

Североамериканская форма касатки, Hirundo rustica erythrogaster
Резюме. Продолжение про быструю эволюцию в связи с изменением миграционного поведения у птиц. Американский подвид касатки и белолобая ласточка загнездились в местах зимовок в Аргентине, в связи с чем быстро и радикально сменили «расписание» стадий годового цикла, прежде всего линьки, и обрели новые места зимовок, соответствующие таковым местных перелётных видов. Важно, что новообразованные популяции успешны, их численность растёт, один из факторов роста приспособленности, как и для новых зимовок славки-черноголовки — сокращение пролётного пути.
***
Продолжаем про быструю эволюцию в связи с изменением миграционного поведения у птиц, сейчас в связи с гнездованием двух североамериканских ласточек в местах их зимовок в Аргентине. Быстрые перестройки времени линьки и, вообще, “расписания” стадий годового цикла были описаны у числа касаток Hirundo rustica erythrogaster (Winkler et al., 2017) и, независимо от них, белолобых скалистых ласточек Petrochelidon pyrrhonota (Areta et al., 2021), вдруг в небольшом числе загнездившихся в районе зимовок в Аргентине. Первый вид поселился там в начале 1980-х, новая популяция вполне успешна: растёт численно, ареал вырос на ~500 км.
Рост приспособленности вызван сокращением пролётного пути примерно втрое, с ~20000 км в обе стороны для исходной популяции с востока Канады, до не более 14600 км у аргентинских касаток. Они также избегают длительного беспосадочного полёта над морем (Мексиканским заливом или западной Атлантикой), предпринимаемого частью птиц из исходной популяции, и делающего перелёт максимально напряжённым и сильно более рискованным.
Одна гипотеза связывает гнездование в местах зимовок с глобальным потеплением: ласточки поселились, когда выросли температуры (и особенно минимальные) в южной части провинции Буэнос-Айрес. Другая, более вероятная, – с распространением в последние 100 лет бетонных мостов, удобных для гнездования, и асфальтированных дорог – для охоты. Бетонные мосты начали строить в Аргентине в 1890-е гг., наиболее распространёнными они стали в 1930-е, первые гнёзда на таком подходящем субстрате отмечены через 50 лет. Проводившие лето отдельные особи там отмечались и раньше (как некоторые североамериканские птицы остаются на лето сейчас), но не гнездились (Winkler et al., 2017).
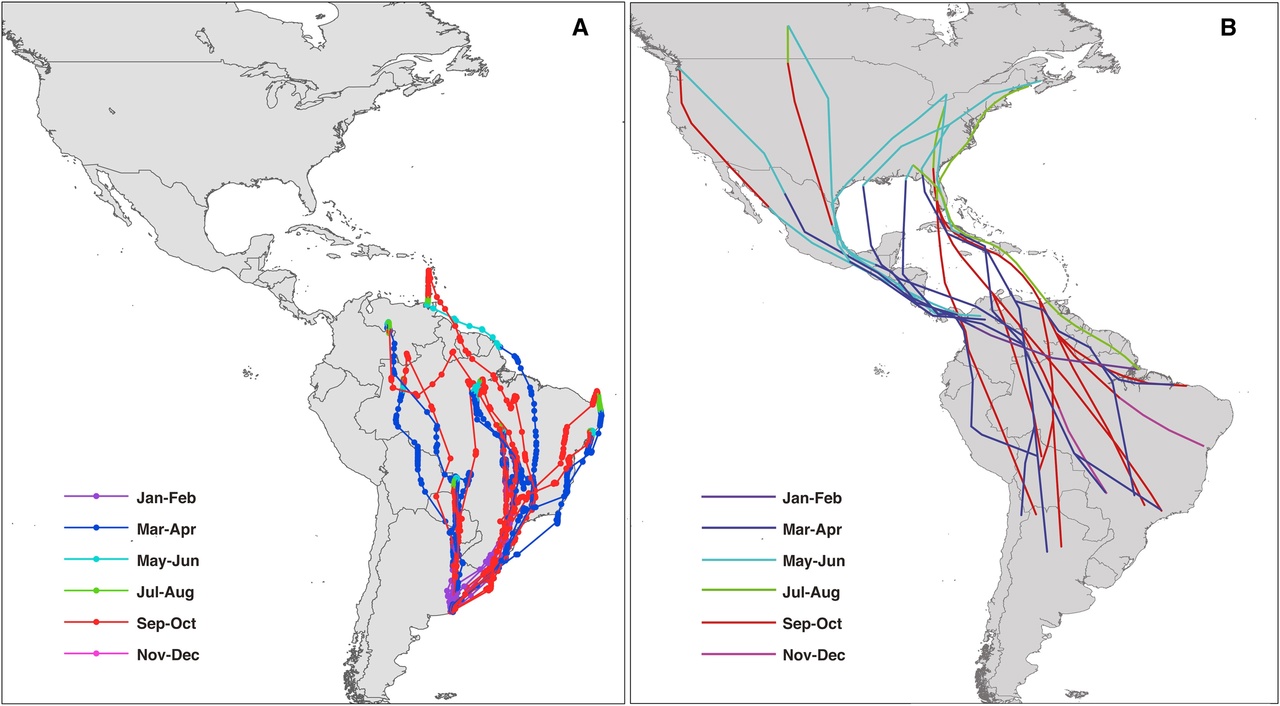
Касатки из Аргентины не начали зимовать в Северной Америке, как было бы ожидаемо исходя из обычной логики “противоположного полушария” (рис.1А). Геолокаторы, установленные в 2014 г. на девяти птицах, выявили зимовки в широкой полосе севера Южной Америки, от Венесуэлы до Центральной Бразилии. Здесь же проводят зиму мигранты, автохтонные для Аргентины. Собственно места зимовок и конкретные пути к ним пока неустойчивы, и у каждой птицы свои – подобно тем пеночкам-зарничкам Ph.inornatus, что осенью движущимся в Европу вместо традиционных зимовок (Winkler et al., 2017).
Однако птицы уже двигались по двум достаточно чётким пролётным путям: 6 пересекли тропик Козерога вдоль побережья и севернее Сан-Паулу или продолжили двигаться по нему, либо летели прямо через зону серраду на север Венесуэлы. Три птицы летели в глубине континента: на север провинции Буэнос-Айрес, пересекли тропик по руслам рек Парана и Уругвай и зазимовали там, либо над влажными тропическими лесами Амазонии отправились на север Южной Америки. Несмотря на широкий географический разброс места зимовок, переход к миграциям в годовом цикле достаточно сжат (3 недели), что сравнимо с исходной североамериканской популяцией: птицы оставили места гнездования между 13 февраля и 6 марта, вернулись туда между 25 сентября и 26 октября (Winkler et al., 2017).
Важно что это – быстрая перестройка, видимо, уже в первом-втором поколении колонистов, не постепенные изменения ориентации миграционного поведения и его продолжительности естественным отбором. Хотя у воробьиных-дальних мигрантов то и другое детерминированы наследственно, и довольно легко изменимы отбором, постепенные изменения были бы гибельны для большинства аргентинских ласточек. Хотя бы частичное следование прежнему маршруту “на юг” с прежней длительностью перелёта гарантированно привело бы птиц в воды Атлантики. При подобном уровне малая популяция колонистов гарантированно погибнет раньше, чем прежний маршрут перестроится в новый “на север”, для малой популяции, особенно у достаточно медленно размножающихся позвоночных, всякая гибель особей существенна и непоправима, независимо от того, случайна она или связана с успешным приспособлением под действием отбора, то и другое равно подрывает её жизнеспособность (Winkler et al., 2017).
Случайная или вызванная отбором, вырабатывающим изменения благоприятные в будущем, всякая элиминация подрывает её жизнеспособность здесь и сейчас. Как показал В.В.Суслов (Suslov, 2013), при быстрых приспособительных изменениях в малых популяциях (а прочие варианты крайне редки) две главных проблемы – снизить плату за отбор и “обойти” дилемму Холдейна. Первое важно везде и всегда; второе же тем важней, чем по большему числу параметров требуется приспосабливаться в ответ на происходящую трансформацию, т.е. в общем случае чем сильнее изменение осваиваемых местообитаний сравнительно с исходными и экстремальней новые условия существования. Не вымереть, продолжить себя в следующих поколениях важней эффективных приспособлений в этих последних – а всякий эффективный отбор неотвратимо снижает жизнеспособность, о чём давно знают селекционеры.
Даже если потенциальна возможна выработка искомого (“северного”) пути миграций за счёт селективных процессов, при известных наследуемости признаков, составляющих “нужное” изменение2 и продолжительности жизни ласточек они будут а) слишком медленны и б) вызовут элиминацию, “несовместимую с жизнью” данной популяции.
 А.
А.
 Б.
Б.
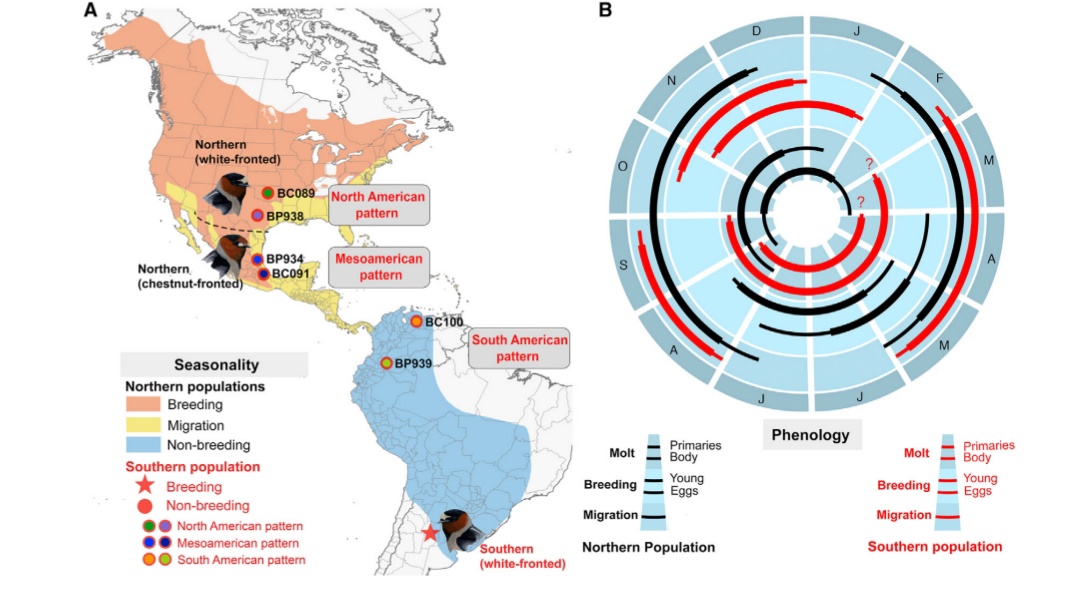
Рисунок 1. Изменения биологии касаток, загнездившихся в Аргентине. А. Слева — реконструированные пути миграций и места зимовок 9 специально прослеженных особей, отрезки пути окрашены в разные цвета сообразно месяцам (подписаны снизу); Справа – пути миграций и места зимовок североамериканских касаток по данным литературы. Б. Сдвиг “расписания” стадий годового цикла при гнездовании в Аргентине (справа, красным) сравнительно с таковым исходной североамериканской популяции, вероятней всего, с востока Канады (слева, чёрным). Сектора круга – месяца года, стадии сверху вниз: линька – первостепенных маховых, контурного оперения, размножение – вылупление птенцов и откладка яиц, миграция.
Другая аналогичная перестройка – сдвиг “расписания” стадий годового цикла сообразно местным условиям (иначе вылупление птенцов “разошлось” бы с максимумом воздушного планктона). Благодаря этому ласточки линяют в нерепродуктивный сезон, как и в исходной популяции. Однако на местах гнездования в Аргентине отмечали отдельных линяющих особей. Видимо, нечастый, но регулярный подток птиц материнской популяции продолжается. Он организован скорее всего по типу “сортировки” особей по поведенческим потенциям – из зимующих североамериканских птиц к аргентинским колониям присоединяются именно те, для кого эти новые места обитания наиболее благоприятны (Winkler et al., 2017).
Большая часть роста численности/расширения ареала аргентинских касаток обеспечена размножением особей новой популяции. Однако на гнездовых колониях единично попадаются ласточки, находящиеся в процессе линьки маховых или с остатками ювенильного оперения. Скорее всего это птицы исходной популяции, т.е. есть некоторый приток из исходной популяции в дочернюю, что подтверждается отсутствием “эффекта основателя” и заметной генетической дивергенции у второй (Winkler et al., 2017).
Однако до сих пор неясна биология этих особей-первопроходцев. Это исключительно молодёжь, родившаяся в Северной Америке <6 месяцев назад или ласточки, уже размножавшиеся там, тоже становятся колонистами в Аргентине? Птицы размножаются сразу по прибытию туда или первое лето в новых местах проходит без размножения? Независимо от возраста иммигрантов, каково влияние на их физиологию шестимесячного сдвига в “прохождении” годового цикла, с учётом осеннего возбуждения, увеличения гонад, осеннего пения и пр.? Объясняется ли фотопериодом гнездование именно на южном краю прежнего района зимовок? (Winkler et al., 2017).
Как именно шло “расщепление” прежней популяционной системы в связи с гнездованием в районе зимовок, возможно, показывают белолобые скалистые ласточки, у которых данный процесс происходит прямо сейчас (Areta et al., 2021). Небольшое число их загнездилось в центральной Аргентине (г.Вилья Мария, провинция Кордова) в 2015 г, ~в 8000 км от прежних мест гнездования (возможно после неудачных попыток там). Уже после второго (в 2017 г.) и четвёртого (2019 г.) сезонов размножения благодаря геолокаторам, повешенным на 18 птиц, из которых потом было отловлено 6, выяснились миграционные пути и места зимовок (Areta et al., 2021).
Их разбросало на широком пространстве (юг США, возможно их прежние места гнездования; Мезоамерика, самая южная часть гнездового ареала вида; север южной Америки, Венесуэла и Колумбия). Места зимовок, отмеченные в 2017 г., были воспроизведены в 2019 г. С новых мест размножения птицы улетали в середине февраля-начале марта, и возвращались в середине сентября того же года. Миграционные маршруты всех птиц в оба конца были сходны (через западную часть континента восточнее Анд), различались только конечные точки – т.е. общая длительность (Areta et a., 2021).
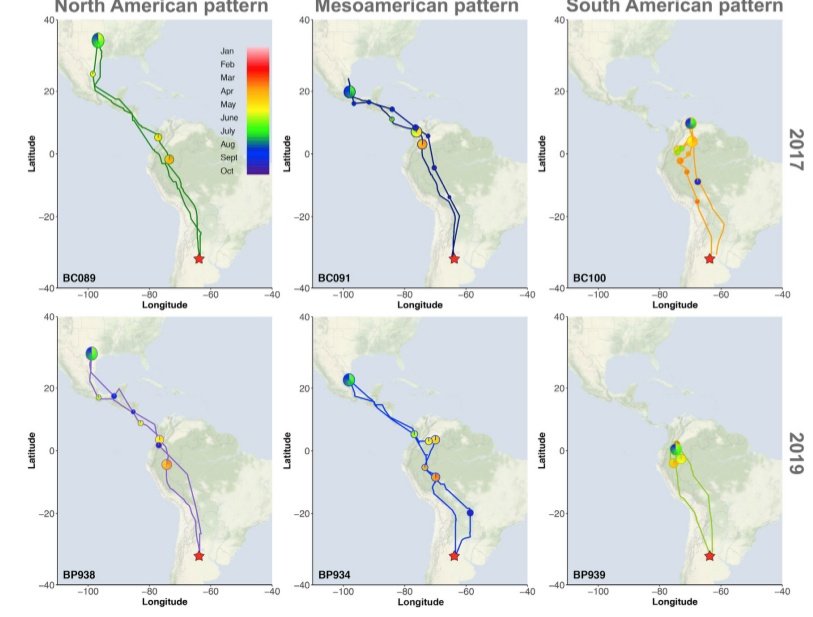
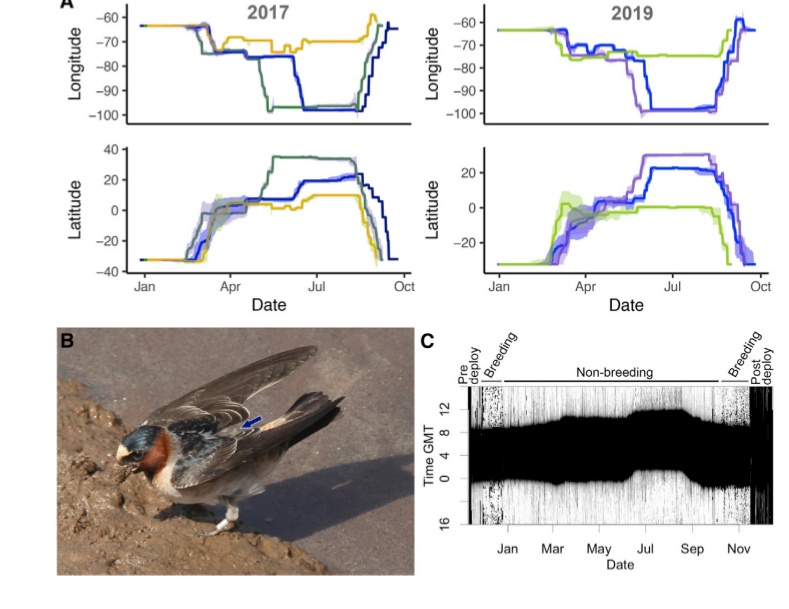 Рисунок 2. Изменения в биологии скалистых ласточек, загнездившихся в Аргентине. А. Миграционные пути 6 птиц, прослеженные по геолокаторам. Кружки — места остановок, цвета соответствуют разным месяцам в которые происходило движение. В. А. Наложение миграционных путей этих 6 птиц на карту изменения километража (сверху) и широты (снизу) в 2017 и 2019 гг. В. Одна из этих птиц, собирающая грязь для строительства гнезда в 2017 г., кроме геолокатора, помечена и кольцом. С. Изменения освещённости, даваемые геолокаторами (тёмное — ночь, светлое — день) в наложении на стади годового цикла. Происходило ли размножение, видно по ежедневному «частоколу» светлых и тёмных полос, когда ласточки часто лезут в закрытое гнездо. Графическое резюме статьи: слева гнездовой ареал вида в Северной Америке, пролётные пути, места зимовок прежние и новые по данным мечения (обведённые кружки), новое место гнездования. Справа — сдвиг прохождения стадий годового цикла на 6 мес. на новом месте размножения (красным) сравнительно с прежним (чёрным).
Рисунок 2. Изменения в биологии скалистых ласточек, загнездившихся в Аргентине. А. Миграционные пути 6 птиц, прослеженные по геолокаторам. Кружки — места остановок, цвета соответствуют разным месяцам в которые происходило движение. В. А. Наложение миграционных путей этих 6 птиц на карту изменения километража (сверху) и широты (снизу) в 2017 и 2019 гг. В. Одна из этих птиц, собирающая грязь для строительства гнезда в 2017 г., кроме геолокатора, помечена и кольцом. С. Изменения освещённости, даваемые геолокаторами (тёмное — ночь, светлое — день) в наложении на стади годового цикла. Происходило ли размножение, видно по ежедневному «частоколу» светлых и тёмных полос, когда ласточки часто лезут в закрытое гнездо. Графическое резюме статьи: слева гнездовой ареал вида в Северной Америке, пролётные пути, места зимовок прежние и новые по данным мечения (обведённые кружки), новое место гнездования. Справа — сдвиг прохождения стадий годового цикла на 6 мес. на новом месте размножения (красным) сравнительно с прежним (чёрным).
Двух кладок за год не отмечено, хотя часть особей лето северного полушария провели в новых местах гнездования. Описанный выше шестимесячный сдвиг “расписания” стадий годового цикла, особенно линьки и размножения, видимо, произошёл уже в первом поколении колонистов, чем могла поспособствовать особенность линьки молодых в исходной популяции. У части молодых предварительная линька задерживается, что ставит как бы вне годового цикла. В популяциях Северного полушария подобные особи скалистой ласточки, как и касатки, не размножаются в первую весну-лето жизни. Летом южного полушария, на второй в жизни зимовке, они в возрасте 1,5 лет реализуют нерастраченный репродуктивный потенциал, тем самым одновременно создав новую популяцию, и сдвинув “расписание” годового цикла (Areta et a., 2021).
Оперение лба аргентинских ласточек белое, т.е. они происходят из северных популяций вида; у более южных, гнездящихся в Мексике, оно рыжее. Особи, зимовавшие в Мезоамерике, находились как раз в гнездовом ареале вторых. Иными словами, даже потенциально в места своего рождения вернуться зимовать могли только две особи из шести, и то это крайне маловероятно. Т.е. скалистые ласточки в новых местах гнездования испытывают практически одномоментную перестройку жизненного цикла и миграционного поведения, в целом соответствующую новым условиям. Скорей всего, здесь видишь первые шаги процесса, вовсю развернувшегося у касаток (Areta et a., 2021).
Другой важный момент: данные по юго-западной Небраске (те же ласточки, что отбираются на короткокрылость при гнездовании близ автострад) показывают мощный селективный контроль сроков прилёта. Аномально холодная и сырая весна 1996 г. была гибельной для слишком рано прилетающих птиц. Прилёт на колонии, прохождение репродуктивного цикла в 1997 г. происходили значимо позже сравнительно с весной 1993 г., хотя их погодные условия не отличались. Анализ метеоданных предшествующих 51 лет показывает, что элиминация слишком рано прилетающих ласточек достаточно продолжительным приходом холодов должна была иметь место минимум 25 раз. До 4 дней возврата холодов вызывают значимую, но невысокую гибель ласточек, 5 дней и более – значительную. Первое отмечали в 1953, 1954, 1972 и 1976 гг. (дважды), 1982, 1983, 1988, 1991, 1992, 1993 гг., второе – в 1953, 1958, 1959, 1961, 1967 (дважды), 1970, 1978 (дважды), 1994, 1995 (дважды) и 1996 гг. 20 событий из 25 пришлись на апрель-первую неделю мая, вызванная ими значительная смертность происходит в среднем через каждые 2,3 года (1-6 лет) (Brown, Brown, 2000).
Рекомендуем прочесть
 Птицы против летучих мышей
Птицы против летучих мышей Манипуляционно-исследовательская активность у морских птиц: поведенческая реакция на предъявленные на колонии искусственные объекты
Манипуляционно-исследовательская активность у морских птиц: поведенческая реакция на предъявленные на колонии искусственные объекты «Рациональное» и «иррациональное» в поведении птиц: сочетание рационально-вероятностной и релизерной моделей поведения при принятии решения о выборе места гнездования
«Рациональное» и «иррациональное» в поведении птиц: сочетание рационально-вероятностной и релизерной моделей поведения при принятии решения о выборе места гнездования Птицы и муравьи не мешают друг другу есть гусениц
Птицы и муравьи не мешают друг другу есть гусениц Красивое доказательство «семантичности» демонстраций у птиц
Красивое доказательство «семантичности» демонстраций у птиц Поведенческие изменения в «городских» популяциях птиц — нет общего с одомашниванием
Поведенческие изменения в «городских» популяциях птиц — нет общего с одомашниванием Пресечение урбанизации птиц как зеркало московского экоцида
Пресечение урбанизации птиц как зеркало московского экоцида Человеческая деятельность формирует зимнюю экологию перелётной птицы, славки-черноголовки
Человеческая деятельность формирует зимнюю экологию перелётной птицы, славки-черноголовки
