изотопом δ13С. Вследствие повышения июньской температуры в период 1980-2005 гг. на 0.3°С (против 0.1°С в июле) и в отсутствие адекватного повышения нормы осадков, произошло нарушение этого баланса. Тренд динамики δ13С целлюлозы лиственницы стал повторять тренд температуры июня и именно она стала определять направление всего процесса изотопного фракционирования (рис.3). Осадки же по прежнему детерминируют только динамику абсолютных значений δ13С.
Чтобы проследить, как меняется характер связи δ13С с этими факторами, сравним коэффициенты корреляции за периоды 1900-1979 гг. и 1980-2005 гг. Для суммы осадков июня они будут равны -0.43 и -0.12, а для температуры воздуха значения составляют 0.41 и -0.13 соответственно. Из сравнения видно, что характер связи δ13С с суммой осадков этого месяца не претерпел кардинальных изменений — изменились только абсолютные значения, чего нельзя сказать в отношении температуры воздуха июня. Произошла смена знака связи и из статистически незначимого параметра она перешла в ранг управляющего фактора. В период 1900-1979 гг. эту роль играла температура июля. Атмосферное увлажнение июля в среднем на 30% больше, чем июня, и в результате баланса между температурой и осадками не наблюдалось заметного утяжеления целлюлозы изотопом δ13С. Вследствие повышения июньской температуры в период 1980-2005 гг. на 0.3°С (против 0.1°С в июле) и в отсутствие адекватного повышения нормы осадков, произошло нарушение этого баланса. Тренд динамики δ13С целлюлозы лиственницы стал повторять тренд температуры июня и именно она стала определять направление всего процесса изотопного фракционирования (рис.3). Осадки же по прежнему детерминируют только динамику абсолютных значений δ13С.
Точно такой же характер связи установлен для δ13С целлюлозы лиственницы и температуры июля в северных лесах Якутии (Николаев и др., 2006) и в высокогорном районе Северного Байкала (Воронин, 2005). Поэтому можно считать, что именно температура стала фактором, определяющим характер изотопных изменений.
В работах, где интерпретация изотопных эффектов опирается на традиционную стационарную модель фотосинтеза (Farquhar et al., 1982), существенная роль отводится устьичиой проводимости и распределению устьиц на листе (Condon et al., 1992; Рicon et al., 1996; Scheidegger et al., 2000) как фактору, определяющему изотопный состав углерода биомассы. Устьичная проводимость, действительно, связана с изменением изотопного состава ассимилируемого углерода. С позиции осцилляционной модели, оно трактуются следующим образом (Ivlev, 2003): чем меньше СО2 попадает в клетку, тем больше степень исчерпывания попадающего в клетку углерода и тем “тяжелее» оказывается изотопный состав ассимилятов. Но эта связь — опосредованная связь двух величин, зависящих от третьего фактора. Таким фактором является доступность влаги (количество осадков), которая, в свою очередь, завиcит от температуры. Выше мы показали, что именно температура становится с 90-х гг. XX в. лимитирующим скорость процесса фактором.
Таким образом, до начала 90-х гг. XX в. в природе действовал механизм компенсации повышения концентрации СO2 в среде за счет усиления фотосинтстичсской ассимиляции. Он отражал известный в термодинамике принцип Ле-Шателье. Увеличение концентрации СО2 усиливало карбоксилазиую активность РБФК/О. Последняя, усиливая ассимиляцию, снижала концентрацию. Но с 90-х гг. XX в. этот механизм перестал работать. Проведенные исследования показали смену фактора, управляющего фотосинтезом деревьев, происшедшую в начале 90-х годов XX века. Это выразилось в утяжелении изотопного состава углерода древесины почти одновременно в разных районах Северной Евразии: в Монголии, Байкальском регионе, Якутии и в лесах циркумполярной зоны (Воронин и др., 20036; Saurer et al., 2004; Николаев и др., 2006). Другими словами, компенсаторная роль лесов в регуляции содержания СО2 в атмосфере резко снизилась.
Это подтверждается данными спутниковых оценок фотосинтетической продуктивности растительного покрова циркумполярной области Северного полушария (севернее широты 50°) за последние 22 года (Bunn, Goetz, 2006; IPCC…, 2007). Результаты анализа показывают: если тундровая растительность позитивно откликнулась на потепление, и интенсивность фотосинтеза растений там возросла, то северные леса, напротив, обнаруживают тенденцию к снижению интенсивности фотосинтеза (за счет усиления дыхания). Причем, в мае-июне фотосинтез деревьев усиливается, а в июле-августе снижается.
Ранее для всех районов субарктического пояса Сибири была выявлена следующая тенденция: скорость нарастания стволовой фитомассы хвойных деревьев, линейно зависящая от ассимиляции СO2, замедляется после 1960-х гг., по сравнению с ростом температуры Северного Полушария (Briffa et al., 1998).
Авторы благодарны Т. Воеttger (Лейпциг-Халле, Германия) за помощь в определении изотопного состава годичных колец, Г.Г. Суворовой и Л.С.Яньковой (СИФИБР СО РАН) за предоставленные материалы по фотосинтезу.
Источник Известия РАН. Серия биологическая. 2007. №6.
Рекомендуем прочесть
 Геоинженерия — шанс для климата Земли или опасные фантазии?
Геоинженерия — шанс для климата Земли или опасные фантазии? Как соотносятся прямое приспособление и эволюционные изменения?
Как соотносятся прямое приспособление и эволюционные изменения? Вибрационные сигналы в коммуникации животных
Вибрационные сигналы в коммуникации животных Отбор адаптивной и неадаптивной пластичности в экспрессии генов при приспособлении анолисов к городскому «острову тепла»
Отбор адаптивной и неадаптивной пластичности в экспрессии генов при приспособлении анолисов к городскому «острову тепла» Восприятие шумов
Восприятие шумов Человеческая деятельность формирует зимнюю экологию перелётной птицы, славки-черноголовки
Человеческая деятельность формирует зимнюю экологию перелётной птицы, славки-черноголовки Пение самок воробьиных птиц: исключение или закономерность?
Пение самок воробьиных птиц: исключение или закономерность? Расшифровка сигналов агрессии у коростелей
Расшифровка сигналов агрессии у коростелей
