
Александр Махров
Признаки и призраки
Содержание
Почти 200 лет назад Жан-Батист Ламарк сформулировал закон „Всё, что природа заставила особей приобрести или утратить под влиянием обстоятельств, в которых с давних пор пребывала их порода, и, следовательно, под влиянием преобладающего употребления известного органа или под влиянием постоянного неупотребления известной части, — всё это она сохраняет путём размножения в новых особях, происходящих от прежних, если только приобретённые изменения общи обоим полам или тем особям, от которых произошли новые“.
Этот „второй закон“ Ламарка и вызвал волну споров, докатившуюся до наших дней Сторонников данного утверждения до сих пор зовут ламаркистами, хотя оно было лишь небольшой частью учения Ламарка. Этому учению часто противопоставляют эволюционное учение Дарвина (см , например, „Химию и жизнь“, 2001, № 1), хотя противопоставление здесь, вообще говоря, неправомерно. Дарвин признавал наследование приобретённых признаков. В первой же главе „Происхождения видов“ он прямо писал „Изменённые привычки оказывают влияние, передающееся по наследству“. Так что, как видим, нет никаких оснований ссорить призраки двух великих учёных.
Предмет спора между тем остаётся. Однако, чтобы быть корректными, мы должны говорить не о „ламаркистах“ и „дарвинистах“, а о сторонниках или противниках наследования благоприобретённых (то есть полезных организму) признаков. „Химия и жизнь“ часто давала слово как тем, так и другим (1978, № 12, 1981, №11, 1984, № 2, 1989, № 2, 1997, № 4). В 2003 году на страницах журнала прошла целая дискуссия по этой проблеме (№ 2, 4, 6).
К сожалению, учёные часто вели спор в теоретическом ключе, обращая внимание на разные факты и строя каждый свою линию доказательств, не оглядываясь на оппонентов. Линии эти почти не пересекались, и у читателей могло создаться впечатление, что специалисты просто расходятся в трактовке тех или иных фактов, и решить, кто прав, — дело вкуса. Например, противники наследования благоприобретённых признаков никак не комментируют книгу Э Стила с соавторами „Что, если Ламарк прав?“, на которую часто опираются их оппоненты.
Кроме того, обсуждение касалось в основном так называемой центральной догмы молекулярной биологии — утверждения о невозможности передачи информации от белков к нуклеиновым кислотам. Однако проблема наследования благоприобретённых признаков далеко не исчерпывается возможностью или невозможностью осуществить такой информационный переход. Поэтому давайте разберёмся, что же всё-таки нужно, чтобы унаследовать благоприобретённые признаки, и нет ли на этом пути непреодолимых препятствий.
А нужно, во-первых, новые признаки приобрести. Во-вторых, оценить их благо, то есть выделить из многих других, ненужных или бесполезных. И наконец, избирательно передать полезные признаки потомкам (преодолеть барьер Вейсмана, препятствующий переходу информации между обычными, соматическими клетками тела и половыми клетками).
Можно ли приобрести признаки?
На первый взгляд приобрести новые признаки очень просто Каждый из нас может ходить в спортзал и увеличивать объём мышц или сидеть в баре и увеличивать объём печени. Но и на физические нагрузки, и на алкоголь разные люди реагируют по-разному, что определяется уже генетической программой. Именно конкретная комбинация генов, полученных от родителей, задаёт пределы, в которых мы можем развить тот или иной орган. Таким образом, способность менять признаки в результате „упражнения“ — это результат, а вовсе не причина эволюции.
Возможности любой программы ограниченны: вспомните притчу о цыганской лошади, которую хозяин отучал от еды. Кляча совсем уж было привыкла к такому обращению, да как-то не вовремя околела.
С ограничениями мы сталкиваемся повсюду: мощный холодильник может разморозиться в жару, а современный компьютер проиграть битву вирусу — и это нас не удивляет. Никто не ждёт, что даже очень умная техника сделает что-то полезное помимо того, что в неё заложено. Почему же от организмов ждут целесообразной реакции вне генетической программы? Откуда ей взяться?
Если быть внимательным и добросовестным, то можно разглядеть, что почти всегда за благоприобретённым признаком стоит уже существующая генетическая программа, которая просто не проявляла себя до поры до времени. Ведь, как и компьютер, организм может содержать несколько разных программ и активировать их только в определённых условиях. Так, я уже писал в „Химии и жизни“ (2002, № 10) о пресноводных карликовых лососях и крупных рыбах того же вида, нагуливающихся в море, — это разные программы развития, заложенные в одном и том же организме.
Какая из них активируется в каждом конкретном случае, зависит от условий среды: меняя их в эксперименте, можно получить зрелых трёхсотграммовых рыбок, никогда не видевших моря, или десятикилограммовую сёмгу, проводящую большую часть жизни на просторах Атлантики. Но вот, скажем, со щукой такой номер не пройдёт — не сможет она жить в солёной воде, раз нет у неё соответствующей генетической программы.
Не меняет дела и тот факт, что у карликовых лососей увеличивается, по всей видимости, число некоторых генов: существует сколько угодно компьютерных программ, способных автоматически копировать некоторые файлы — была бы включена соответствующая опция.
К тому же надо иметь в виду, что генетические программы гораздо сложнее компьютерных. Как в большинстве компьютерных игр можно выбирать разные уровни сложности, так и генетические программы могут иметь разные режимы функционирования. Даже самые простые организмы — фаги и вирусы — не обязательно разрушают клетку, предварительно создав внутри неё свои многочисленные копии. Иногда они встраиваются в клеточный геном и передают свои копии потомкам материнской клетки. Что уж тут говорить о бактериях, а тем более о многоклеточных!
При изменении условий среды происходит перенастройка генетических программ. Если в запасе у тебя ничего подходящего нет или ты не успеваешь отреагировать вовремя — выбываешь из дальнейшей эволюционной игры[1]. Идёт отбор организмов с генетической программой, подходящей для новых условий.
Поверхностный наблюдатель может принять это явление за наследование благоприобретённых признаков, особенно если учесть, что среди особей, живущих и размножающихся некоторое время в изменившихся условиях, иногда появляются такие, у которых полезное свойство проявляется уже без всякого стимула со стороны среды.
Однако если проанализировать подобные случаи более внимательно, то выясняется, что эти особи — вновь возникшие мутанты, и отличаются они от прочих не только по интересующему нас признаку, но и по целому ряду других, никакого отношения „к упражнению органа“ не имеющих.
Более того, часто бывает, что генетическая адаптация идёт совсем не тем путём, что физиологическая. Например, у животных, попавших в горы, где недостаточно кислорода, увеличивается интенсивность дыхания и кислородная ёмкость крови, однако эти признаки и не думают наследоваться. Зато кое у кого из потомков высокогорного стада понижается интенсивность кровообращения да к тому же падает общая интенсивность обмена[2].
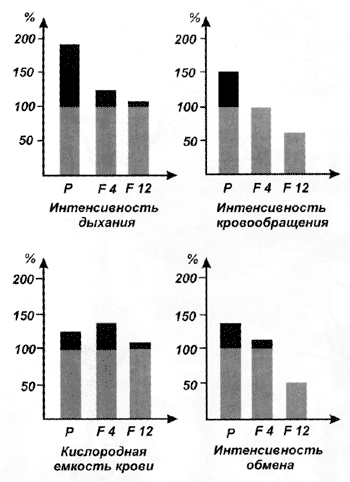
Изменение адаптивной реакции у животных, попавших в горы. Физиологические особенности прослежены в ряду поколений: Р — родительское поколение; F 4 — потомки 4-го поколения; F 12 — потомки 12-го поколения (по Ю.О. Раушенбаху, „Закономерности экогенеза домашних животных“, 1981)
Чтобы подобные изменения стали возможны, иногда не требуется даже новых мутаций достаточно перетасовать геном, скомбинировать его по-другому. Такая перетасовка идёт постоянно, и отбору всегда есть из чего выбирать[3]. Поэтому процесс адаптации к новым условиям обитания сопровождается изменениями в генофонде популяции под влиянием естественного отбора в нём начинают преобладать те варианты генов, которые раньше были редки.
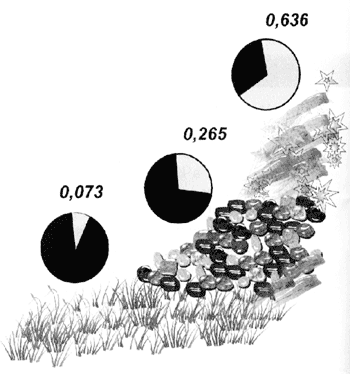
При длительной адаптации стада животных к условиям жизни в горах в ряду поколений происходит постепенная замена одного вида гемоглобина на другой (меняются частоты встречаемости разных вариантов гена гемогломина). Серым цветом обозначена доля особей с вариантом НЬ А, чёрным — с вариантом НЬ В в условиях равнин, среднегорья и высокогорья (по Ю.О. Раушенбаху, „Закономерности экогенеза домашних животных“, 1981)
Вот так и получается, что никаких новых признаков, кроме следов травм, организм в ходе жизни не приобретает. Особенности конкретных особей уже заданы их геномом, а среда позволяет проявиться одним потенциально возможным признакам и не даёт реализоваться другим. Если же в какой-то момент у организма не окажется в геноме подходящей программы, „природа заставляет особей приобрести“ синяки и шишки. Но они, как известно, не наследуются.
Как оценить „благо“?
А теперь предположим, что некий удивительный механизм всё-таки существует и он даёт возможность вносить в геномы отдельных клеток усовершенствования, позволяющие организму лучше приспосабливаться к изменениям среды обитания. Пусть, например, клетки печени научились обезвреживать какой-нибудь токсин и занесли это достижение в собственный геном. Как же отличить это генетическое изменение от случайных мутаций, которые с течением времени накапливаются во всех клетках организма?
Представьте себе, что автомобили на заводе собирают не по чертежам, а производят путём тщательного копирования изрядно послуживших машин, пусть даже в чём-то улучшенных хозяевами. Неужели покупателей обрадуют проржавевший кузов, заедающие тормоза и прочие беды престарелого автомобиля? Боюсь, что даже некоторые усовершенствования, внесённые умельцами, их в этом случае не утешат.
Между тем, если организмы будут передавать потомкам все изменения, происходящие в соматических клетках, получится то же самое. Дети неизбежно получат „в наследство“ всевозможные виды рака и дефекты обмена веществ — весь список поломок в клетках родителей, потому что различить „хорошие“ и „плохие“ изменения в генах на уровне клетки не всегда возможно. Разве знает раковая клетка, что она несёт гибель всему организму? Нет, конечно, хотя в данном случае патология налицо. А сколько существует зловещих мутаций, которые не могут проявиться в тех соматических клетках, где они возникли, но способны здорово навредить, если их передать потомкам?
В случае клеток, отвечающих за иммунитет, природа вроде бы обошла трудность с отбором перспективных кандидатов: клетка, обладающая нужным признаком, поощряется усиленным размножением. Это, фактически, естественный отбор в пределах организма, что отмечают и авторы книги „Что, если Ламарк прав?“. Может быть, и в самом деле клетки с опасными мутациями просто потонут в полноводной реке „передовиков производства“?
Беда, однако, в том, что у высших животных интенсивно делятся только немногие клетки. Более того, могут возникнуть и вовсе неожиданные трудности: куда, к примеру, записать информацию безъядерному эритроциту, добившемуся выдающихся успехов в переносе кислорода? Ведь в процессе дифференцировки эта клетка теряет свой геном и уже не может ни мутировать, ни делиться! Пока есть геном — нет эритроцита, есть эритроцит — нет генома. Прямо сказка про дудочку и кувшинчик получается: если есть куда собирать ягодки — их не видно, а когда ягодки видны — их некуда собирать.
Принципы кибернетики, как правильно заметил в своей статье Л. Верховский (см. „Химию и жизнь“, 2003, № 2), действительно требуют наличия обратной связи — от фенотипа к генотипу. Однако изобретать её заново нет никакой необходимости, такой путь передачи информации давно известен. Только вот, как показал ещё в середине XX века академик И.И. Шмальгаузен в своей книге „Кибернетические вопросы биологии“, осуществляется обратная связь не на уровне клетки. Благо или вред от любого признака проявляются, как правило, на уровне целого организма[4]. А выставить оценку может только среда обитания — через естественный отбор.
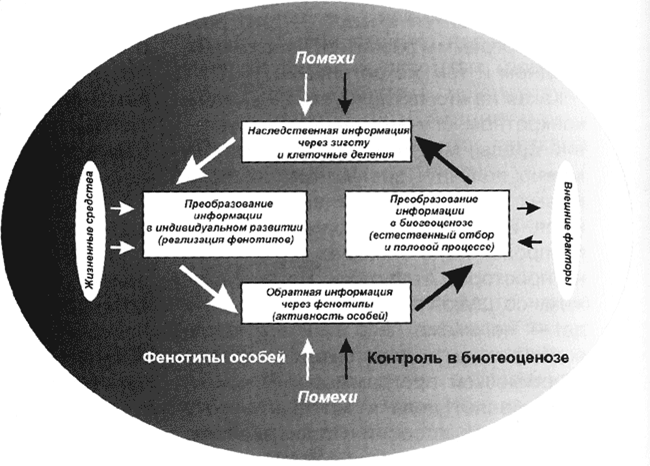
Схема регулирующего механизма эволюции с учётом обратной связи между фенотипом и генотипом (по И.И. Шмальгаузену, „Кибернетические вопросы биологии“, 1968)
Как передать приобретённое потомкам?
Только теперь мы добрались наконец, до знаменитого „барьера Вейсмана“, и, чтобы попытаться его преодолеть, нам придётся предположить, что соматические клетки организма неким таинственным образом всё-таки накопили груз ценных мутаций. Как же переправить улучшенные гены в ДНК половых клеток?
Добраться до главной молекулы организма ничуть не легче, чем заполучить Кащееву смерть. ДНК в половых клетках аккуратно свёрнута, облеплена белками, упакована в ядро да ещё прикрыта сверху цитоплазмой и клеточной оболочкой о трёх слоях.
Даже генетическим инженерам, вооружённым всей мощью современных методов и приборов, далеко не всегда удаётся включить нужные гены в ДНК половых клеток — природа совсем не заинтересована в том, чтобы в геном нового организма попадало что либо постороннее.
Но может быть, путь к Кащеевой смерти знают три богатыря — Э. Стил, Р. Линдли и Р. Бланден — авторы нашумевшей книги „Что, если Ламарк прав?“, на которую постоянно ссылаются сторонники наследования благоприобретённых признаков? Да нет, они тоже этого не знают, а просто выражают уверенность, что барьер Вейсмана будет взят хотя „при современных возможностях результатов может не быть много лет“ (с 166).
Нескоро дело делается, зато скоро сказка сказывается. Нам предлагают много косвенных улик и предположений. Одно из самых перспективных, на взгляд авторов, состоит в том, что гены из соматических клеток в половые переносят вирусы. Но почему вдруг вирусы взялись помогать эволюции хозяев? Как они отличают новые варианты генов от старых? Как выделяют полезные мутации из огромного числа вредных? Ответов на эти „занудные“ вопросы в книге, увы, нет.
Между тем даже если вирусам и удастся перетащить кусочек ДНК из соматической в половую клетку — это будет абсолютно случайное событие. Просто произойдёт одна из многих мутаций, а мутации как раз и обеспечивают материалом эволюционный процесс.
Таким образом, барьер Вейсмана до сих пор никем не взят, как не взяты и два других барьера, упомянутых выше[5]. Наследование благоприобретённых признаков никем и никогда не было показано, как не развенчана до сих пор и „центральная догма молекулярной биологии“.
Нужна ли потомкам „блажь“ предков?
Ну а напоследок давайте задумаемся над простым вопросом: зачем понадобилось природе воздвигать „на пути прогресса“ те мощные барьеры, о которых мы столько рассуждали? Неужели для того, чтобы героически их преодолевать?
Нет, конечно. Барьеры — это тоже эволюционное достижение, и возникли они именно потому, что были необходимы. Приобрести новое полезное свойство ох как нелегко, — такими достижениями не разбрасываются, их стараются закрепить, а закрепив — оградить от утраты.
Скажем, пластичность стрелолиста, возникшая в процессе эволюции, очень полезное свойство: в воде, в воздухе и на поверхности воды растение имеет листья разной формы. Нужно ли конкретному экземпляру, выросшему на сухом месте, закреплять в генах и передавать потомкам характерную острую форму листа, если при этом они утеряют способность образовывать плавающие и подводные листья? Вряд ли это целесообразно. Стоит ли выбрасывать полезную вещь, полученную от предков, только потому, что лично тебе она не понадобилась? А вдруг она пригодится детям?
Центральная догма молекулярной биологии тоже имеет глубокий эволюционно-биологический смысл: ДНК (или, по современным взглядам, РНК), как очень правильно отметил С.В. Багоцкий (см „Химию и жизнь“, 2003, № 4), едва появившись, сразу же берёт под контроль эволюцию всех структур клетки — любая самодеятельность тут же пресекается, и наследование приобретённых признаков становится невозможным. И это вполне целесообразно, если молекулы проживают в клетке сообща. Если каждая из них будет иметь собственных наследников — внутри клетки неизбежна конкуренция, которая нарушит оптимальное соотношение молекул.
Своеобразный реликт времён белкового сепаратизма — прионы: молекулы белка, изменённые таким образом, что они оказываются способными перековывать на свой манер другие белки. Результат хорошо известен — это тяжёлые болезни вроде коровьего бешенства, которые ведут организм к неминуемой гибели.
Не случайно появляется в ходе эволюции и барьер Вейсмана. У растений он низок — из одного листочка часто может сформироваться целый организм. Это свойство используют цветоводы: из листьев с соматическими мутациями выводят новые сорта. Но у животных по мере усложнения их строения способность к размножению делением исчезает очень быстро. Более того, клетки, которые дадут начало половым, выделяются из общей массы на очень ранних стадиях развития организма. При этом они часто оказываются довольно далеко от будущих половых органов и вынуждены бывают долго пробираться к ним в процессе формирования организма.
Какой в этом смысл? Почему бы половым клеткам не возникнуть из первой попавшейся клетки? Представим, что каждая клетка животного может дать начало другим организмам, и это шанс увековечить себя со всеми своими индивидуальными особенностями. Вполне понятно, что в этом случае каждая клетка постарается бросить свой „пост“ и пробраться поближе к половым органам. Колоритная получится картина, не правда ли? Впрочем, хаос будет не намного меньше и в том случае, если клетки останутся на местах, а в половые органы отправят свои любимые гены. Вряд ли организм, допустивший подобную анархию, просуществует долго.
Так что выбора у эволюции нет: соматические и половые клетки надо разделять очень жёстко, окружая последние непроницаемыми барьерами. Любое вмешательство извне может привести к гибели последующие поколения, а эволюция должна продолжаться — живое вовсе не нацелено на самоуничтожение.
Примечания:
[1] Модель парфорсной эволюции В.В.Суслова показывает, как отбором на устойчивость к стрессу, созданному ухудшением условий среды или новой адаптивной задачей, продлить время до выбытия, увеличив вероятность решения адаптивной задачи, создав нужное приспособление «из готового». Вот как последнее выглядит на генетическом уровне.
[2]Одни и те же приспособления на уровне морфологии или физиологии в разных популяциях почти всегда связаны с разными генетическими изменениями. См., например, независимо приобретённые низкорослость и другие составляющие «пигмея» у народов бава и бака, или адаптация к высокогорью у тибетцев, жителей Анд, и эфиопских горцев, происходившая принципиально по-разному.
[3] Ещё один вариант, контринтуитивный для программистов и математиков, интересующихся биологией — одна и та же программа (ген) на разных процессорах (организмах, отличающихся планом строения) работает по-разному, производит разные продукты.См. статью Виктории Скобеевой «Кирпичи и корни дерева жизни«.
А всё потому, что материализм рулит: смысл текста, генетического в том числе, определяется обращающимся к нему компетентным читателем, и условиями этого обращения. А вот так, как кажется естественным программистам, руководителям и гуманитариям — творение по слову, написал программу, и по ней создалась вещь — так не бывает. Однако дискретный текст, устойчиво воспроизводящийся в поколениях, тоже должен существовать, без него никуда: книги и инструкции в библиотеке, к которой обращаешься, нельзя менять в связи с обращением.
[4] Сейчас известно, что поведение управляет генами, и понятно как — через изменение экспрессии в связи с изменением социальных задач, как изменения рыночной конъюнктуры меняют загрузку фабрик, производящих разную продукцию, вплоть до закрытия фабрик одного вида и закладки дополнительных другого. Но для этого дискретные объекты регуляции, ответственные за синтез разных важных продуктов (вроде гена стресса КРГ (кортикостерон-релизинг-гормона), антистрессорного гена (глюкокортикоидного рецептора, находящийся в гиппокампе), рецепторов окситоцина, опиоидов и т.д. биохимических посредников между стимулом на входе и ответом на выходе) уже должны существовать и устойчиво передаваться в поколениях. Только так регуляция извне, поведением ли социальных партнёров, изменением ли значимых факторов среды (они составляют умвельт данного вида) будет точной, а приспособление к ним — специфическим.
К воздействиям, не входящим в умвельт, особи не приспосабливаются, а просто «платят им дань», как жители современного общества — ДТП. Другое дело, размеры последней могут быть минимизированы как побочное следствие других приспособлений. Кибернетический механизм обратной связи между геномом и внешней средой подобно рассмотрен в этой работе автора.
[5] Чтобы расшифровать те интереснейшие частные случаи, где он был взят, необходима была именно история моргановской, а потом молекулярной генетики, с её дискретными генами и барьером Вейсмана. Статья в «Элементах» показывает, что с другого «плацдарма» — от идеи слитной наследственности и второго закона Ламарка — их не понять, ибо не выйдешь к идее репликаторов, копирующихся с ошибками, обращение к которым необходимо для осуществления признака в онтогенезе. Для этого на первом этапе необходимо использовать дискретные гены как единицы текста, и «точки приложения» процессов осуществления признака и регуляции оного, управляемых генными сетями. То и другое предполагает выбор, что усилить, что ослабить, в какой степени, а выбор требует отдельных субъектов выбора, они должны быть некисельными. Гены не «эгоистичны», это часть организма, управляемая как все прочие части, целым, а целостность оборачивается дифференциацией частей, почему гены отделены друг от друга и «фона» в виде прочего организма — среды осуществления генов, из которой приходят регуляторные воздействия. Диалектика состоит в том, что успехи эпигенетики или современные данные о «наследовании приобретённых признаков» имеют предпосылкой именно дискретные гены моргановской генетики. В природе — это то, над чем надстраиваются процессы описанные по ссылкам, чтобы получилось то, что получается, верхушка айсберга осуществления признака на «чужом» основании, в теории — печка, от которой танцует концепция, чтобы понять как это получается в соответствующих частных случаях.
Источник Всячина
Рекомендуем прочесть
 Антропология здоровья и материнская смертность в западной Африке
Антропология здоровья и материнская смертность в западной Африке Столичный дрейф: из каких городов чаще всего переезжают жить в Москву
Столичный дрейф: из каких городов чаще всего переезжают жить в Москву Тысячи англичан протестуют против приватизации здравоохранения
Тысячи англичан протестуют против приватизации здравоохранения Пазухи свода собора святого Марка и парадигма Панглосса: критика адаптационистской программы
Пазухи свода собора святого Марка и парадигма Панглосса: критика адаптационистской программы От испанского правительства требуют отозвать законопроект, ограничивающий право на аборт
От испанского правительства требуют отозвать законопроект, ограничивающий право на аборт  Вашингтон готовит интервенцию в Нигерию
Вашингтон готовит интервенцию в Нигерию С нами Феликс и Христос
С нами Феликс и Христос Самая страшная экологическая катастрофа планеты
Самая страшная экологическая катастрофа планеты
