

Anolis allisoni
Резюме. В городах неизменно жарче, чем за их пределами, из-за образования «острова тепла». Для эктотермных существ, вроде ящериц с насекомыми, это большой стресс. Неудивительно, что у них отмечается рост термостойкости, скажем у городских муравьёв. Т.е. способность справляться с повышенной температурой (устойчиво действовать, пользоваться присадами, убежищами и пр.) важный аспект адаптации к урбосреде. Приводимые ниже исследования гребенчатых анолисов на Пуэрто-Рико показали, что для анолисов поведенческая адаптация к «островам тепла» недостаточна, при освоении урбосреды идут изменения генов с физиологией под действием отбора. Верно и наоборот: виды карибских анолисов, приспособленные к большей жаре и сухости в их естественных местообитаниях, которые деятельны при более высоких температурах, легче осваивают урбосреду в городах разных островов этого моря. Об этом — три представляемых материала.
Большая термостойкость облегчает урбанизацию у анолисов
В городах неизменно жарче, чем за их пределами, из-за образования «острова тепла». Для эктотермных существ, вроде ящериц с насекомыми, это большой стресс. Неудивительно, что у них отмечается рост термостойкости, скажем у городских муравьёв. Т.е. способность справляться с повышенной температурой (устойчиво действовать, пользоваться присадами, убежищами и пр.) важный аспект адаптации к урбосреде.
Приводимые ниже исследования гребенчатых анолисов на Пуэрто-Рико показали, что для анолисов поведенческая адаптация к «островам тепла» недостаточна, при освоении урбосреды идут генов под действием отбора.
Верно и наоборот: виды карибских анолисов, приспособленные к большей жаре и сухости в их естественных местообитаниях, которые деятельны при более высоких температурах, легче осваивают урбосреду в городах разных островов этого моря (Winchell et al. 2020), см. рис.1: красный цвет — самые толерантные, зелёный — самые нетерпимые, синий — промежуточные. Исследование геномов кубинских анолисов выявило гены, связанные с термочувствительностью к теплу (Akashi et al., 2016); на Кубе богаче всего факуна анолисов, шире разнообразие их жизненных форм, и температурных условий природных местообитаний данных видов.
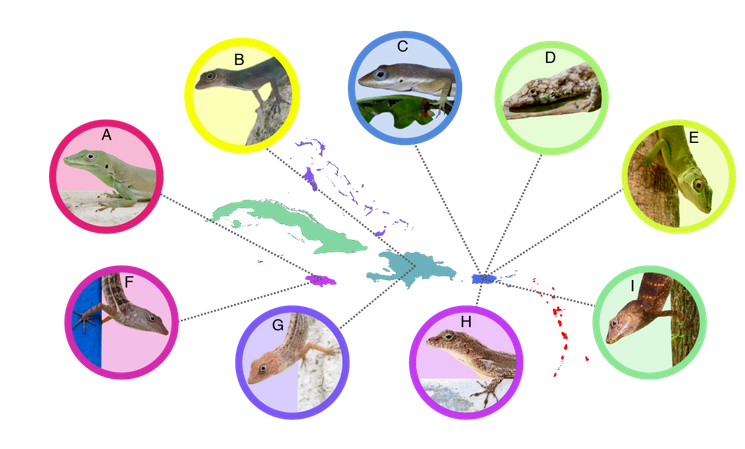
Рис.1. Анолисы разных карибских о-вов, изученные Кристин Винчелл с соавт. (2020).
На этой основе Shunsuke Kanamori с соавт. (2021) предположили, не может быть большая термостойкость в исходных биотопах стать эффективной преадаптацией к городским «островам тепла»? Они проанализировали транскриптом 9 кубинских анолисов (природные популяции!), отыскивая в тём признаки отбора на рост термостойкости в городах, так называемые «геномные подписи», сравнением его у видов, процветающих с городах, с видами, урбанизирующимися медленно и плохо. Для этого рассчитывали по Fst для разных участков генетического и белкового «текста» и искали значимые выбросы, по которым определяли гены, бывшие под отбором в ветвях, ведущих к видам с самыми жаркими и сухими природными биотопами.
Жарче и суше всего в природных местообитаниях трёх видов из 9, относящихся к двум неродственным (!!) филогенетическим ветвям — A.porcatus, A.allisoni и A.sagrei. Они же и самые урбанизированные — освоили Гавану, Майами и др. города Флориды (вне основного ареала, здесь это закрепившиеся инвазивные виды). Пять других видов (A. alutaceus, A. isolepis, A.garrido, A.allogus и A.mystery, ) наоборот, населяют сомкнутые леса, сильно более прохладные и затенённые. Наконец, A.homolechis живёт на тенистых участках лесных опушек.
Авторы исследовали 5962 гена; под отбором в ветвях, включающих урбанофильные виды (A.porcatus, A.allisoni и A. sagrei), из них были 21. Правда, это разных гены в двух ведущих к ним кладах, общих не было. Т.е. каждая филогенетическая ветвь нашла свой собственный путь приспособления к жаре и сухости в «своих» биотопах.

Anolis sagrei

Anolis porcatus
Инвентаризация урботолерантных видов
Содержание
Дальше Кристин Винчелл с соавт. (2020) анализировала разные виды карибских анолисов (некоторые из них см. рис.1, но всего их более 100) на предмет урботолерантности и термостойкости, оцененным по собственным исследованиям и из литературы, накладывая эти данные на филогенетическое дерево, составленное методами ДНК-генеалогии (рис.2). Самым информативным был справочник Henderson & Powell’s (2009) book по естественной истории амфибий и рептилий, с крайне детальным описанием естественной истории и ключевых характеристики разных анолисов. Однако здесь оставалось много пробелов, которые закрывали, используя данные гражданской науки, фото соответствующих видов в их среде обитания, городской и/или природной, в сети iNaturalist, в предположении что люди в целом фотографируют ящериц там, где они (те и другие) живут, с частотой, пропорциональной встречаемости разных видов в разных ландшафтах. Степень урбанизации территорий, где ящериц фотографировали, оценивали по спутниковым снимкам соответствующих территорий. Дальше использовали баллы урботолерантности: их добавляли, если вид чаще фотографировали на зданиях; не встречающиеся там виды, сфотканные лишь на природных территориях, получали баллы за нетерпимость.

Дальше подряд пять избегающих урбанизации видов. A.homolechis
A.alutaceus

A.mestrei

A.isolepis
Филогегенетический анализ урботолерантности у анолисов
Вероятность предпочтения или избегания урболандшафта у видов с разной балльной оценкой рассчитывали, исходя из объёма выборки наблюдений, достаточных для проявления данного предпочтения. Затем по филогенетическим алгоритмам реконструировали предковые уровни термостойкости для разных клад 131 вида карибских анолисов (рис.2). Конечно, это не реконструкция освоения ящерицами урбосреды, анолисы намного старше неё! Это попытка понять эволюцию поведенческих, физиологических, экологических и морфологических признаков, влияющих на первую реакцию данных видов, когда он в своих биотопа «столкнётся» с урбанизацией: будет он сразу использовать города или отступит? Оказывается, термостойкость это предсказывает.

Рис.2. Филогенетическое древо анолисов разных островов. Внизу градиент термостойкости, от максимальной (красное слева, самые обычные в урбосреде) к минимальной (жёлтый справа, городов избегают) через промежуточную (синий в центре, без выраженного предпочтения или избегания).
Ключевые характеристики урботолерантных видов
Прежде всего, урботолерантность широко распространена на Карибах и демонстрирует сильный филогенетический сигнал, по которому можно прогнозировать её уровень у малоизученных видов или ещё не контактирующих с урбосредой. Важно подчеркнуть, что данный подход нельзя использовать для прогноза устойчивости видов к антропогенной потере их природных биотопов, но лишь для поиска видов-удобных модельных объектов для городской экологии и роли городов как «арен» микроэволюции.
Далее авторы рассчитывали корреляции баллов урботолерантности/нетерпимости с разными экологическими и морфологическими характеристиками. У более толерантных видов выше температура тела, меньше брюшных чешуек, больше пластинок сзади (важно для удержания на гладких поверхностях), короче задние ноги, природные биотопы теплее и суше. Всё это может быть преадаптацией к быстрому заселению города и использования преимущества антропогенных ниш (например, на зданиях и вокруг них).
Авторов удивило, что ни один вид не подходит идеально для урбосреды (т.е. для всех она экстремальна, без выработки приспособлений отблром не обойтись). Сперва они думали, что наземные виды более урботолерантны, но многие из них урбанизацию не переносят урбанизацию, и много видов других жизненных форм, более чем терпимых к ней (например, A.equestris или A. distichus)!
Источник Anole Annals

Anolis lineatopus, один из многих урботолерантных видов
Отбор «городских» гребенчатых анолисов на жароустойчивость
Осваивая разные города о.Пуэрто-Рико, гребенчатые анолисы Anolis cristatellus (рис.7А) сделались более термостойкими, что адаптивно в условиях городского «острова тепла», нагретого асфальта, металлических заграждений и бетонных стен. У «городских» ящериц значимо выше максимальные температуры, при которых они ещё могут двигаться и охотиться. Выше и переносимая температура тела, что значительно расширяет набор доступных присад и поверхностей. Критический максимум температуры для «городских» анолисов в каждом из 4-х городов острова (Mayagüez, Aguadilla, Arecibo, San Juan) в среднем на 0,870 С выше, чем для «лесной» популяции, из которой возникла данная городская (рис.7Б).
Разные города освоены независимо: в г.Mayagüez ящерицы активны при максимальной температуре 400C1. Только у ящериц, выросших в городе, рост термостойкости сопряжён также с большей способностью к поведенческой регуляции температуры тела, что расширяет диапазон благоприятных температур (Campbell-Staton et al., 2020, fig.1).
А.

А.cristatellus

Рис.7. Гребенчатый анолис и генетические изменения ящериц разных городов при урбанизации. А. Сама ящерица. Б. Независимый рост термостойкости в разных городах. Попарное сравнение генофондов «городских» популяций с лесными (серые vs чёрные кружки) по Fst между разными участками генома показывает независимое происхождение первых от вторых в каждом городе (d). У «городских» ящериц выше предельная температура активного действия (а), возможности пользоваться присадами (b) и переносимая температура тела (с), чем в соответствующих «лесных» популяциях. У первых также выше термостойкость (Стmax), чем у вторых (полые и заполненные кружки). У них же сильней повышается температура тела сравнительно с температурой среды, показывая недостаточность одной поведенческой терморегуляции для успешного приспособления (нужен отбор генных вариантов, обеспечивающих большую термостойкость). Источник: Campbell-Staton et al., 2020.
«Городские» ящерицы генетически дивергировали от «лесных» независимо в каждом городе острова. Степень дивергенции тем выше, чем крупнее освоенный город, максимум — в г.Сан-Хуан (рис.7Б).
Определяя полиморфизмы по всему геному, авторы искали локусы с максимумом различий между «городскими» vs «лесными» популяциями, оцениваемых Fst. Их больше всего найдено в гене RARS1 первой хромосомы, кодирующем аргинил т-РНК-синтетазу, обеспечивающую синтез белка; на нематодах показана его связь с адаптацией к абиотическому стрессу (Anderson et al., 2009, цит. по Campbell-Staton et al., 2020). Поэтому он вовлекается в развитие термоустойчивости, идущей независимо во всех 4-х городах и достигнутой там в разной степени (рис.8).
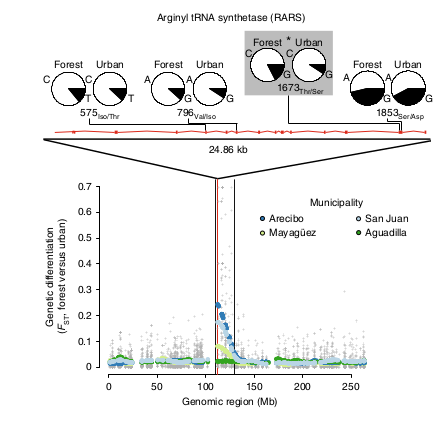 Рис.8. В хромосоме 1 найден участок с гораздо большим уровнем дивергенции «городских» популяций от «лесных», чем в среднем по геному. Ось Х — участки генетического «текста», мегабайтные интервалы, серые крестики — базовый уровень FST для SNP по всей хромосоме; цветные кружки — FST для 5-мегабайтных интервалов генома в разных парах сопоставляемых популяций: Маягуэз (светло-зеленый), Сан-Хуан (светло-голубой), Агуадилья (темно-зеленый) и Аресибо (темно-синий). Круговые диаграммы показывают частоту основных (белый сектор) и второстепенных (черный сектор) аллелей в несинонимичных заменах, приходящихся участки гена аргинил-тРНК-синтетазы, различные у «городских» и «лесных» популяций. Самый сильный сдвиг частот (8,4%) замена Thr на Ser в позиции 1673 (обозначена * и заштрихована). Вертикальный пунктир — уровень генетической дивергенции.
Рис.8. В хромосоме 1 найден участок с гораздо большим уровнем дивергенции «городских» популяций от «лесных», чем в среднем по геному. Ось Х — участки генетического «текста», мегабайтные интервалы, серые крестики — базовый уровень FST для SNP по всей хромосоме; цветные кружки — FST для 5-мегабайтных интервалов генома в разных парах сопоставляемых популяций: Маягуэз (светло-зеленый), Сан-Хуан (светло-голубой), Агуадилья (темно-зеленый) и Аресибо (темно-синий). Круговые диаграммы показывают частоту основных (белый сектор) и второстепенных (черный сектор) аллелей в несинонимичных заменах, приходящихся участки гена аргинил-тРНК-синтетазы, различные у «городских» и «лесных» популяций. Самый сильный сдвиг частот (8,4%) замена Thr на Ser в позиции 1673 (обозначена * и заштрихована). Вертикальный пунктир — уровень генетической дивергенции.
Источник: Campbell-Staton et al., 2020.
Урбанизация ведёт к накоплению 4-х несинонимичных замен в этом гене в позициях 1673, 1853, 595, 796. Первая из них функционально самая значимая: среди «городских» анолисов на 21,6% повышена доля особей, имеющих там С-С (это позиция 558 в белке, треонин заменён на серин), и понижена на 20,4% — С-G. Только этот полиморфизм показывал значимые различия между популяциями во всех 4-х городах; именно у «городских» анолисов гомозиготы С-С более термоустойчивы, за счёт лучшей акклимации в ответ на тепловой стресс (рис.9).
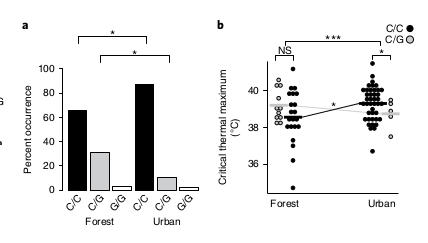 Рис.9. Локус наибольшего вклада в термостойкость «городских» анолисов, позиция 1673 в RAS1. a. Особи с генотипом С/С в этой позиции перепредставлены в «городских» популяциях сравнительно с «лесными», С/G — недопредставлены, различия высоко значимы. b. Особи С/С из «лесных» популяций демонстрируют большее изменение Ctmax при выращивании в условиях «городских» температур, и больший Сmax в этом случае, чем у ящериц генотипа С/G, т. е. обладают повышенной термопластичностью.
Рис.9. Локус наибольшего вклада в термостойкость «городских» анолисов, позиция 1673 в RAS1. a. Особи с генотипом С/С в этой позиции перепредставлены в «городских» популяциях сравнительно с «лесными», С/G — недопредставлены, различия высоко значимы. b. Особи С/С из «лесных» популяций демонстрируют большее изменение Ctmax при выращивании в условиях «городских» температур, и больший Сmax в этом случае, чем у ящериц генотипа С/G, т. е. обладают повышенной термопластичностью.
Источник: Campbell-Staton et al., 2020, fig.2-3
Параллельные сдвиги частот полиморфизма во всех 4-х городах говорят об их обусловленности положительным отбором. Что подтверждается анализом транскриптомов скелетных мышц ящериц в нормальных условиях и под действием теплового стресса (320 С и 250 С). В «лесных» популяциях в этом случае различалась экспрессия 818 генов, в «городских» — 421, из них общих 286. Среди них выделили гены, предположительно связанные с термотолерантностью (у них значительней разница в экспрессии между «городскими» и «лесными» популяциями, чем у остального генома). Далее авторы из кладок «лесных» и «городских» анолисов в стандартных условиях искусственно вырастили потомство (n=7 и 9). Гены, характеризующиеся разницей в экспрессии между популяциями, также показывали большую дисперсию экспрессии при 320 С, чем при 250 С, создавая возможность дальнейшего роста термоустойчивости под действием тех же селективных давлений (Campbell-Staton et al., 2020, fig.4).
Примечания
1Длина самой ящерки без хвоста ~5 см. Термостойкость оценивали, укладывая их на спину при постепенно растущей температуре. Её значение, при котором ящерицы не сразу «встают на ноги», и есть предел термостойкости, CTmax. Т.е. это температурный предел способности городских ящериц действовать и, шире, существовать в среде, экстремальной для исходной популяции.
Рекомендуем прочесть
 В эпоху глобального экологического кризиса города — не только «проблема», но и “решение”
В эпоху глобального экологического кризиса города — не только «проблема», но и “решение” Крупнейшие городские агломерации и формы расселения надагломерационного уровня в России
Крупнейшие городские агломерации и формы расселения надагломерационного уровня в России Города как убежища дикой природы
Города как убежища дикой природы Эволюционные модели урбанизации и охрана природы
Эволюционные модели урбанизации и охрана природы Урбанизация как основной фактор негативного влияния на местообитания диких животных Московского региона
Урбанизация как основной фактор негативного влияния на местообитания диких животных Московского региона Урбанизация «диких» видов птиц/млекопитающих как доместикация домашних
Урбанизация «диких» видов птиц/млекопитающих как доместикация домашних Регенеративная урбанизация: Экополис
Регенеративная урбанизация: Экополис Урбанизация и антропогенез: неожиданные параллели
Урбанизация и антропогенез: неожиданные параллели
