![Print PDF В продолжение темы копирования поведения у позвоночных Межвидовое копирование поведенческих стереотипов столь же обычно, как межвидовое Перескажу классическую статью Fisher, Hinde, 1949 об вскрывании городскими синицами крышечек бутылок […]](http://www.socialcompas.com/wp-content/uploads/2024/06/blue-tit-milk-bottle-465x190.jpg "Внутри- и межвидовое копирование поведения, на примере открывания молочных бутылок синицами")

В продолжение темы копирования поведения у позвоночных
Межвидовое копирование поведенческих стереотипов столь же обычно, как межвидовое
Перескажу классическую статью Fisher, Hinde, 1949 об вскрывании городскими синицами крышечек бутылок с молоком, традиционно оставляемым в Англии у домов, а то её цитируют все, но почти никто не читал. Вторая пересказываемая статья их же про то же самое в Nature 1952 г. — выжимка с данной с некоторыми дополнительными новыми наблюдениями про использование этого неожиданного источника пищи. Плюс выскажу некоторые мысли по поводу и в связи, сопоставляя классику с современными данными.
Обе статьи, между прочим, показывают не только рождение и распространение поведенческих инноваций, но и куда более интересное — очень быстро «изобретение» копируется близкими видами, у которых гомологичны двигательные стереотипы (то что Оскар Хейнрот назвал arteigende Triebhandlung — видоспецифическая манера держаться, aTh), и в передвижении, и в кормодобывании. И наоборот, чем дальше виды друг от друга, чем меньше можно говорить о такой гомологии, тем позже и менее полно копируется данный навык, скорей, стимулированные новой возможностью добытия пищи, данные виды изобретают что-то подобное заново. См. например, таблицу числа случаев открывания бутылок несиницами в конце статьи.
Действие животного, особенно хорошо структурованное и стереотипно исполненное, побуждает к его повторению другими животными, видящими его или воспринимающими как-то иначе. Это копирование поведения создано двумя причинами — рост общего возбуждения, увеличивающий общую готовность действовать, и социальное облегчение по Р.Зайонцу, задающее исполнение того же самого действия, а не иного. Во втором случае критически важно сходство форм действия-модели и ответного действия воспринимающей особи: чем оно выше, тем гарантированней ответ и точнее копирование поведения: сильнее и специфичней всего последнее обеспечивается ритуализированными деймонстрациями, несколько менее — ath, типологически определёнными паттернами, слагающими повседневное поведение, ещё меньше — пластичными и изменчивыми прямыми действиями, непосредственно связанными с овладением добычей, спариванием с самкой, спасением жизни и пр.
«Индукция пищевой активности в террариумной группе кавказских гадюк (Vipera kaznakovi), отказывающихся брать корм, достигается подсадкой змеи, хорошо берущей корм. При первых проявлениях пищевой реакции у «новой змеи» (поднятая голова, учащённое мелькание языка), прежде неактивные особи проявляют признаки пищевого поведения. Когда подсаженная змея начинала поиск укушенной жертвы, все змеи в террариуме стали проявлять беспокойство и активно двигаться. Когда охотившаяся змея заглатывала добычу, некоторые соседи делали явные попытки её перехватить, а затем сами начинали охотиться и питаться. Эта стимуляция пищевой активности характерна для ямкоголовых и гадюковых змей, особенно для подвижных змей сем. Colubridae, а среди них более всего батрахофагам, которым свойственны резкие броски на добычу (Ананьева, Орлов, 1983).
При передаче даже столь неспецифических сигналов наиболее существенны особенности динамической формы стимулирующих движений. Стимуляция возможна среди особей одного вида и между разными видами, имеющими однотипные движения и специфические особенности локомоции в момент охоты. Например, красноспинный полоз Elaphe rufodorsalis не реагирует на бросок и удушение лягушки зелёным питоном Chondropython viridis, но его пищевое поведение может быть стимулировано охотой другого вида полозов или ужей Natrix. У полозов и ужей первые стадии охоты принципиально сходны, и лишь затем красноспинный полоз добычу душит и заглатывает, тогда как уж глотает живьём (Ананьева, Орлов, 1983).
Следовательно, для того, чтобы стимулирующее движение «активной» змеи вызвало у «пассивного» животного адекватные действия охоты, а не просто общий подъём поисковой активности, нужно некое минимальное сходство в форме стимулирующих и вызванных охотничьих движений. Тогда первые могут быть сигналом для выбора вторых и одновременно моделью реализации выбранного поведения (моделирование идёт по таким параметрам, как стереотипность отдельных действий в составе реализуемого поведения, устойчивость и направленность развёртывания программы, её долгосрочность и пр.). Также действует всякий коммуникативный сигнал: как символ (знак) он указывает на определённую проблемную ситуацию и на возможности её разрешения в соответствующей системе отношений через запуск специфических программ поведения, допустимых по отношению к партнёру, а как модель он указывает на тот режим реализации выбранного поведения, при котором последнее будет действенным.
Другой пример аналогичного воздействия — половое поведение обыкновенных подвязковых змей Thamnophis sirtalis parietalis в природе и в лаборатории. После спаривания у самцов этого вида интерес к половому поведению пропадает на 12-24 ч., а затем восстанавливается. Половое поведение не спаривавшихся самцов мало чем отличается от такового уже спаривавшихся, то есть имеющийся половой опыт слабо влияет на стереотипы. Обнаружено сильно тормозное воздействие размножающейся пары на половое поведение не участвующего в размножении самца, которое, однако, сразу снимается предоставлением рецептивной самки (Whittier et al., 1985)»
Фридман В.С. От стимула к символу: сигналы в коммуникации позвоночных. Т.1. М.: URSS, 2013.

Иными словами, специфические телодвижения змей при спаривании выступают как сигналы-стимулы, подавляющие половую активность «свободных» самцов, чьё вмешательство может дезорганизовать процесс, наподобие сигналов обороны-угрозы у такырной круглоголовки. В общем случае, чем инвариантней образ телодвижения, чем оно специфичней относительно повседневной активности, тем сильней эффект стимуляции и/или торможения, оказываемый им на однотипные действия других особей в соответствующей ситуации.
Этот эффект объясняется открытым М.Е.Гольцманом правилом стимуляции подобного подобным. Его механизм сам по себе неспецифичен: «работает» с телодвижениями любой степени экспрессии и любой стереотипности исполнения, независимо от их мотивационной подосновы «внутри» и сигнальности «вовне» животного. Однако он нимало не противоречит существованию специфических сигналов, дифференцированные формы которых «вкраплены» в целом бесструктурный поток поведения, как отдельные льдины на реке с шугой, вместе со специфическими внутренними механизмами, поддерживающими стереотипность «продуцирования» характерных форм таких «льдин» в телодвижениях одной особи и одновременно точность распознавания их другой (см. современные данные по нейросетевому анализу проблемы).
Более того, он даже предполагает их появление при эволюционной оптимизации течения соответствующих взаимодействий и функции элементов, внутри них осуществляющих «нужную» особям стимуляцию. Действительно, обмен сигналами, позволяющими «оценить» качество потенциального партнёра, и опережающе скорректировать модель поведения в отношении него, много эффективней обмена демонстрациями, лишь привлекающими последнего и/или возбуждающими обоих; при том, что затраты и рискованность первых сопоставима со вторыми, а часто и существенно меньше. См. пример с малой южной колюшкой Pungitius platygaster.
Соответственно, если самые гомологичные aTh разных видов связаны с перемещением, они будут склонны держаться в общих стаях, двигаться и отдыхать вместе. Скажем, на Северном Кавказе городские сизари стали садиться на ветви деревьев в 1980-1985 г. вслед за вновь проникшими в регион кольчатыми горлицами Streptopelia decaocto. После кормежки на земле рядом с голубями редкие горлицы летели на деревья. Часть голубей им подражала, большинство после двух-трех попыток летели на привычные присады, но некоторые голуби с трудом продолжали пытаться сидеть на ветвях.
К 1990 гг. это стало уже обычным, и совершалось без трудностей. В предковом ареале оба вида хорошо различают друг друга и не следуют друг за другом (наблюдения В.В.Суслова в г.Грозном).
При освоении одним видом нового для него района, где живёт другой, само присутствие его особей делается сигналом для выбора территорий и/или гнездования именно здесь. Скажем, обыкновенная гага Somateria mollissima регулярно залетала на Чёрное море с 1950 г.; утки держались у каменистых островов, напоминающих излюбленные места гнездования на севере, а среди них предпочитали районы, уже заселённые гоголем Bucephala clangula и средним крохалем Mergus serrator. Близкие виды, с которыми гага сообитает на севере, как бы «сигнализировали», что данная местность — подходящая или даже предпочтительная.
Однако физиономические сходства новых местообитаний с исходными — лишь отдельный момент, притягательный для птиц, но другие, сплавленные с ним в общую амальгаму условий существования на данной территории, — новые, необычные, требующие изменения биологии вида. И они появляются, причём сразу соответствующие новым условиям, с которыми птицы не сталкиваются, отбору не приходится их «переделывать».
В 1975 и 1977 гг. гага загнездилась на о.Долгий и Круглый Черноморского заповедника, к 1986 г. её численность выросла с 2 до 118 пар, и доныне устойчива. Однако здесь гаги гнездятся иначе: 90-95% гнёзд в тростнике, >50% в береговых зарослях. Главное уткам — близость открытой воды, а не степень укрытия: многие шалашики, настилы, ящики и брёвна не используются. В тростнике возникают своего рода колонии гаг, а 2-10% уток гнездится совершенно открыто, на выбросах и в колониях чайки-хохотуньи (Ардамацкая, 2007). В 1980-х гг. ареал расширяется на соседние острова, и к 1998 г., с гнездованием на о.Джарылыгач и Каланчакских о-вах (~150 км по побережью от первоначальных мест поселения). Сходным образом длинноносый крохаль Mergus serrator до середины ХХ в. на Чёрном море лишь зимовал. С 1925 г. вид загнездился в северо-западном Причерноморье, в Крыму с 1950 г. (Аверин, 1951). Численность вида в новых местах гнездования в 1930-1970-х гг. возрастала, достигнув максимума в 900 гнёзд. В 1980-х гг. она ещё оставалась довольно высокой — 600-700 гнёзд, но рухнула в следующие десятилетия — до 50-60 пар в 2000-2004 гг., 40 в 2005 г., и 10-15 в 2006-2010 гг. (Руденко, 2011). Причиной указывается неблагоприятная средовая динамика («ухудшение экологического состояния заливов Чёрного моря (загрязнение, изменение гидрологического состояния), ухудшение условий гнездования (высокий уровень воды в заливах, что приводит к затоплению гнёзд, постепенному разрушению островов и сокращению пригодной для гнездования территории)»), однако не меньшую роль может играть сокращение подтока вселенцев из основной части ареала на фоне его общего отступления к северу и снижения численности вида в Европейской России (Костин и др., 2021), при недостаточности воспроизводства в самом изоляте при том уровне разлёта молодых, который имеет место быть у этих уток.
«Оценка» качества местообитаний по присутствию и поведению (как часто тревожатся или, напротив, спокойно кормятся и поют) других видов орнитокомплекса — норма для самых разных групп птиц, подробней всего изучавшаяся у массовых видов дендрофильной авифауны (Seppänen et al., 2007; Magrath, Bennett, 2011; Szymkowiak et al., 2017). Эффекты вторичных сигналов сильны и долговременны: побуждают гнездиться на следующий год даже в неподходящем местообитании (данные по Dendroica caerulescens, Betts et al., 2008), гнёзда, пение или тревожные позывки этого года влияют на выбор мест гнездования в следующем (Nosera et al., 2005),
Здесь важны видовые особенности: одни и те же сигналы у одних видов (боболинк Dolichonyx oryzivorus) формируют территориальную связь молодых даже с неподходящим местообитанием, у других (овсянка Passerculus sandwichensis) — нет (Nocera et al., 2005, 2009). Другой пример. Позже прилетающие болотные камышевки Acrocephalus palustris поселялись раньше и в большем числе на участках, где имитировалось присутствие и пение теньковки Phylloscopus collybita, но избегали участков, занятых славкой-черноголовкой Sylvia atricapilla. Оба вида, в свою очередь, привлекаются пением конспецификов (Szymkowiak et al., 2017).
Если самые гомологичные aTh связаны с кормодобыванием (способами обследования субстрата и/или собственно методами взятия кормовых объектов), данные виды с высокой вероятностью кормятся в смешанных стаях и копируют друг у друга места кормления (как вообще выборы микростации для обследований вообще, так и кормление в данном конкретном месте) и/или кормовые методы.
Так, в опытах американских хохлатых синиц Parus bicolor приучили брать мучных червей в определенных лунках искусственного кормового дерева на глазах у других видов птиц – обычных членов синичьих стай: каролинских гаичек, каролинских поползней и пушистоперых дятлов.
Другое однотипное дерево рядом с «обучающим» было «миметическим» и содержало корм в сходных позициях и разной степени доступности. Затем загородку убирали, и следили за поведением высокои низкоранговых членов стаи. Первые концентрировали внимание именно там, где кормилась модель; вторые искали в таких же местах на другом дереве, и проникали в субстрат теми же кормовыми методами (Waite et al., 1988).
Обучение кормлению там же и тем же методом, что и «модели», обычно между видами смешанных синичьих стай (Dhondt, 1989; Ekman, 1989) и многократно воспроизводилось в эксперименте (Иваницкий, 1986; Sasvari, Hegyi, 1994). Если доминантные виды легко сближаются с кормящимися подчиненными, выучивая их место и метод кормления, то подчиненные избегают сближаться с кормящимися доминантами, хотя, наблюдая, выучивают не хуже [Там же]. Выученные кормовые методы других видов используются и в отсутствие «модели» в поле зрения, хотя и недолго (Galef, Giraldeau, 2001; Farine et al., 2012; 1986; Sridhar, Shanker, 2014).
Современные данные показывают, что между внутривидовым и межвидовым копированием поведения нельзя провести грань, второе идёт с той же лёгкостью и свободой, что и первое. Или, наоборот, с нелегкостью и несвободой — важно, что той же самой, с поправкой на гомологию aTh. Это верно как для видовых стереотипов, aTh, так и для поведенческих инноваций.
Откупоривание молочных бутылок синицами
Вернёмся к английским синицам. Впервые вскрывать крышки бутылок с молоком оба вида синиц начали в 1921 г. (Swaythling близ Саутгемптона). В следующие 30 лет этот кормовой метод распространился по юго-востоку Англии, и появился в других местах: вокруг Белфаста, позже и на севере Англии (Fisher, Hinde, 1949, c.348-349).
Позже он появляется в ряде других районов Великобритании и Европы (Швеция, Дания, Голландия), также поддерживаясь обучением. Так, с 1947-1948 гг. оба вида синиц регулярно открывали бутылки минимум в 21 местности южнее Зейдер-Зее. Дальше >10 видов птиц были замечены как за открыванием молочных бутылок, так и питьём из тех, которые не обязательно открыли сами.
Среди них домовые воробьи, чёрные дрозды, зарянки, московки Parus ater, певчие дрозды и лесные завирушки (Fisher, Hinde, 1949, таблица; Lefebre, Aplin, 2017, fig.6.1). В Скандинавии помимо синиц бутылки открывали сороки, чёрные дрозды, галки, большой пёстрый дятел (регулярное открывание бутылок с молоком в Копенгагене). В Сиэттле (США) молочные бутылки открывали голубые сойки Cyanocitta cristata (Hinde, Fisher, 1951; Fisher, Hinde, 1952). В дальнейшем к ним добавились серые вороны и болотные гаички Parus palustris; в эксперименте это начинали делать черноголовые гаички, хотя не за счёт копирования «учителя» (Lefebre, Aplin, 2017).
Напротив, для лазоревок и больших синиц копирование открывания молочных бутылок и, вообще, вновь изобретаемых способов доступа к скрытому корму строго доказано в специальных экспериментах.
Для анализа темпов распространения инновации и её освоения прочими видами исследовательский комитет British Trust of Ornithology (ВТО) опросил его членов и местных натуралистов. Ответы получены от 126, давших 141 наблюдение об открывании молочных бутылок синицами разных видов в какой-то местности. 18 таких наблюдений получено от членов местных обществ естествоиспытателей, 43 — из писем в British Medical Journal и Lancet, т.е. от лиц с научной подготовкой. Ещё около 200 таких наблюдений получено из заметок в местной прессе или личного опроса орнитологами; в 223 случаях наблюдатель отметил, что был в той же местности в предыдущем году, наблюдал, но открывания пока что не видел, но увидел бы его, если б оно было. Это позволяло определить год появления инновации: в остальных случаях можно было фиксировать только ее наличие, но момент появления пропустили.
Эта инновация возникает толко там, где бутылки с молоком оставляют у домов, поэтому её нет в большинстве сельских районов Ирландии. Однако инновация может не появляться ещё много лет после того, как подобная практика доставки молока станет обычной в какой-то местности — и так было в 23 из 30 районов наблюдений, т.е. подобная инновация — это поведенческая новизна, синицы не предрасположены к этому и не осваивают эту возможность немедленно. В этих 23 местностях удалось точно определить год появления инновации.
Главные открыватели бутылок – синицы большая Parus major, лазоревка P.caeruleus и московка Paris ater — в Великобритании оседлы, даже при расселении молодых не перемещаются больше чем на 15 миль. Поэтому встреча открываний бутылок на большем расстоянии считалась самостоятельным «изобретением» местных птиц, а не копированием у ближайших соседей.
Распределение мест и лет наблюдений показывает, что этот источник пищи «открывается» лишь небольшой частью популяции, и затем передаётся ближайшим соседям, но не более того. Т.е. инновация распространяется из нескольких очень локальных центров, где изобретается независимо (в Англии, Уэлльсе, Шотландии и С.Ирландии, см.распространение в окрестностях Белфаста). Лучше всего это видно в Восточной Англии, где сеть наблюдателей плотнее всего.

Рис.1. Карты распространения вскрываний бутылок в Великобритании по сообщениям членов British Trust for Ornithology (ВТО) и другим (в основном местных натуралистов), каждая карта — встречи до и включая указанный год. Карты те же что использованы для национальных учётов птиц, с квадратами 5 * 5 км, точка карты соответствует 15 кв. миль.
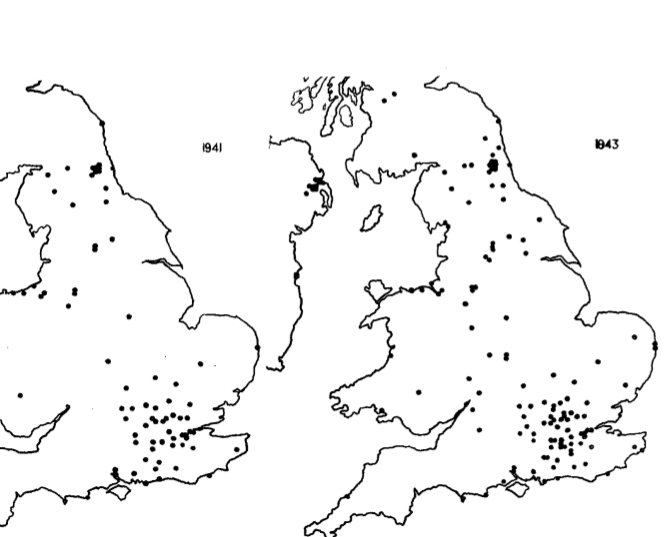
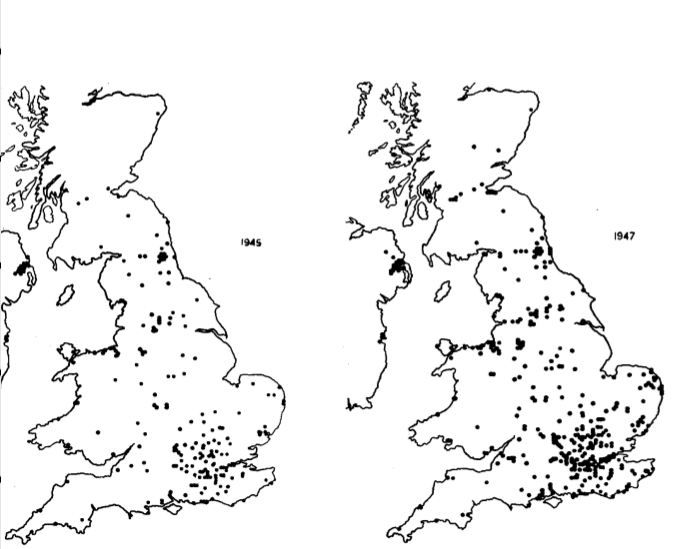
Всех приславших сведения просили специально отметить, если они были в этом же месте в прошлые годы перед первой встречей вскрывания, и вели наблюдения за теми же птицами, т.е. если бы был уже этот навык они б могли его видеть — но не видели.

Рис.2. Большие синицы и лазоревки прокалывают крышки бутылок с молоком. Фото V.L. Breeze. Источник Fisher, Hinde, 1949.

На рис.3 видно, что в первые годы наблюдения открывания единичны, а затем нарастают лавинообразно, когда новый кормовой метод «захватывает» всю популяцию. Однако распространяется он лишь к ближайшим соседям. Также она распространялась из мест первого обнаружения везде где это могли проследить за несколько последовательных лет: в Ковентри, Лондоне, Лланели и пр. Часто открывание присутствовало в одной местности, но годами отсутствовало в соседней, хотя там тоже ставили молоко и были наблюдатели — например в Little Aston у Бирмингема было, но не отмечено в соседних населённых пунктах Streetly and Sutton Coldfield.

Рис.3. Карта Белфаста и окрестностей: показаны годы начала вскрывания крышек в разных местах. Цифры в кружках показывают наличие наблюдателя здесь в предыдущем году: возникни кормовой метод тогда, он был бы замечен.
Синицы много чаще вскрывают бутылки зимой, чем в другие сезоны, в том числе потому что зимой их делается больше в городах. Отмечали и следование птиц за тележкой молочника, и вскрывание крышечек на ходу. Обычно крышки вскрывают уже через несколько минут после оставления бутылок.
Способы открывания очень различны. Крышечку из фольги птицы прокалывают, а затем открывают её тонкими полосами, иногда снимая её целиком, иногда только делают в ней отверстие. Картонные крышки также вскрывают разными способами. Их снимают целиком, поддевая клювом, или пробивают отверстие в центре, или отрывают картон слой за слоем, пока он не истончится достаточно, чтобы можно было проколоть отверстие.
В каждом районе встречается несколько способов вскрытия и наоборот, одна и та же птица может использовать несколько: скажем, отмечено последовательное использование большой синицей двух разных способов на двух разных бутылках, после того, как с первой птицу вспугнули. На первой птица пробила крышечку в центре, на второй пробка уже было слегка подснята, птица увидев это поддела её клювом и сорвала целиком, даже не пробуя пробивать. Когда крышка срывается целиком, под ней часто остаются сливки — то, ради чего птицы стараются.
Подобные крышки птицы часто уносят и выклёвывают еду в удобных местах, где накапливаются целые россыпи крышечек. Двое наблюдателей предлагали просто крышечки птицам, и синицы пробовали их клевать. Прокалывая крышку, синицы также пьют молоко из бутылок, до 10-20 г., не только берут сливки. Иногда они жадничают, опускают голову слишком глубоко, теряют равновесие и захлёбываются.
В более чем 400 наблюдениях вид синиц могли определить точно. В 246 из них это были лазоревки (у них клюв сильней, и пробивает более толстую фольгу/отрывает более крепко посаженную крышку), 142 — большие, 11 — московки и 1 — болотные гаички Parus palustris. Вскрытие крышек московками отмечено в ю.-в. Англии (7 мест), в Сомерсете (1), южном Уэлльсе (1) и центральной Англии (2). Вскрытие болотными гаичками видели лишь в St. Leonard’s, Сассекс, в 1942 г., в районе, где 3 других вида синиц успешно делали это с 1941 года (!).
Авторы пишут, что черноголовые гаички Parus atricapillus не открывают бутылки с молоком в США, но в дальнейшие годы и они этим занялись.

Рис.4. Несиницы, замеченные за открыванием молочных бутылок. Строки сверху вниз — виды: домовый воробей, чёрный дрозд, скворец, зарянка, зяблик, лесная завирушка. Столбцы — число наблюдений, как эти птицы сами открывают бутылки (левый), или как пьют из бутылки, открытой не ими (правый).
Рекомендуем прочесть
 Пение самок воробьиных птиц: исключение или закономерность?
Пение самок воробьиных птиц: исключение или закономерность? Быстрая перестройка фенологии в новых местах гнездования ласточек
Быстрая перестройка фенологии в новых местах гнездования ласточек Что не так с критикой Ильенкова
Что не так с критикой Ильенкова Человеческая деятельность формирует зимнюю экологию перелётной птицы, славки-черноголовки
Человеческая деятельность формирует зимнюю экологию перелётной птицы, славки-черноголовки Отбор адаптивной и неадаптивной пластичности в экспрессии генов при приспособлении анолисов к городскому «острову тепла»
Отбор адаптивной и неадаптивной пластичности в экспрессии генов при приспособлении анолисов к городскому «острову тепла» Вибрационные сигналы в коммуникации животных
Вибрационные сигналы в коммуникации животных Как соотносятся прямое приспособление и эволюционные изменения?
Как соотносятся прямое приспособление и эволюционные изменения? Рынок и супермаркет
Рынок и супермаркет
